
Sonic Hedgehog Signaling and Development of the Dentition

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
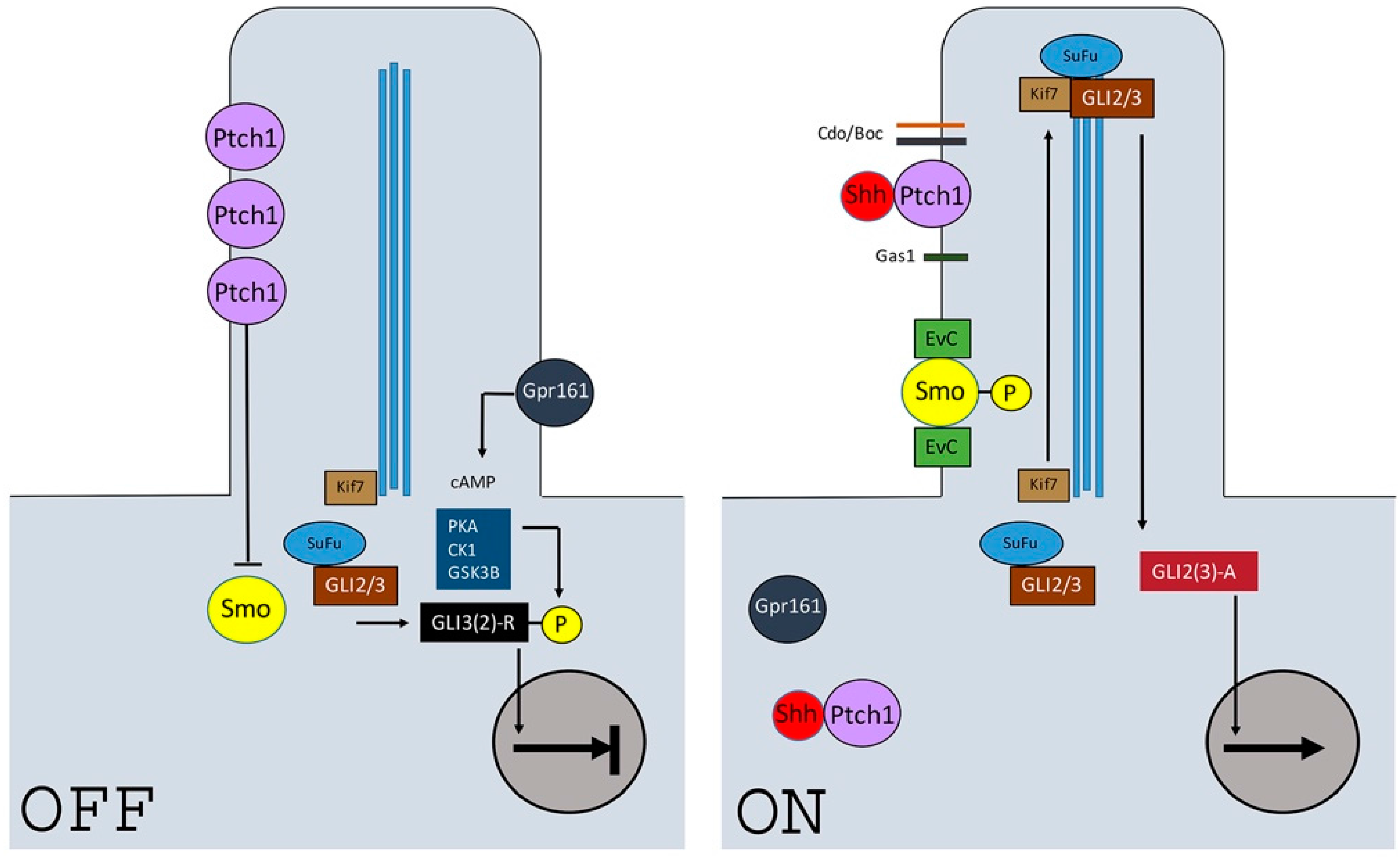
2. Shh Pathway Components and Signaling Activity
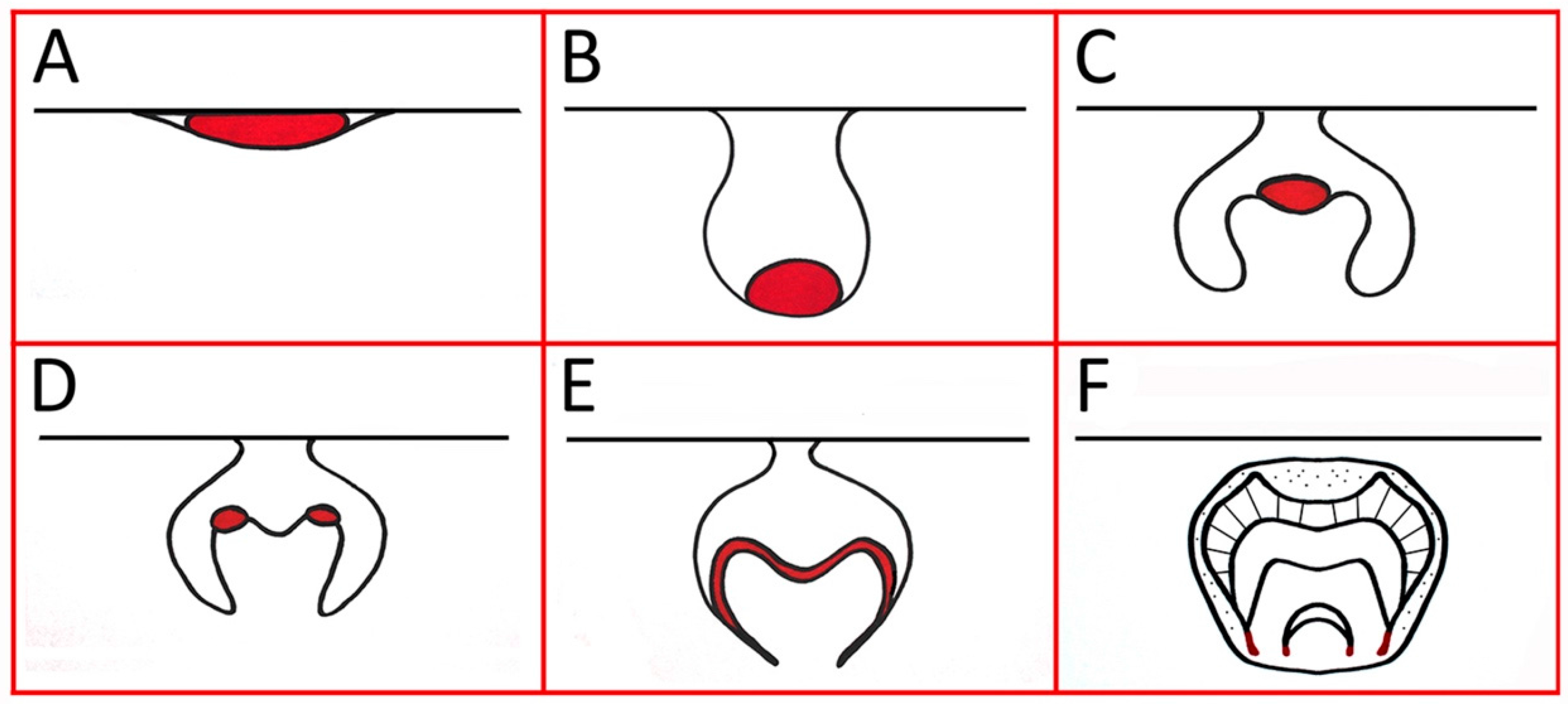
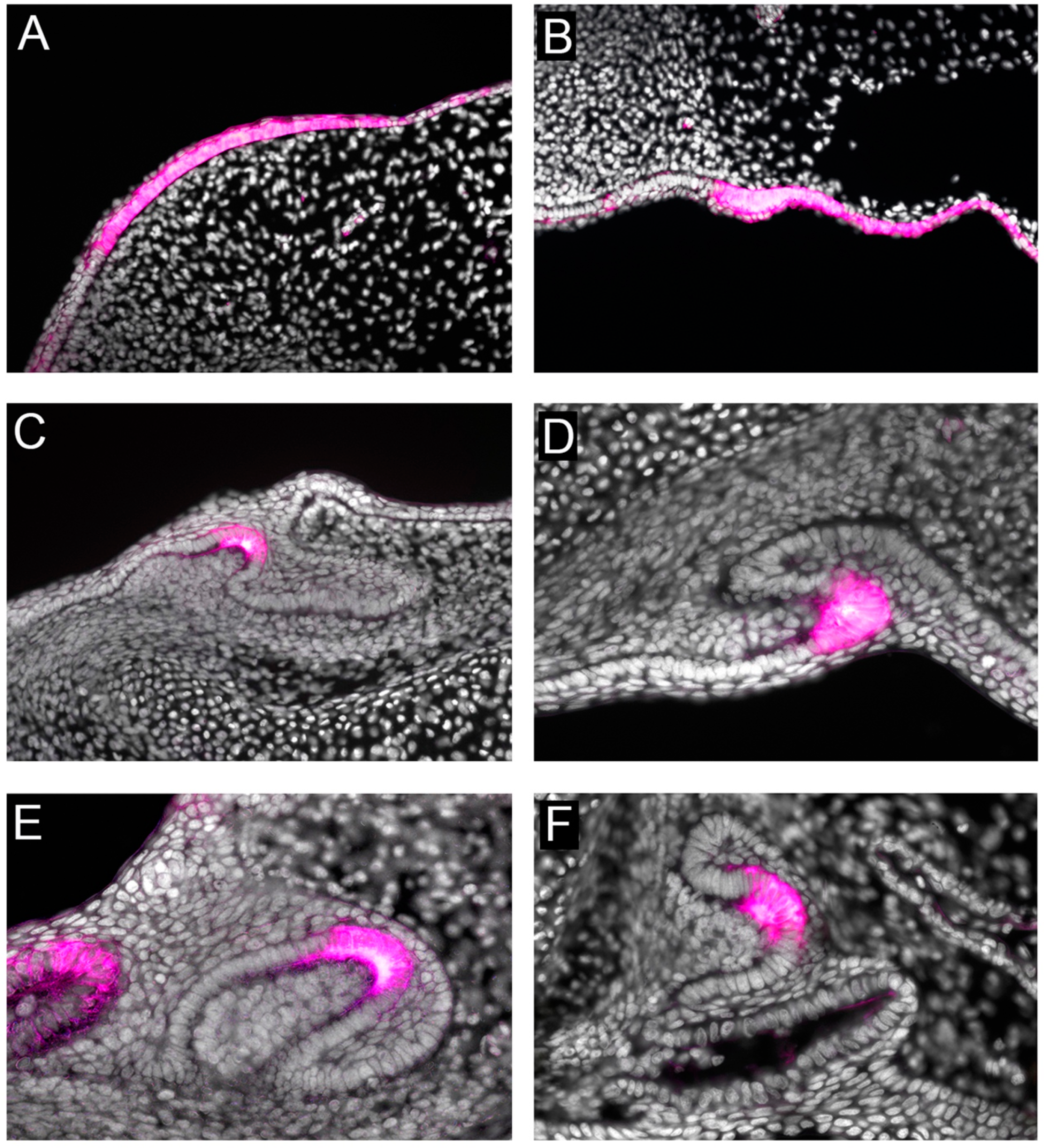
3. Shh Induces Invagination of the Early Dental Lamina
4. Shh Governs Early Molar Morphogenesis
5. A Lack of Shh Results in a Failure of Ameloblast Polarization
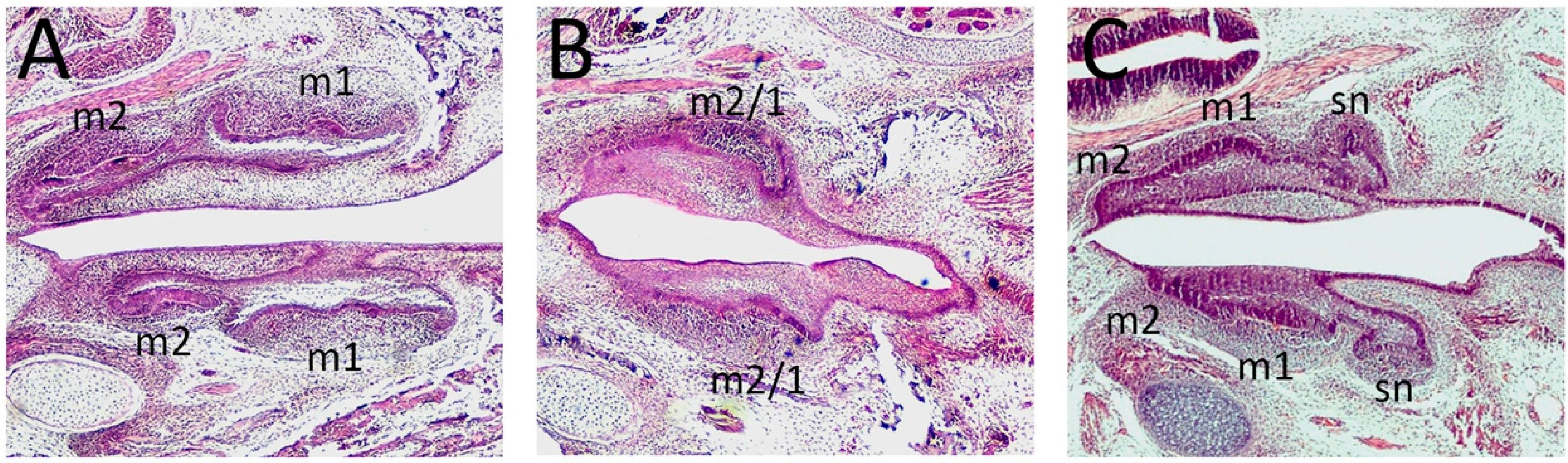
6. Disrupted Shh Signaling Causes Molar Fusion and Altered Cusp Morphology
7. Shh Influences Tooth Root Development
8. Shh and Its Role in Defining the Tooth Formula
9. Shh as a Regulator of Dental Stem Cells
10. Shh as a Universal Marker of Vertebrate Tooth Development
11. Conclusions
Acknowledgments
Conflicts of Interest
References
- Briscoe, J.; Therond, P.P. The mechanisms of hedgehog signalling and its roles in development and disease. Nat. Rev. Mol. Cell Biol. 2013, 14, 418–431. [Google Scholar] [CrossRef] [PubMed]
- Ingham, P.W.; McMahon, A.P. Hedgehog signaling in animal development: Paradigms and principles. Genes Dev. 2001, 15, 3059–3087. [Google Scholar] [CrossRef] [PubMed]
- Xavier, G.M.; Seppala, M.; Barrell, W.; Birjandi, A.A.; Geoghegan, F.; Cobourne, M.T. Hedgehog receptor function during craniofacial development. Dev. Biol. 2016, 415, 198–215. [Google Scholar] [CrossRef] [PubMed]
- Buchtova, M.; Handrigan, G.R.; Tucker, A.S.; Lozanoff, S.; Town, L.; Fu, K.; Diewert, V.M.; Wicking, C.; Richman, J.M. Initiation and patterning of the snake dentition are dependent on sonic hedgehog signaling. Dev. Biol. 2008, 319, 132–145. [Google Scholar] [CrossRef] [PubMed]
- Handrigan, G.R.; Richman, J.M. Autocrine and paracrine shh signaling are necessary for tooth morphogenesis, but not tooth replacement in snakes and lizards (squamata). Dev. Biol. 2010, 337, 171–186. [Google Scholar] [CrossRef] [PubMed]
- Rasch, L.J.; Martin, K.J.; Cooper, R.L.; Metscher, B.D.; Underwood, C.J.; Fraser, G.J. An ancient dental gene set governs development and continuous regeneration of teeth in sharks. Dev. Biol. 2016, 415, 347–370. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.M.; Fraser, G.J.; Chaplin, N.; Hobbs, C.; Graham, A. Reiterative pattern of sonic hedgehog expression in the catshark dentition reveals a phylogenetic template for jawed vertebrates. Proc. Biol. Sci. 2009, 276, 1225–1233. [Google Scholar] [CrossRef] [PubMed]
- Bitgood, M.J.; McMahon, A.P. Hedgehog and bmp genes are coexpressed at many diverse sites of cell-cell interaction in the mouse embryo. Dev. Biol. 1995, 172, 126–138. [Google Scholar] [CrossRef] [PubMed]
- Hardcastle, Z.; Mo, R.; Hui, C.C.; Sharpe, P.T. The shh signalling pathway in tooth development: Defects in gli2 and gli3 mutants. Development 1998, 125, 2803–2811. [Google Scholar] [PubMed]
- Iseki, S.; Araga, A.; Ohuchi, H.; Nohno, T.; Yoshioka, H.; Hayashi, F.; Noji, S. Sonic hedgehog is expressed in epithelial cells during development of whisker, hair, and tooth. Biochem. Biophys. Res. Commun. 1996, 218, 688–693. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Seppala, M.; Zoupa, M.; Cobourne, M.T. Hedgehog pathway gene expression during early development of the molar tooth root in the mouse. Gene Expr. Patterns 2007, 7, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Vaahtokari, A.; Åberg, T.; Jernvall, J.; Keranen, S.; Thesleff, I. The enamel knot as a signaling center in the developing mouse tooth. Mech. Dev. 1996, 54, 39–43. [Google Scholar] [CrossRef]
- Hu, X.; Zhang, S.; Chen, G.; Lin, C.; Huang, Z.; Chen, Y.; Zhang, Y. Expression of shh signaling molecules in the developing human primary dentition. BMC Dev. Biol. 2013, 13, 11. [Google Scholar] [CrossRef] [PubMed]
- Cobourne, M.T.; Hardcastle, Z.; Sharpe, P.T. Sonic hedgehog regulates epithelial proliferation and cell survival in the developing tooth germ. J. Dent. Res. 2001, 80, 1974–1979. [Google Scholar] [CrossRef] [PubMed]
- Dassule, H.R.; Lewis, P.; Bei, M.; Maas, R.; McMahon, A.P. Sonic hedgehog regulates growth and morphogenesis of the tooth. Development 2000, 127, 4775–4785. [Google Scholar] [PubMed]
- Gritli-Linde, A.; Bei, M.; Maas, R.; Zhang, X.M.; Linde, A.; McMahon, A.P. Shh signaling within the dental epithelium is necessary for cell proliferation, growth and polarization. Development 2002, 129, 5323–5337. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, L.; Cobourne, M.; Naylor, S.; Smalley, M.; Dale, T.; Sharpe, P.T. Wnt/shh interactions regulate ectodermal boundary formation during mammalian tooth development. Proc. Natl. Acad. Sci. USA 2000, 97, 4520–4524. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.W.; Kwak, S.; Woolley, T.E.; Lee, M.J.; Kim, E.J.; Baker, R.E.; Kim, H.J.; Shin, J.S.; Tickle, C.; Maini, P.K.; et al. Interactions between shh, sostdc1 and wnt signaling and a new feedback loop for spatial patterning of the teeth. Development 2011, 138, 1807–1816. [Google Scholar] [CrossRef] [PubMed]
- Ohazama, A.; Haycraft, C.J.; Seppala, M.; Blackburn, J.; Ghafoor, S.; Cobourne, M.; Martinelli, D.C.; Fan, C.M.; Peterkova, R.; Lesot, H.; et al. Primary cilia regulate shh activity in the control of molar tooth number. Development 2009, 136, 897–903. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chatzeli, L.; Panousopoulou, E.; Tucker, A.S.; Green, J.B. Epithelial stratification and placode invagination are separable functions in early morphogenesis of the molar tooth. Development 2016, 143, 670–681. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Feng, J.; Seidel, K.; Shi, S.; Klein, O.; Sharpe, P.; Chai, Y. Secretion of shh by a neurovascular bundle niche supports mesenchymal stem cell homeostasis in the adult mouse incisor. Cell Stem Cell 2014, 14, 160–173. [Google Scholar] [CrossRef] [PubMed]
- Porter, J.A.; Ekker, S.C.; Park, W.J.; von Kessler, D.P.; Young, K.E.; Chen, C.H.; Ma, Y.; Woods, A.S.; Cotter, R.J.; Koonin, E.V.; et al. Hedgehog patterning activity: Role of a lipophilic modification mediated by the carboxy-terminal autoprocessing domain. Cell 1996, 86, 21–34. [Google Scholar] [CrossRef]
- Porter, J.A.; von Kessler, D.P.; Ekker, S.C.; Young, K.E.; Lee, J.J.; Moses, K.; Beachy, P.A. The product of hedgehog autoproteolytic cleavage active in local and long-range signalling. Nature 1995, 374, 363–366. [Google Scholar] [CrossRef] [PubMed]
- Pepinsky, R.B.; Zeng, C.; Wen, D.; Rayhorn, P.; Baker, D.P.; Williams, K.P.; Bixler, S.A.; Ambrose, C.M.; Garber, E.A.; Miatkowski, K.; et al. Identification of a palmitic acid-modified form of human sonic hedgehog. J. Biol. Chem. 1998, 273, 14037–14045. [Google Scholar] [CrossRef] [PubMed]
- Saha, K.; Schaffer, D.V. Signal dynamics in sonic hedgehog tissue patterning. Development 2006, 133, 889–900. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Goetz, J.A.; Suber, L.M.; Scott, W.J., Jr.; Schreiner, C.M.; Robbins, D.J. A freely diffusible form of sonic hedgehog mediates long-range signalling. Nature 2001, 411, 716–720. [Google Scholar] [CrossRef] [PubMed]
- Cobourne, M.T.; Sharpe, P.T. Expression and regulation of hedgehog-interacting protein during early tooth development. Connect. Tissue Res. 2002, 43, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.; Kicheva, A.; Ribeiro, A.; Blassberg, R.; Page, K.M.; Barnes, C.P.; Briscoe, J. Ptch1 and gli regulate shh signalling dynamics via multiple mechanisms. Nat. Commun. 2015, 6, 6709. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.M.; Porter, J.A.; Chiang, C.; Chang, D.T.; Beachy, P.A.; Tessier-Lavigne, M. Long-range sclerotome induction by sonic hedgehog: Direct role of the amino-terminal cleavage product and modulation by the cyclic amp signaling pathway. Cell 1995, 81, 457–465. [Google Scholar] [CrossRef]
- Harfe, B.D.; Scherz, P.J.; Nissim, S.; Tian, H.; McMahon, A.P.; Tabin, C.J. Evidence for an expansion-based temporal shh gradient in specifying vertebrate digit identities. Cell 2004, 118, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Bangs, F.; Anderson, K.V. Primary cilia and mammalian hedgehog signaling. Cold Spring Harb. Perspect. Biol. 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Goodrich, L.V.; Milenković, L.; Higgins, K.M.; Scott, M.P. Altered neural cell fates and medulloblastoma in mouse patched mutants. Science 1997, 277, 1109–1113. [Google Scholar] [CrossRef] [PubMed]
- Haycraft, C.J.; Banizs, B.; Aydin-Son, Y.; Zhang, Q.; Michaud, E.J.; Yoder, B.K. Gli2 and gli3 localize to cilia and require the intraflagellar transport protein polaris for processing and function. PLoS Genet. 2005, 1, e53. [Google Scholar] [CrossRef] [PubMed]
- Tukachinsky, H.; Kuzmickas, R.P.; Jao, C.Y.; Liu, J.; Salic, A. Dispatched and scube mediate the efficient secretion of the cholesterol-modified hedgehog ligand. Cell Rep. 2012, 2, 308–320. [Google Scholar] [CrossRef] [PubMed]
- Taipale, J.; Cooper, M.K.; Maiti, T.; Beachy, P.A. Patched acts catalytically to suppress the activity of smoothened. Nature 2002, 418, 892–897. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.M.; Ramalho-Santos, M.; McMahon, A.P. Smoothened mutants reveal redundant roles for shh and ihh signaling including regulation of l/r asymmetry by the mouse node. Cell 2001, 105, 781–792. [Google Scholar] [CrossRef]
- Corbit, K.C.; Aanstad, P.; Singla, V.; Norman, A.R.; Stainier, D.Y.; Reiter, J.F. Vertebrate smoothened functions at the primary cilium. Nature 2005, 437, 1018–1021. [Google Scholar] [CrossRef] [PubMed]
- Aberger, F.; Ruiz, I.A.A. Context-dependent signal integration by the gli code: The oncogenic load, pathways, modifiers and implications for cancer therapy. Semin. Cell Dev. Biol. 2014, 33, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Hui, C.C.; Angers, S. Gli proteins in development and disease. Annu. Rev. Cell. Dev. Biol. 2011, 27, 513–537. [Google Scholar] [CrossRef] [PubMed]
- Casali, A.; Struhl, G. Reading the hedgehog morphogen gradient by measuring the ratio of bound to unbound patched protein. Nature 2004, 431, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Struhl, G. Dual roles for patched in sequestering and transducing hedgehog. Cell 1996, 87, 553–563. [Google Scholar] [CrossRef]
- Martinelli, D.C.; Fan, C.M. Gas1 extends the range of hedgehog action by facilitating its signaling. Genes Dev. 2007, 21, 1231–1243. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.S.; Gao, M.; Feinleib, J.L.; Cotter, P.D.; Guadagno, S.N.; Krauss, R.S. Cdo: An oncogene-, serum-, and anchorage-regulated member of the ig/fibronectin type iii repeat family. J. Cell Biol. 1997, 138, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.S.; Mulieri, P.J.; Hu, Y.; Taliana, L.; Krauss, R.S. Boc, an ig superfamily member, associates with cdo to positively regulate myogenic differentiation. EMBO J. 2002, 21, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Biggs, L.C.; Mikkola, M.L. Early inductive events in ectodermal appendage morphogenesis. Semin. Cell Dev. Biol. 2014, 25–26, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Pispa, J.; Thesleff, I. Mechanisms of ectodermal organogenesis. Dev. Biol. 2003, 262, 195–205. [Google Scholar] [CrossRef]
- Cobourne, M.T.; Miletich, I.; Sharpe, P.T. Restriction of sonic hedgehog signalling during early tooth development. Development 2004, 131, 2875–2885. [Google Scholar] [CrossRef] [PubMed]
- Chiang, C.; Litingtung, Y.; Lee, E.; Young, K.E.; Corden, J.L.; Westphal, H.; Beachy, P.A. Cyclopia and defective axial patterning in mice lacking sonic hedgehog gene function. Nature 1996, 383, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, C.; Yamagishi, H.; Maeda, J.; Tsuchihashi, T.; Ivey, K.; Hu, T.; Srivastava, D. Sonic hedgehog is essential for first pharyngeal arch development. Pediatr. Res. 2006, 59, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Dassule, H.R.; McMahon, A.P. Analysis of epithelial-mesenchymal interactions in the initial morphogenesis of the mammalian tooth. Dev. Biol. 1998, 202, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Kettunen, P.; Karavanova, I.; Thesleff, I. Responsiveness of developing dental tissues to fibroblast growth factors: Expression of splicing alternatives of fgfr1, -2, -3, and of fgfr4; and stimulation of cell proliferation by fgf-2, -4, -8, and -9. Dev. Genet. 1998, 22, 374–385. [Google Scholar] [CrossRef]
- Kettunen, P.; Laurikkala, J.; Itaranta, P.; Vainio, S.; Itoh, N.; Thesleff, I. Associations of fgf-3 and fgf-10 with signaling networks regulating tooth morphogenesis. Dev. Dyn. 2000, 219, 322–332. [Google Scholar] [CrossRef]
- Sarkar, L. The Wnt Signalling Pathway in Tooth Development. Ph.D. Thesis, University of London, London, UK, 1999. [Google Scholar]
- Ahn, Y.; Sanderson, B.W.; Klein, O.D.; Krumlauf, R. Inhibition of wnt signaling by wise (sostdc1) and negative feedback from shh controls tooth number and patterning. Development 2010, 137, 3221–3231. [Google Scholar] [CrossRef] [PubMed]
- Klein, O.D.; Minowada, G.; Peterkova, R.; Kangas, A.; Yu, B.D.; Lesot, H.; Peterka, M.; Jernvall, J.; Martin, G.R. Sprouty genes control diastema tooth development via bidirectional antagonism of epithelial-mesenchymal fgf signaling. Dev. Cell 2006, 11, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Pummila, M.; Fliniaux, I.; Jaatinen, R.; James, M.J.; Laurikkala, J.; Schneider, P.; Thesleff, I.; Mikkola, M.L. Ectodysplasin has a dual role in ectodermal organogenesis: Inhibition of bmp activity and induction of shh expression. Development 2007, 134, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Murashima-Suginami, A.; Takahashi, K.; Sakata, T.; Tsukamoto, H.; Sugai, M.; Yanagita, M.; Shimizu, A.; Sakurai, T.; Slavkin, H.C.; Bessho, K. Enhanced bmp signaling results in supernumerary tooth formation in usag-1 deficient mouse. Biochem. Biophys. Res. Commun. 2008, 369, 1012–1016. [Google Scholar] [CrossRef] [PubMed]
- Nakatomi, M.; Hovorakova, M.; Gritli-Linde, A.; Blair, H.J.; MacArthur, K.; Peterka, M.; Lesot, H.; Peterkova, R.; Ruiz-Perez, V.L.; Goodship, J.A.; et al. Evc regulates a symmetrical response to shh signaling in molar development. J. Dent. Res. 2013, 92, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Perez, V.L.; Blair, H.J.; Rodriguez-Andres, M.E.; Blanco, M.J.; Wilson, A.; Liu, Y.N.; Miles, C.; Peters, H.; Goodship, J.A. Evc is a positive mediator of ihh-regulated bone growth that localises at the base of chondrocyte cilia. Development 2007, 134, 2903–2912. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Chen, W.; Chen, Y.; Jiang, J. Smoothened transduces hedgehog signal by forming a complex with evc/evc2. Cell Res. 2012, 22, 1593–1604. [Google Scholar] [CrossRef] [PubMed]
- Sharpe, P.T. Neural crest and tooth morphogenesis. Adv. Dent. Res. 2001, 15, 4–7. [Google Scholar] [CrossRef] [PubMed]
- Thesleff, I.; Hurmerinta, K. Tissue interactions in tooth development. Differentiation 1981, 18, 75–88. [Google Scholar] [CrossRef] [PubMed]
- Koyama, E.; Yamaai, T.; Iseki, S.; Ohuchi, H.; Nohno, T.; Yoshioka, H.; Hayashi, Y.; Leatherman, J.L.; Golden, E.B.; Noji, S.; et al. Polarizing activity, sonic hedgehog, and tooth development in embryonic and postnatal mouse. Dev. Dyn. 1996, 206, 59–72. [Google Scholar] [CrossRef]
- Gritli-Linde, A.; Lewis, P.; McMahon, A.P.; Linde, A. The whereabouts of a morphogen: Direct evidence for short- and graded long-range activity of hedgehog signaling peptides. Dev. Biol. 2001, 236, 364–386. [Google Scholar] [CrossRef] [PubMed]
- Kratochwil, K.; Galceran, J.; Tontsch, S.; Roth, W.; Grosschedl, R. Fgf4, a direct target of lef1 and wnt signaling, can rescue the arrest of tooth organogenesis in lef1(-/-) mice. Genes Dev. 2002, 16, 3173–3185. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Chu, E.Y.; Watt, B.; Zhang, Y.; Gallant, N.M.; Andl, T.; Yang, S.H.; Lu, M.M.; Piccolo, S.; Schmidt-Ullrich, R.; et al. Wnt/beta-catenin signaling directs multiple stages of tooth morphogenesis. Dev. Biol. 2008, 313, 210–224. [Google Scholar] [CrossRef] [PubMed]
- Nakatomi, M.; Morita, I.; Eto, K.; Ota, M.S. Sonic hedgehog signaling is important in tooth root development. J. Dent. Res. 2006, 85, 427–431. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Feng, J.; Li, J.; Zhao, H.; Ho, T.V.; Chai, Y. An nfic-hedgehog signaling cascade regulates tooth root development. Development 2015, 142, 3374–3382. [Google Scholar] [CrossRef] [PubMed]
- Steele-Perkins, G.; Butz, K.G.; Lyons, G.E.; Zeichner-David, M.; Kim, H.J.; Cho, M.I.; Gronostajski, R.M. Essential role for nfi-c/ctf transcription-replication factor in tooth root development. Mol. Cell. Biol. 2003, 23, 1075–1084. [Google Scholar] [CrossRef] [PubMed]
- Lesot, H.; Peterkova, R.; Viriot, L.; Vonesch, J.L.; Tureckova, J.; Peterka, M.; Ruch, J.V. Early stages of tooth morphogenesis in mouse analyzed by 3d reconstructions. Eur. J. Oral Sci. 1998, 106 (Suppl. S1), 64–70. [Google Scholar] [CrossRef] [PubMed]
- Peterkova, R.; Hovorakova, M.; Peterka, M.; Lesot, H. Three-dimensional analysis of the early development of the dentition. Aust. Dent. J. 2014, 59 (Suppl. S1), 55–80. [Google Scholar] [CrossRef] [PubMed]
- Peterkova, R.; Peterka, M.; Viriot, L.; Lesot, H. Dentition development and budding morphogenesis. J. Craniofac. Genet. Dev. Biol. 2000, 20, 158–172. [Google Scholar] [PubMed]
- Peterková, R.; Peterka, M.; Viriot, L.; Lesot, H. Development of the vestigial tooth primordia as part of mouse odontogenesis. Connect. Tissue Res. 2002, 43, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Viriot, L.; Lesot, H.; Vonesch, J.L.; Ruch, J.V.; Peterka, M.; Peterkova, R. The presence of rudimentary odontogenic structures in the mouse embryonic mandible requires reinterpretation of developmental control of first lower molar histomorphogenesis. Int. J. Dev. Biol. 2000, 44, 233–240. [Google Scholar] [PubMed]
- Tenzen, T.; Allen, B.L.; Cole, F.; Kang, J.S.; Krauss, R.S.; McMahon, A.P. The cell surface membrane proteins cdo and boc are components and targets of the hedgehog signaling pathway and feedback network in mice. Dev. Cell. 2006, 10, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Allen, B.L.; Tenzen, T.; McMahon, A.P. The hedgehog-binding proteins gas1 and cdo cooperate to positively regulate shh signaling during mouse development. Genes Dev. 2007, 21, 1244–1257. [Google Scholar] [CrossRef] [PubMed]
- Okada, A.; Charron, F.; Morin, S.; Shin, D.S.; Wong, K.; Fabre, P.J.; Tessier-Lavigne, M.; McConnell, S.K. Boc is a receptor for sonic hedgehog in the guidance of commissural axons. Nature 2006, 444, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Seppala, M.; Depew, M.J.; Martinelli, D.C.; Fan, C.M.; Sharpe, P.T.; Cobourne, M.T. Gas1 is a modifier for holoprosencephaly and genetically interacts with sonic hedgehog. J. Clin. Investig. 2007, 117, 1575–1584. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Kang, J.S.; Cole, F.; Yi, M.J.; Krauss, R.S. Cdo functions at multiple points in the sonic hedgehog pathway, and cdo-deficient mice accurately model human holoprosencephaly. Dev. Cell 2006, 10, 657–665. [Google Scholar] [CrossRef] [PubMed]
- Chuang, P.T.; McMahon, A.P. Vertebrate hedgehog signalling modulated by induction of a hedgehog- binding protein. Nature 1999, 397, 617–621. [Google Scholar] [CrossRef] [PubMed]
- Reich, A.; Sapir, A.; Shilo, B. Sprouty is a general inhibitor of receptor tyrosine kinase signaling. Development 1999, 126, 4139–4147. [Google Scholar] [PubMed]
- Harada, H.; Kettunen, P.; Jung, H.S.; Mustonen, T.; Wang, Y.A.; Thesleff, I. Localization of putative stem cells in dental epithelium and their association with notch and fgf signaling. J. Cell Biol. 1999, 147, 105–120. [Google Scholar] [CrossRef] [PubMed]
- Juuri, E.; Saito, K.; Ahtiainen, L.; Seidel, K.; Tummers, M.; Hochedlinger, K.; Klein, O.D.; Thesleff, I.; Michon, F. Sox2+ stem cells contribute to all epithelial lineages of the tooth via sfrp5+ progenitors. Dev. Cell 2012, 23, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Seidel, K.; Ahn, C.P.; Lyons, D.; Nee, A.; Ting, K.; Brownell, I.; Cao, T.; Carano, R.A.; Curran, T.; Schober, M.; et al. Hedgehog signaling regulates the generation of ameloblast progenitors in the continuously growing mouse incisor. Development 2010, 137, 3753–3761. [Google Scholar] [CrossRef] [PubMed]
- Rodda, S.J.; McMahon, A.P. Distinct roles for hedgehog and canonical wnt signaling in specification, differentiation and maintenance of osteoblast progenitors. Development 2006, 133, 3231–3244. [Google Scholar] [CrossRef] [PubMed]
- Passman, J.N.; Dong, X.R.; Wu, S.P.; Maguire, C.T.; Hogan, K.A.; Bautch, V.L.; Majesky, M.W. A sonic hedgehog signaling domain in the arterial adventitia supports resident sca1+ smooth muscle progenitor cells. Proc. Natl. Acad. Sci. USA 2008, 105, 9349–9354. [Google Scholar] [CrossRef] [PubMed]
- Fraser, G.J.; Graham, A.; Smith, M.M. Conserved deployment of genes during odontogenesis across osteichthyans. Proc. Biol. Sci. 2004, 271, 2311–2317. [Google Scholar] [CrossRef] [PubMed]
- Fraser, G.J.; Bloomquist, R.F.; Streelman, J.T. A periodic pattern generator for dental diversity. BMC Biol. 2008, 6, 32. [Google Scholar] [CrossRef] [PubMed]
- Martin, K.J.; Rasch, L.J.; Cooper, R.L.; Metscher, B.D.; Johanson, Z.; Fraser, G.J. Sox2+ progenitors in sharks link taste development with the evolution of regenerative teeth from denticles. Proc. Natl. Acad. Sci. USA 2016, 113, 14769–14774. [Google Scholar] [CrossRef] [PubMed]
- Fraser, G.J.; Bloomquist, R.F.; Streelman, J.T. Common developmental pathways link tooth shape to regeneration. Dev. Biol. 2013, 377, 399–414. [Google Scholar] [CrossRef] [PubMed]




© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seppala, M.; Fraser, G.J.; Birjandi, A.A.; Xavier, G.M.; Cobourne, M.T. Sonic Hedgehog Signaling and Development of the Dentition. J. Dev. Biol. 2017, 5, 6. https://doi.org/10.3390/jdb5020006
Seppala M, Fraser GJ, Birjandi AA, Xavier GM, Cobourne MT. Sonic Hedgehog Signaling and Development of the Dentition. Journal of Developmental Biology. 2017; 5(2):6. https://doi.org/10.3390/jdb5020006
Chicago/Turabian StyleSeppala, Maisa, Gareth J. Fraser, Anahid A. Birjandi, Guilherme M. Xavier, and Martyn T. Cobourne. 2017. "Sonic Hedgehog Signaling and Development of the Dentition" Journal of Developmental Biology 5, no. 2: 6. https://doi.org/10.3390/jdb5020006
APA StyleSeppala, M., Fraser, G. J., Birjandi, A. A., Xavier, G. M., & Cobourne, M. T. (2017). Sonic Hedgehog Signaling and Development of the Dentition. Journal of Developmental Biology, 5(2), 6. https://doi.org/10.3390/jdb5020006