Signaling by Retinoic Acid in Embryonic and Adult Hematopoiesis



Abstract
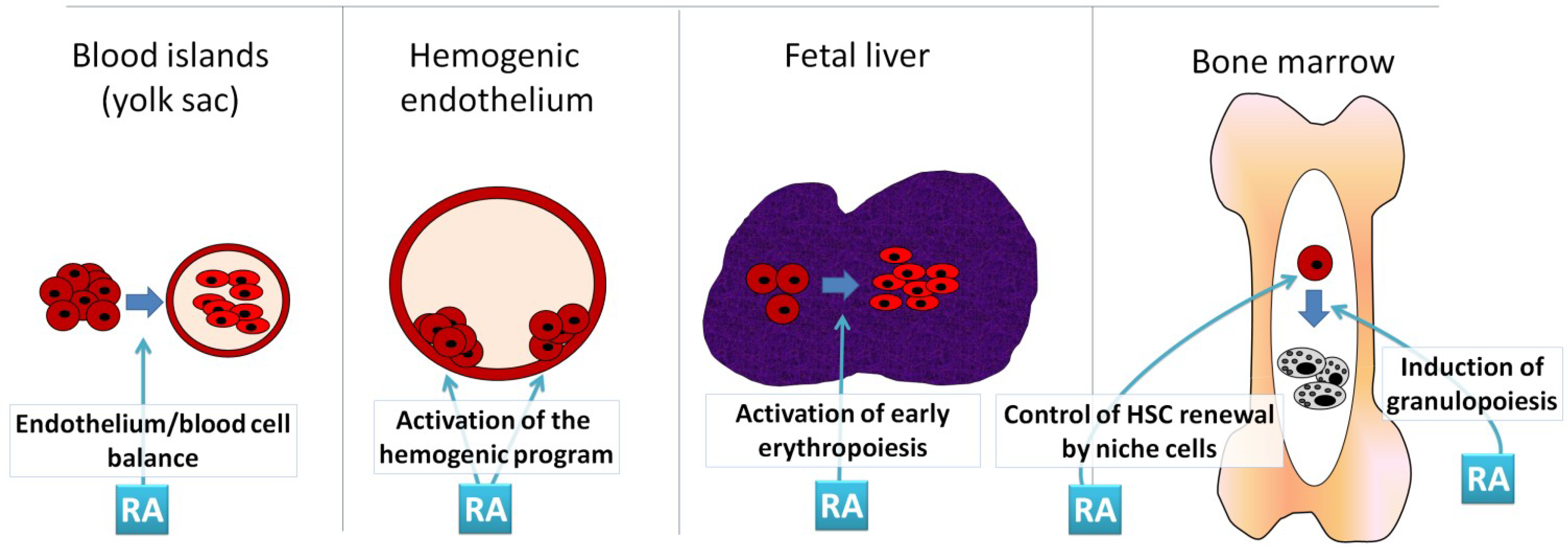
:1. Introduction
2. Transgenic Mouse Models for RA Signaling and Hematopoiesis
3. RA and Developmental Hematopoiesis

3.1. Early Hematoposiesis
3.2. Hematopoiesis in the AGM Region
3.3. Hematopoiesis in the Fetal Liver
4. RA and Adult Hematopoiesis
5. Vitamin D and Hematopoiesis
6. Potentiating Factors for RA Signaling in Hematopoiesis
7. Concluding Remarks and Future Directions

{kind=link}
| Knock out | Phenotype | References | ||
|---|---|---|---|---|
| RAR | α | RARα −/− | Early lethality (lifespan usually < 2 months). Accelerated granulocytic differentiation of bone marrow in culture. No defects in bone marrow or erythropoiesis in vivo. | [17,20,22,24,67,68] |
| RARα1 −/− | Normal phenotype. | [17,18] | ||
| β | RARβ −/− | Growth deficiency and several defects, although no apparent hematopoietic abnormalities. | [21,69,70] | |
| RARβ2 −/− | Normal phenotype. | [69] | ||
| RARβ1 −/− RARβ3 −/− | Normal phenotype. | [71] | ||
| γ | RARγ −/− | Early lethality (<40% survival by 3 months). Increased granulocyte/macrophage progenitors in bone marrow. Chronic myeloproliferative syndrome. Reduced erythropoiesis. Phenotype probably caused by RARϒ function in stroma. | [19,22,23,24,72] | |
| RARγ1 −/− | Growth deficiency and several defects, although no apparent hematopoietic abnormalities. | [73] | ||
| RARγ2 −/− | Normal phenotype. | [19,73] | ||
| Double mutants | RARα −/− RARγ −/− | Embryonic lethality, although no apparent hematopoietic abnormalities. | [19,20,74] | |
| RARα1 −/− RARγ −/− | Postnatal lethality. E18.5 bone marrow cells show a blockage of myelocitic differentiation. | [25] | ||
| Conditional deletion of RARα, RARγ and both receptors in EPO+ cells | Normal erythropoiesis. | [24] | ||
| RARα −/− RARγ1 −/− | No additional defects to those described for RXRα /RXRγ -/-. | [73,74] | ||
| RARα −/− RARγ2 −/− | No additional defects to those described for RXRα /RXRγ -/-. | [73] | ||
| RARβ2 −/−RARγ −/− | Ocular defects. No apparent hematopoietic abnormalities. | [74,75] | ||
| RARα −/− RARβ2 −/− | Several defects, although no apparent hematopoietic abnormalities. | [74] | ||
| RXR | α | RXRα −/− | Die at E13.5-E16.5 with severe cardiac defects. Fetal liver erythropoiesis is transiently compromised prior E12.5. | [14,15,39,65] |
| Conditional deletion of RXRα −/− in HSC | Normal phenotype. In vitro, RXRα −/− bone marrow cells form colonies more efficiently than those from control mice in the absence of ligand. This colony formation capacity is suppressed by the addition of 9cis-RA. | [16] | ||
| β | RXRβ −/− | 50% of mutants died during gestation or at birth, the other half are normal except for an abnormal spermatogenesis in males. | [12,13] | |
| γ | RXRγ −/− | Normal phenotype. | [12] | |
| Double mutants | RXRα −/− RXRϒ −/− | Growth deficiency and several defects, although no apparent hematopoietic abnormalities. | [12] | |
| RXRβ −/− RXRγ −/− | No additional defects to those described for RXRα /RXRβ −/−. | [12] | ||
| RXRα −/− RXRβ −/− RXRϒ −/− | Growth deficiency and male sterility, no apparent hematopoietic abnormalities. | [12] | ||
| RALDH | 2 | RALDH2 −/− | Die at E10.5 with multiple anomalies. Vascular defects due to an abnormal endothelial cell development. Hematopoietic defects not described. | [9,28,29,30,37] |
| Conditional deletion of RALDH2 in VE-cadherin expressing cells | Defective yolk sac hematopoiesis. No long-term HSC formation in AGM (no reconstitution when transplanted into irradiated mice). | [35] | ||
| 3 | RALDH3 −/− | Die at birth because respiratory failure. No apparent hematopoietic abnormalities. | [8] | |
| Vitamin A deficiency | Mice fed a vitamin A deficient diet | Expansion of myeloid cells in bone marrow, spleen and peripheral blood. No changes in CFU potential. | [45] | |
| CRBPI −/− | Low vitamin A store. When fed with vitamin A deficient diet, animals develop an expansion of neutrophils in spleen and peripheral blood with an increase of relatively immature granulocytes. | [20] | ||
Acknowledgments
Author Contributions
Conflicts of Interest
References and Notes
- Clements, W.K.; Traver, D. Signalling pathways that control vertebrate haematopoietic stem cell specification. Nat. Rev. Immunol. 2013, 13, 336–48. [Google Scholar] [CrossRef]
- Cao, H.; Oteiza, A.; Nilsson, S.K. Understanding the role of the microenvironment during definitive hemopoietic development. Exp. Hematol. 2013, 41, 761–768. [Google Scholar] [CrossRef]
- Morrison, S.J.; Scadden, D.T. The bone marrow niche for haematopoietic stem cells. Nature 2014, 16, 327–334. [Google Scholar] [CrossRef]
- Hirschi, K.K. Hemogenic endothelium during development and beyond. Blood 2012, 24, 4823–4827. [Google Scholar] [CrossRef]
- Costa, G.; Kouskoff, V.; Lacaud, G. Origin of blood cells and HSC production in the embryo. Trends Immunol. 2012, 33, 215–223. [Google Scholar] [CrossRef]
- Swiers, G.; Baumann, C.; O’Rourke, J.; Giannoulatou, E.; Taylor, S.; Joshi, A.; de Bruijn, M.F.T. Early dynamic fate changes in haemogenic endothelium characterized at the single-cell level. Nat. Commun. 2013, 4, 1–10. [Google Scholar]
- Fan, X.; Molotkov, A.; Manabe, S.; Donmoyer, C.M.; Deltour, L.; Foglio, M.H.; Cuenca, A.E.; Blaner, W.S.; Lipton, S.A.; Duester, G. Targeted disruption of Aldh1a1 (Raldh1) provides evidence for a complex mechanism of retinoic acid synthesis in the developing retina. Mol. Cell. Biol. 2003, 23, 4637–4648. [Google Scholar] [CrossRef]
- Dupé, V.; Matt, N.; Garnier, J.M.; Chambon, P.; Mark, M.; Ghyselinck, N.B. A newborn lethal defect due to inactivation of retinaldehyde dehydrogenase type 3 is prevented by maternal retinoic acid treatment. Proc. Natl. Acad. Sci. USA 2003, 25, 14036–14041. [Google Scholar]
- Niederreither, K.; Subbarayan, V.; Dollé, P.; Chambon, P. Embryonic retinoic acid synthesis is essential for early mouse post-implantation development. Nat. Genet. 1999, 21, 444–448. [Google Scholar] [CrossRef]
- Rochette-Egly, C.; Germain, P. Dynamic and combinatorial control of gene expression by nuclear retinoic acid receptors (RARs). Nucl. Recept. Sign. 2009, 8, 1–18. [Google Scholar]
- McKenna, N.J. EMBO Retinoids 2011: Mechanisms, biology and pathology of signaling by retinoic acid and retinoic acid receptors. Nucl. Recept. Sign. 2012, 10, 8–12. [Google Scholar]
- Krezel, W.; Dupé, V.; Mark, M.; Dierich, A.; Kastner, P.; Chambon, P. RXR gamma null mice are apparently normal and compound RXR alpha +/−/RXR beta −/−/RXR gamma −/− mutant mice are viable. Proc. Natl. Acad. Sci. USA 1996, 20, 9010–9014. [Google Scholar]
- Kastner, P.; Mark, M.; Leid, M.; Gansmuller, A.; Chin, W.; Grondona, J.M.; Décimo, D.; Krezel, W.; Dierich, A.; Chambon, P. Abnormal spermatogenesis in RXR beta mutant mice. Genes Dev. 1996, 10, 80–92. [Google Scholar] [CrossRef]
- Kastner, P.; Grondona, J.M.; Mark, M.; Gansmuller, A.; LeMeur, M.; Decimo, D.; Vonesch, J.L.; Dollé, P.; Chambon, P. Genetic analysis of RXR alpha developmental function: Convegence of RXR and RAR signaling pathways in heart and eye morphogenesis. Cell 1994, 78, 987–1003. [Google Scholar] [CrossRef]
- Sucov, H.M.; Dyson, E.; Gumeringer, C.L.; Price, J.; Chien, K.R.; Evans, R.M. RXR alpha mutant mice establish a genetic basis for vitamin A signaling in heart morphogenesis. Genes Dev. 1994, 8, 1007–1018. [Google Scholar] [CrossRef]
- Ricote, M.; Snyder, C.S.; Leung, H.; Chen, J.; Chien, K.R.; Glass, C.K. Normal hematopoiesis after conditional targeting of RXR α in murine hematopoietic stem/progenitor cells. J. Leukocyte Biol. 2006, 80, 850–861. [Google Scholar] [CrossRef]
- Lufkin, T.; Lohnes, D.; Mark, M.; Dierich, A.; Gorry, P.; Gaub, M.P.; Lemeur, M.; Chambon, P. High postnatal lethality and testis degeneration in retinoic acid receptor alpha mutant mice. Proc. Natl. Acad. Sci. USA 1993, 90, 7225–7229. [Google Scholar] [CrossRef]
- Li, E.; Sucov, H.M.; Lee, K.F.; Evans, R.M.; Jaenisch, R. Normal development and growth of mice carrying a targeted disruption of the alpha 1 retinoic acid receptor gene. Proc. Natl. Acad. Sci. USA 1993, 90, 1590–1594. [Google Scholar] [CrossRef]
- Lohnes, D.; Mark, M.; Mendelsohn, C.; Dollé, P.; Dierich, A.; Gorry, P.; Gansmuller, A.; Chambon, P. Function of the retinoic acid receptors (RARs) during development (I). Craniofacial and skeletal abnormalities in RAR double mutants. Development 1994, 120, 2723–2748. [Google Scholar]
- Kastner, P.; Lawrence, H.J.; Waltzinger, C.; Ghyselinck, N.B.; Chambon, P.; Chan, S. Positive and negative regulation of granulopoiesis by endogenous RARalpha. Blood 2001, 97, 1314–1320. [Google Scholar] [CrossRef]
- Luo, J.; Pasceri, P.; Conlon, R.A.; Rossant, J.; Giguère, V. Mice lacking all isoforms of retinoic acid receptor beta develop normally and are susceptible to the teratogenic effects of retinoic acid. Mech. Dev. 1995, 53, 61–71. [Google Scholar] [CrossRef]
- Purton, L.E.; Dworkin, S.; Olsen, G.H.; Walkley, C.R.; Fabb, S.A.; Collins, S.J.; Chambon, P. RARgamma is critical for maintaining a balance between hematopoietic stem cell self-renewal and differentiation. J. Exp. Med. 2006, 203, 1283–1293. [Google Scholar] [CrossRef]
- Walkley, C.R.; Olsen, G.H.; Dworkin, S.; Fabb, S.A.; Swann, J.; McArthur, G.A.; Westmoreland, S.V.; Chambon, P.; Scadden, D.T.; Purton, L.E. A microenvironment-induced myeloproliferative syndrome caused by retinoic acid receptor gamma deficiency. Cell 2007, 15, 1097–1110. [Google Scholar]
- Dewamitta, S.R.; Joseph, C.; Purton, L.E.; Walkley, C.R. Erythroid-extrinsic regulation of normal erythropoiesis by retinoic acid receptors. Brit. J. Haematol. 2014, 164, 280–285. [Google Scholar] [CrossRef]
- Labrecque, J.; Allan, D.; Chambon, P.; Iscove, N.N.; Lohnes, D.; Hoang, T. Impaired granulocytic differentiation in vitro in hematopoietic cells lacking retinoic acid receptors alpha1 and gamma. Blood 1998, 92, 607–615. [Google Scholar]
- Lohnes, D.; Mark, M.; Mendelsohn, C.; Dollé, P.; Dierich, A.; Gorry, P.; Gansmuller, A.; Chambon, P. Function of the retinoic acid receptors (RARs) during development (I). Craniofacial and skeletal abnormalities in RAR double mutants. Development 1994, 120, 2723–2728. [Google Scholar]
- Purton, L.E. Roles of retinoids and retinoic acid receptors in the regulation of hematopoietic stem cell self-renewal and differentiation. PPAR Res. 2007. [Google Scholar] [CrossRef]
- Bohnsack, B.L.; Lai, L.; Dolle, P.; Hirschi, K.K. Signaling hierarchy downstream of retinoic acid that independently regulates vascular remodeling and endothelial cell proliferation. Genes Dev. 2004, 18, 1345–1358. [Google Scholar] [CrossRef]
- Lai, L.; Bohnsack, B.L.; Niederreither, K.; Hirschi, K.K. Retinoic acid regulates endothelial cell proliferation during vasculogenesis. Development 2003, 130, 6465–6474. [Google Scholar] [CrossRef]
- Goldie, L.C.; Lucitti, J.L.; Dickinson, M.E.; Hirschi, K.K. Cell signaling directing the formation and function of hemogenic endothelium during murine embryogenesis. Blood 2008, 112, 3194–3204. [Google Scholar] [CrossRef]
- Ghatpande, S. Retinoid signaling regulates primitive (yolk sac) hematopoiesis. Blood 2002, 99, 2379–2386. [Google Scholar] [CrossRef]
- De Jong, J.L.O.; Davidson, A.J.; Wang, Y.; Palis, J.; Opara, P.; Pugach, E.V.; Daley, G.; Zon, L.I. Interaction of retinoic acid and scl controls primitive blood development. Blood 2010, 116, 201–209. [Google Scholar] [CrossRef]
- Van Handel, B.; Montel-Hagen, A.; Sasidharan, R.; Nakano, H.; Ferrari, R.; Boogerd, C.J.; Schredelseker, J.; Wang, Y.; Hunter, S.; Org, T.; Zhou, J.; Li, X.; Pellegrini, M.; Chen, J.; Orkin, S.H.; Kurdistani, S.; Evans, S.; Nakano, A.; Mikkola, H.K.A. Scl represses cardiomyogenesis in prospective hemogenic endothelium and endocardium. Blood 2010, 116, 201–209. [Google Scholar] [CrossRef]
- Nakano, H.; Nakano, H.; Liu, X.; Arshi, A.; Nakashima, Y.; van Handel, B.; Sasidharan, R.; Harmon, A.W.; Shin, J.; Schwartz, R.J.; Conway, S.; Harvey, R.P.; Pashmforoush, M.; Mikkola, H.K.A.; Nakano, A. Hemogenic endocardium contributes to transient definitive hematopoiesis. Nat. Commun. 2013, 5, 1564–1575. [Google Scholar]
- Dieterlen-Lièvre, F.; Pouget, C.; Bollérot, K.; Jaffredo, T. Are intra-aortic hemopoietic cells derived from endothelial cells during ontogeny? Trends Cardiovasc. Med. 2006, 16, 128–139. [Google Scholar] [CrossRef]
- Dzierzak, E.; Speck, N.A. Of lineage and legacy-the development of mammalian hemopoetic stem cells. Nat. Immunol. 2008, 9, 129–136. [Google Scholar] [CrossRef]
- Chanda, B.; Ditadi, A.; Iscove, N.N.; Keller, G. Retinoic acid signaling is essential for embryonic hematopoietic stem cell development. Cell 2013, 155, 215–227. [Google Scholar] [CrossRef]
- Ruiz-Herguido, C.; Guiu, J.; D’Altri, T.; Inglés-Esteve, J.; Dzierzak, E.; Espinosa, L.; Bigas, A. Hematopoietic stem cell development requires transient Wnt/β-catenin activity. J. Exp. Med. 2012, 209, 1457–1468. [Google Scholar] [CrossRef]
- Marcelo, K.L.; Sills, T.M.; Coskun, S.; Vasavada, H.; Sanglikar, S.; Goldie, L.C.; Hirschi, K.K. Hemogenic endothelial cell specification requires c-Kit, Notch Signaling, and p27-mediated cell-cycle control. Dev. Cell 2013, 27, 504–515. [Google Scholar] [CrossRef]
- Burns, C.; Galloway, J.; Smith, A.; Keefe, M.; Cashman, T.; Paik, E.; Mayhall, E.; Amsterdam, A.; Zon, L.I. A genetic screen in zebrafish defines a hierarchical network of pathways required for hematopoietic stem cell emergence. Blood 2009, 113, 5776–5782. [Google Scholar] [CrossRef]
- Makita, T.; Duncan, S.A.; Sucov, H.M. Retinoic acid, hypoxia, and GATA factors cooperatively control the onset of fetal liver erythropoietin expression and erythropoietic differentiation. Dev. Biol. 2005, 280, 59–72. [Google Scholar] [CrossRef]
- Varnum-Finney, B.; Xu, L.; Brashem-Stein, C.; Nourigat, C.; Flowers, D.; Bakkour, S.; Pear, W.S.; Bernstein, I.D. Pluripotent, cytokine-dependent, hematopoietic stem cells are immortalized by constitutive Notch1 signaling. Nat. Med. 2000, 6, 1278–1281. [Google Scholar] [CrossRef]
- Antonchuk, J.; Sauvageau, G.; Humphries, R.K. HOXB4 overexpression mediates very rapid stem cell regeneration and competitive hematopoietic repopulation. Exp. Hematol. 2001, 29, 1125–1134. [Google Scholar] [CrossRef]
- Dzhagalov, I.; Chambon, P.; He, Y.W. Regulation of CD8+ T lymphocyte effector function and macrophage inflammatory cytokine production by retinoic acid receptor gamma. J. Immunol. 2007, 178, 2113–2121. [Google Scholar]
- Chute, J.P.; Muramoto, G.G.; Whitesides, J.; Colvin, M.; Safi, R.; Chao, N.J.; McDonnell, D.P. Inhibition of aldehyde dehydrogenase and retinoid signaling induces the expansion of human hematopoietic stem cells. Proc. Natl. Acad. Sci. USA 2006, 103, 11707–11712. [Google Scholar]
- Ghiaur, G.; Yegnasubramanian, S.; Perkins, B.; Gucwa, J.L.; Gerber, J.M.; Jones, R.J. Regulation of human hematopoietic stem cell self-renewal by the microenvironment’s control of retinoic acid signaling. Proc. Natl. Acad. Sci. USA 2013, 110, 16121–16126. [Google Scholar] [CrossRef]
- Kuwata, T.; Wang, I.M.; Tamura, T.; Ponnamperuma, R.M.; Levine, R.; Holmes, K.L.; Morse, H.C.; De Luca, L.M.; Ozato, K. Vitamin A deficiency in mice causes a systemic expansion of myeloid cells. Blood 2000, 95, 3349–3356. [Google Scholar]
- Walkley, C.R.; Yuan, Y.D.; Chandraratna, R.A.S.; McArthur, G.A. Retinoic acid receptor antagonism in vivo expands the numbers of precursor cells during granulopoiesis. Leukemia 2002, 16, 1763–1772. [Google Scholar] [CrossRef]
- Zelent, A.; Guidez, F.; Melnick, A.; Waxman, S.; Licht, J. Translocations of the RARalpha gene in acute promyelocytic leukemia. Oncogene 2001, 20, 7186–7203. [Google Scholar] [CrossRef]
- Tallman, M.S.; Andersen, J.W.; Schiffer, C.A.; Appelbaum, F.R.; Feusner, J.H.; Ogden, A.; Shepherd, L.; Willman, C.; Bloomfield, C.D.; Rowe, J.M.; Wiernik, P.H. All-trans-retinoic acid in acute promyelocytic leukemia. N. Engl. J. Med. 1997, 337, 1021–1028. [Google Scholar] [CrossRef]
- Huang, M.E.; Ye, Y.C.; Chen, S.R.; Chai, J.R.; Lu, J.X.; Zhoa, L.; Gu, L.J.; Wang, Z.Y. Use of all-trans retinoic acid in the treatment of acute promyelocytic leukemia. Blood 1988, 72, 567–572. [Google Scholar]
- Castaigne, S.; Chomienne, C.; Daniel, M.T.; Ballerini, P.; Berger, R.; Fenaux, P.; Degos, L. All-trans retinoic acid as a differentiation therapy for acute promyelocytic leukemia. I. Clinical results. Blood 1990, 76, 1704–1709. [Google Scholar]
- Warrell, R.P.; Frankel, S.R.; Miller, W.H.; Scheinberg, D.A.; Itri, L.M.; Hittelman, W.N.; Vyas, R.; Andreeff, M.; Tafuri, A.; Jakubowski, A. Differentiation therapy of acute promyelocytic leukemia with tretinoin (all-trans-retinoic acid). N. Engl. J. Med. 1991, 324, 1385–1393. [Google Scholar] [CrossRef]
- Thomas, M.; Sukhai, M.A.; Kamel-Reid, S. An emerging role for retinoid X receptor α in malignant hematopoiesis. Leukemia Res. 2012, 36, 1075–1081. [Google Scholar] [CrossRef]
- Di Rosa, M.; Malaguarnera, L.; Nicolosi, A.; Sanfilippo, C.; Mazzarino, C.; Pavone, P.; Berretta, M.; Cosentino, S.; Cacopardo, B.; Pinzone, M.R.; Nunnari, G. Vitamin D3: An ever green molecule. Front. Biosci. 2013, 5, 247–260. [Google Scholar]
- Luong, Q.T.; Koeffler, H.P. Vitamin D compounds in leukemia. J. Steroid Biochem. Mol. Biol. 2005, 97, 195–202. [Google Scholar] [CrossRef]
- O’Kelly, J.; Hisatake, J.; Hisatake, Y.; Bishop, J.; Norman, A.; Koeffler, H.P. Normal myelopoiesis but abnormal T lymphocyte responses in vitamin D receptor knockout mice. J. Clin. Invest. 2002, 109, 1091–1099. [Google Scholar] [CrossRef]
- Jeanson, N.T.; Scadden, D.T. Vitamin D receptor deletion leads to increased hematopoietic stem and progenitor cells residing in the spleen. Blood 2010, 116, 4126–4129. [Google Scholar] [CrossRef]
- Kawamori, Y.; Katayama, Y.; Asada, N.; Minagawa, K.; Sato, M.; Okamura, A.; Shimoyama, M.; Nakagawa, K.; Okano, T.; Tanimoto, M.; Kato, S.; Matsui, T. Role for vitamin D receptor in the neuronal control of the hematopoietic stem cell niche. Hematology 2010, 116, 5528–5535. [Google Scholar]
- Bunce, C.M.; Brown, G.; Hewison, M. Vitamin D and hematopoiesis. Trends Endocrinol. Metab. 1997, 8, 245–251. [Google Scholar] [CrossRef]
- Grande, A.; Montanari, M.; Tagliafico, E.; Manfredini, R.; Zanocco Marani, T.; Siena, M.; Tenedini, E.; Gallinelli, A.; Ferrari, S. Physiological levels of 1alpha, 25 dihydroxyvitamin D3 induce the monocytic commitment of CD34+ hematopoietic progenitors. J. Leukocyte Biol. 2002, 71, 641–651. [Google Scholar]
- Hall, A.C.; Juckett, M.B. The role of vitamin d in hematologic disease and stem cell transplantation. Nutrients 2013, 5, 2206–2221. [Google Scholar] [CrossRef]
- Olsson, I.L.; Breitman, T.R.; Gallo, R.C. Priming of human myeloid leukemic cell lines HL-60 and U-937 with retinoic acid for differentiation effects of cyclic adenosine 3':5'-monophosphate-inducing agents and a T-lymphocyte-derived differentiation factor. Cancer Res. 1982, 42, 3928–3933. [Google Scholar]
- Nakamaki, T.; Hino, K.; Yokoyama, A.; Hisatake, J.; Tomoyasu, S.; Honma, Y.; Hozumi, M.; Tsuruoka, N. Effect of cytokines on the proliferation and differentiation of acute promyelocytic leukemia cells: Possible relationship to the development of “retinoic acid syndrome”. Anticancer Res. 1994, 14, 817–823. [Google Scholar]
- Si, J.; Collins, S.J. IL-3-induced enhancement of retinoic acid receptor activity is mediated through Stat5, which physically associates with retinoic acid receptors in an IL-3-dependent manner. Blood 2002, 100, 4401–4409. [Google Scholar] [CrossRef]
- Johnson, B.S.; Mueller, L.; Si, J.; Collins, S.J. The cytokines IL-3 and GM-CSF regulate the transcriptional activity of retinoic acid receptors in different in vitro models of myeloid differentiation. Blood 2002, 99, 746–753. [Google Scholar] [CrossRef]
- Ghyselinck, N.B.; Dupé, V.; Dierich, A.; Messaddeq, N.; Garnier, J.M.; Rochette-Egly, C.; Chambon, P.; Mark, M. Role of the retinoic acid receptor beta (RARbeta) during mouse development. Int. J. Dev. Biol. 1997, 41, 425–447. [Google Scholar]
- Massaro, G.D.; Massaro, D.; Chambon, P. Retinoic acid receptor-alpha regulates pulmonary alveolus formation in mice after, but not during, perinatal period. Am. J. Physiol. Lung Cell. Mol. Physiol. 2003, 284, L431–L433. [Google Scholar]
- Mendelsohn, C.; Mark, M.; Dollé, P.; Dierich, A.; Gaub, M.P.; Krust, A.; Lampron, C.; Chambon, P. Retinoic acid receptor b2 null mutant mice appear normal. Dev. Biol. 1994, 166, 246–258. [Google Scholar] [CrossRef]
- Massaro, G.D.; Massaro, D.; Chan, W.Y.; Clerch, L.B.; Ghyselinck, N.; Chambon, P.; Chandraratna, R.A. Retinoic acid receptor-beta: An endogenous inhibitor of the perinatal formation of pulmonary alveoli. Physiol. Genomics 2000, 4, 51–571. [Google Scholar]
- Ghyselinck, N.B.; Wendling, O.; Messaddeq, N.; Dierich, A.; Lampron, C.; Décimo, D.; Viville, S.; Chambon, P.; Mark, M. Contribution of retinoic acid receptor beta isoforms to the formation of the conotruncal septum of the embryonic heart. Dev. Biol. 1998, 198, 303–318. [Google Scholar] [CrossRef]
- McGowan, S.; Jackson, S.K.; Jenkins-Moore, M.; Dai, H.H.; Chambon, P.; Snyder, J.M. Mice bearing deletions of retinoic acid receptors demonstrate reduced lung elastin and alveolar numbers. Am. J. Respir. Cell. Mol. Biol. 2000, 23, 162–167. [Google Scholar] [CrossRef]
- Subbarayan, V.; Kastner, P.; Mark, M.; Dierich, A.; Gorry, P.; Chambon, P. Limited specificity and large overlap of the functions of the mouse RAR gamma 1 and RAR gamma 2 isoforms. Mech. Dev. 1997, 66, 131–142. [Google Scholar] [CrossRef]
- Mendelsohn, C.; Lohnes, D.; Décimo, D.; Lufkin, T.; Lemeur, M.; Chambon, P.; Mark, M. Function of the retinoic acid receptors (RARs) during development (II) Multiple abnormalities at various stages of organogenesis in RAR double mutants. Development 1994, 2771, 2749–2771. [Google Scholar]
- Grondona, J.M.; Kastner, P.; Gansmuller, A.; Décimo, D.; Chambon, P.; Mark, M. Retinal dysplasia and degeneration in RARbeta2/RARgamma2 compound mutant mice. Development 1996, 122, 2173–2188. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cano, E.; Ariza, L.; Muñoz-Chápuli, R.; Carmona, R. Signaling by Retinoic Acid in Embryonic and Adult Hematopoiesis. J. Dev. Biol. 2014, 2, 18-33. https://doi.org/10.3390/jdb2010018
Cano E, Ariza L, Muñoz-Chápuli R, Carmona R. Signaling by Retinoic Acid in Embryonic and Adult Hematopoiesis. Journal of Developmental Biology. 2014; 2(1):18-33. https://doi.org/10.3390/jdb2010018
Chicago/Turabian StyleCano, Elena, Laura Ariza, Ramón Muñoz-Chápuli, and Rita Carmona. 2014. "Signaling by Retinoic Acid in Embryonic and Adult Hematopoiesis" Journal of Developmental Biology 2, no. 1: 18-33. https://doi.org/10.3390/jdb2010018
APA StyleCano, E., Ariza, L., Muñoz-Chápuli, R., & Carmona, R. (2014). Signaling by Retinoic Acid in Embryonic and Adult Hematopoiesis. Journal of Developmental Biology, 2(1), 18-33. https://doi.org/10.3390/jdb2010018