The Roles of the Wnt-Antagonists Axin and Lrp4 during Embryogenesis of the Red Flour Beetle Tribolium castaneum

Abstract
:1. Introduction
2. Materials and Methods
3. Results
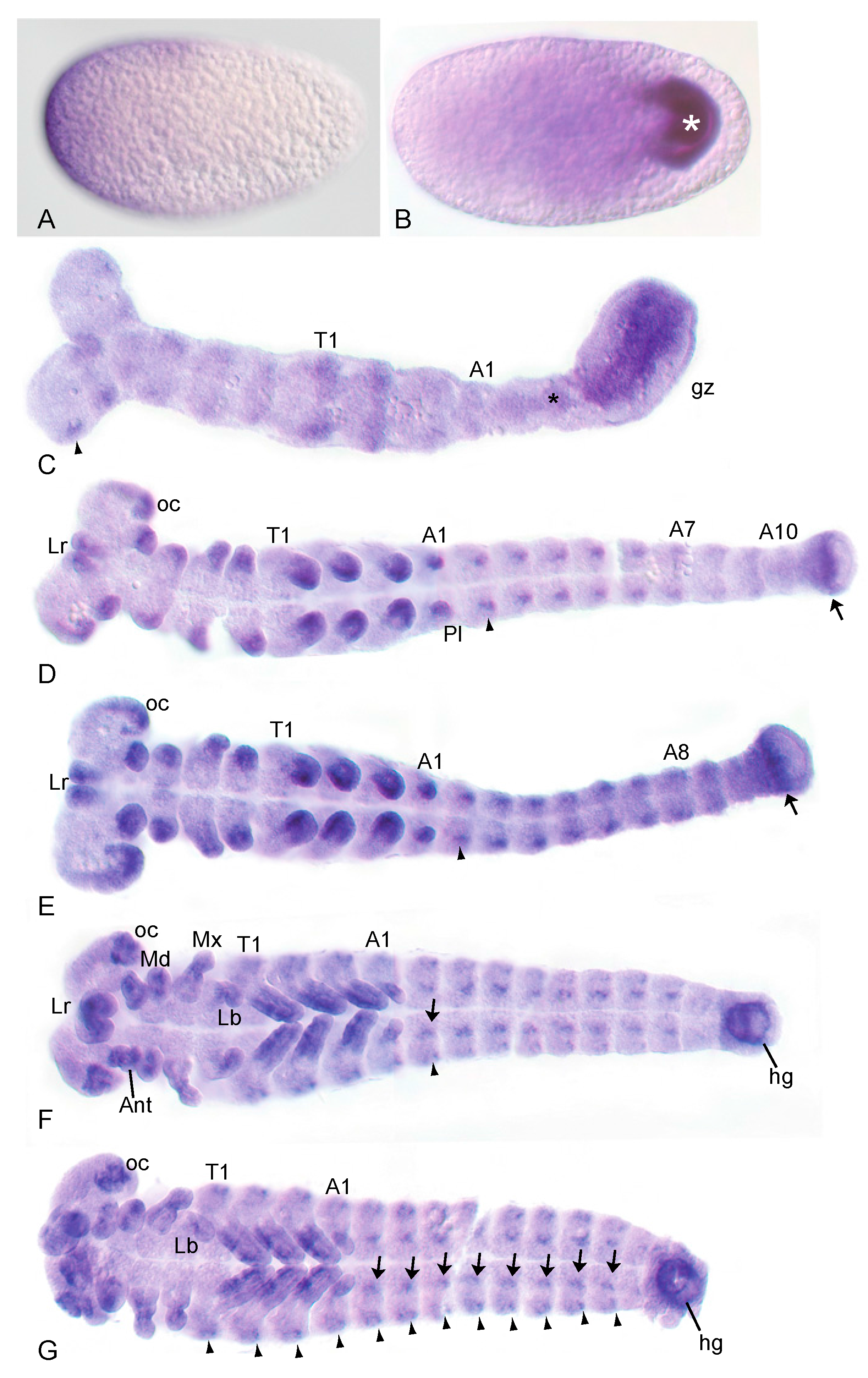
3.1. The Wnt-Inhibitor Tc-axin Is Asymmetrically Expressed in the Egg and Dynamically with Distinct Patterns during Further Embryogenesis
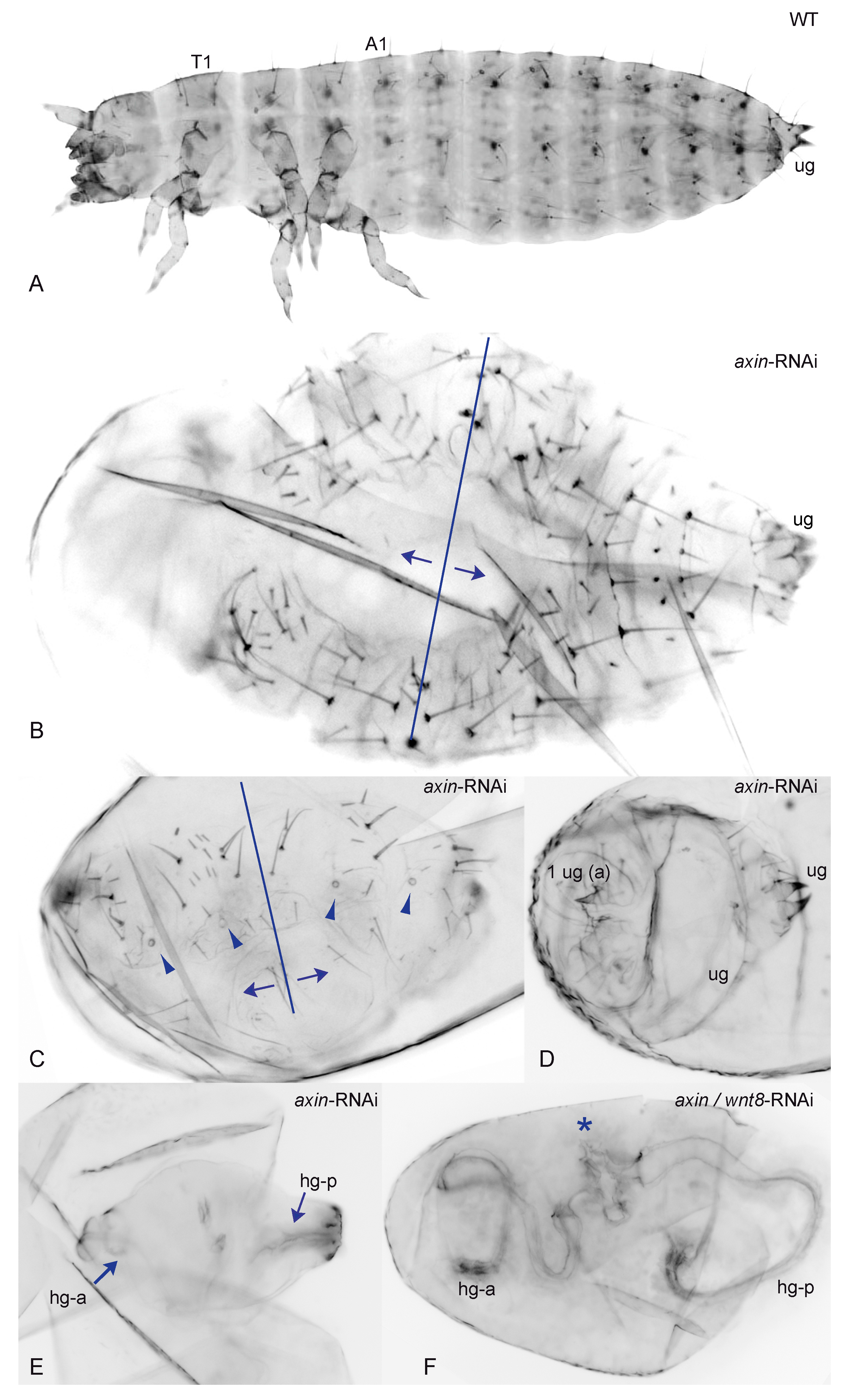
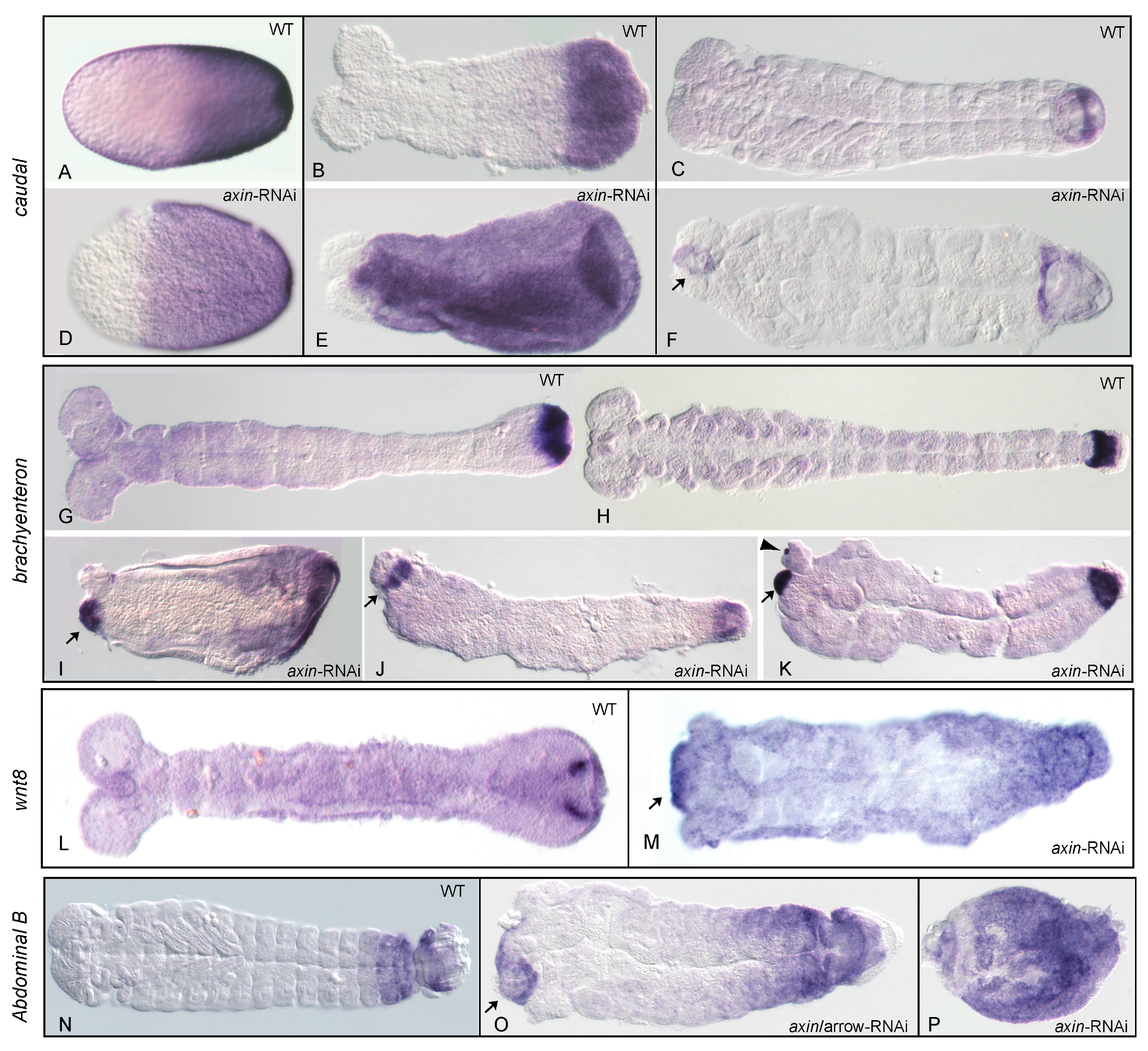
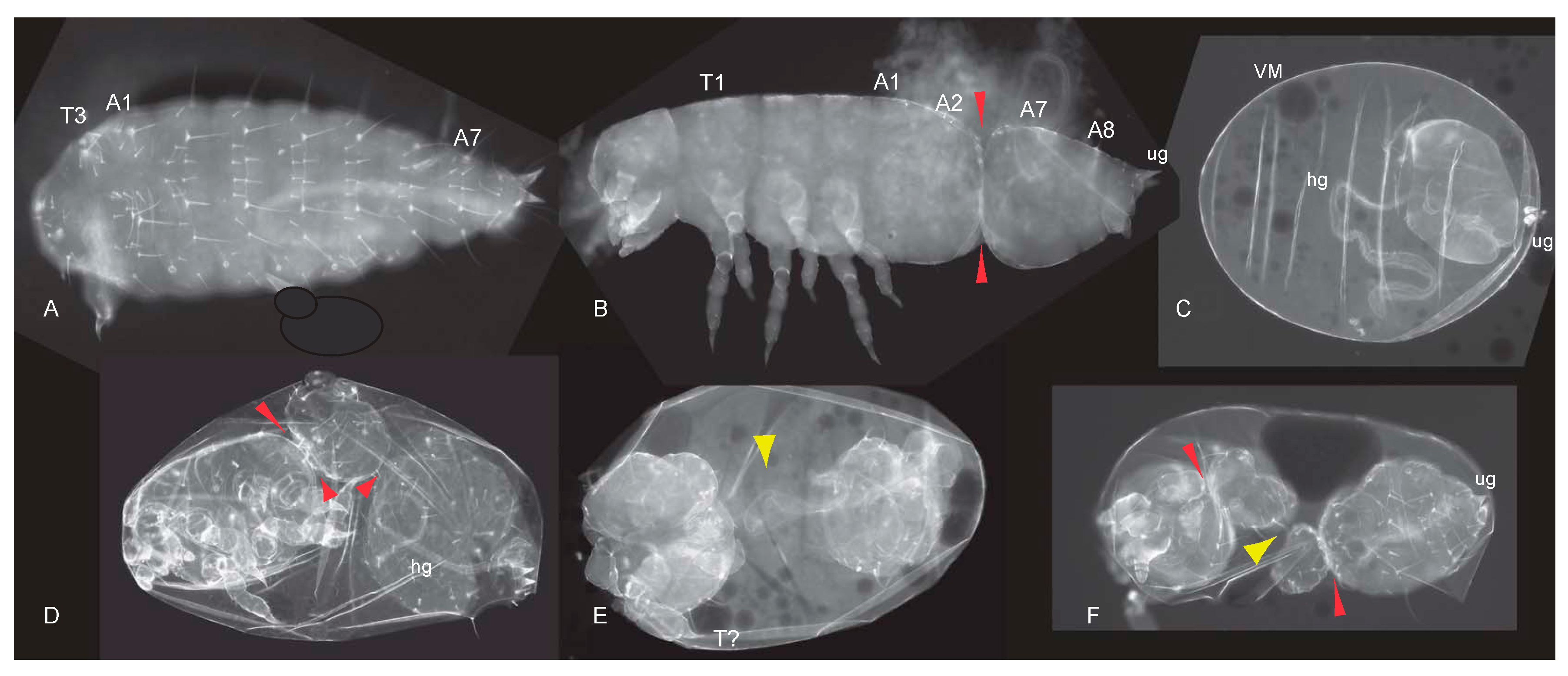
3.2. Reduction of Tc-axin Function Results in Anterior Truncated, Mild Bicaudal and Symmetric Double Abdomen Embryos
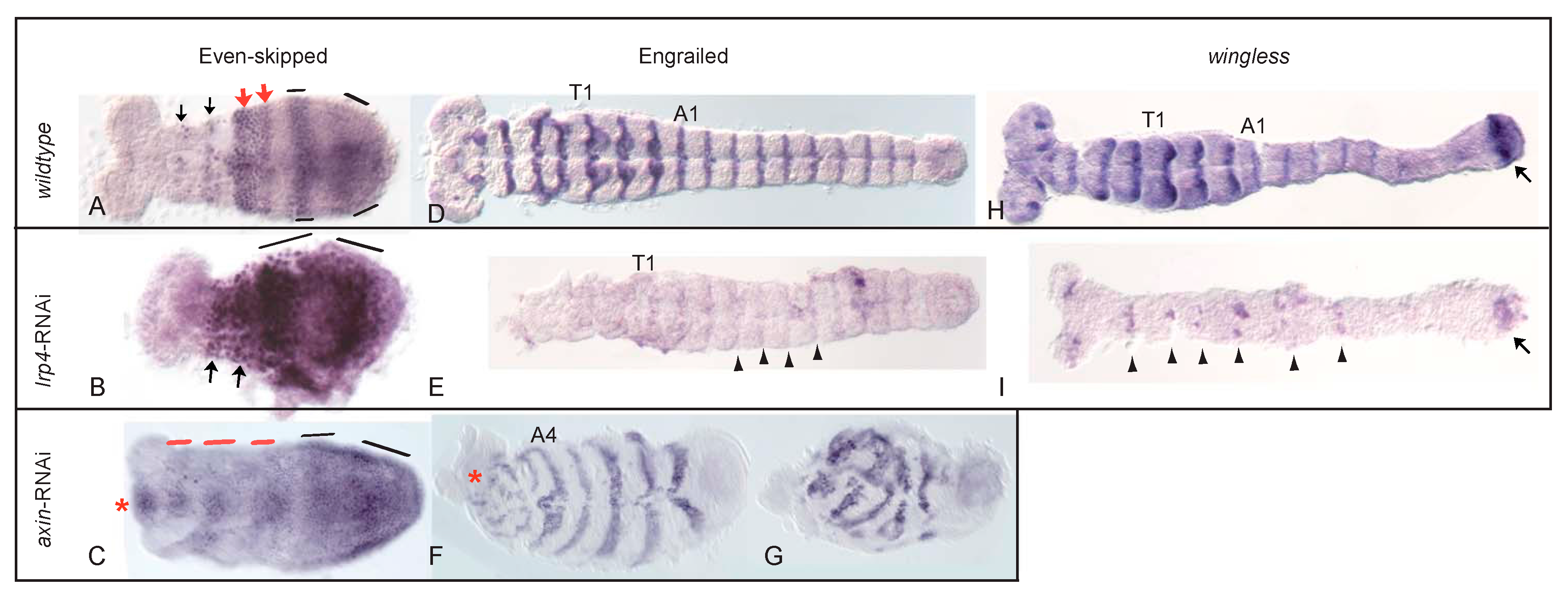
3.3. Structure, Expression and Functional Analysis of Tc-lrp4
3.4. Pair-Rule and Segment-Polarity Gene Expression in Tc-lrp4 RNAi Embryos Reveal Instability of the Segment Boundary
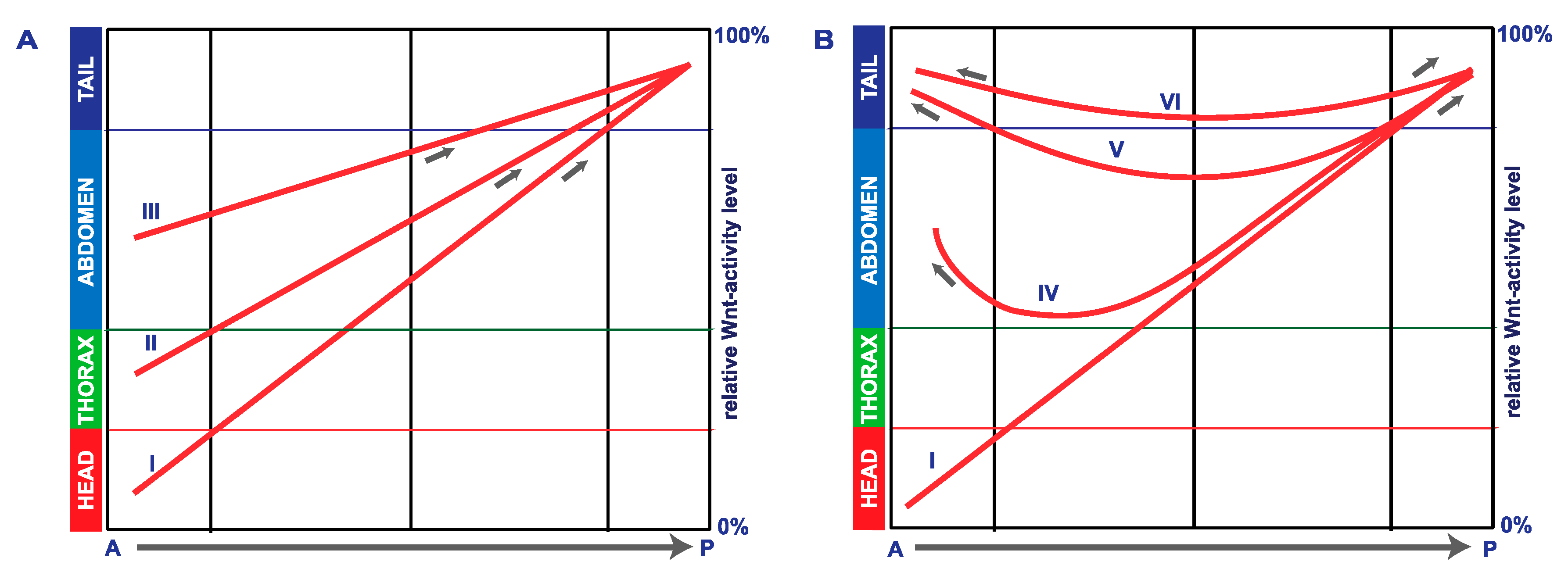
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Nusse, R.; Clevers, H. Wnt/beta-Catenin signaling, disease, and emerging therapeutic modalities. Cell 2017, 169, 985–999. [Google Scholar] [CrossRef] [PubMed]
- Loh, K.M.; van Amerongen, R.; Nusse, R. Generating Cellular Diversity and Spatial Form: Wnt Signaling and the Evolution of Multicellular Animals. Dev. Cell 2016, 38, 643–655. [Google Scholar] [CrossRef] [PubMed]
- Moon, R.T.; Kohn, A.D.; De Ferrari, G.V.; Kaykas, A. WNT and beta-catenin signalling: Diseases and therapies. Nat. Rev. Genet. 2004, 5, 691–701. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H.; Nusse, R. Wnt/beta-catenin signaling and disease. Cell 2012, 149, 1192–1205. [Google Scholar] [CrossRef] [PubMed]
- Lara-Castillo, N.; Johnson, M.L. LRP receptor family member associated bone disease. Rev. Endocr. Metab. Disord. 2015, 16, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Green, D.; Whitener, A.E.; Mohanty, S.; Lekven, A.C. Vertebrate nervous system posteriorization: Grading the function of Wnt signaling. Dev. Dyn. 2015, 244, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Niehrs, C. Regionally specific induction by the Spemann-Mangold organizer. Nat. Rev. Genet. 2004, 5, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Hikasa, H.; Sokol, S.Y. Wnt signaling in vertebrate axis specification. Cold Spring Harb. Perspect. Biol. 2013, 5. [Google Scholar] [CrossRef] [PubMed]
- Umesono, Y.; Tasaki, J.; Nishimura, Y.; Hrouda, M.; Kawaguchi, E.; Yazawa, S.; Nishimura, O.; Hosoda, K.; Inoue, T.; Agata, K. The molecular logic for planarian regeneration along the anterior-posterior axis. Nature 2013, 500, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Glinka, A.; Wu, W.; Onichtchouk, D.; Blumenstock, C.; Niehrs, C. Head induction by simultaneous repression of Bmp and Wnt signalling in Xenopus. Nature 1997, 389, 517–519. [Google Scholar] [PubMed]
- Niehrs, C. On growth and form: A Cartesian coordinate system of Wnt and BMP signaling specifies bilaterian body axes. Development 2010, 137, 845–857. [Google Scholar] [CrossRef] [PubMed]
- Bolognesi, R.; Beermann, A.; Farzana, L.; Wittkopp, N.; Lutz, R.; Balavoine, G.; Brown, S.J.; Schröder, R. Tribolium Wnts: Evidence for a larger repertoire in insects with overlapping expression patterns that suggest multiple redundant functions in embryogenesis. Dev. Genes Evol. 2008, 218, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Bolognesi, R.; Farzana, L.; Fischer, T.D.; Brown, S.J. Multiple Wnt genes are required for segmentation in the short-germ embryo of Tribolium castaneum. Curr. Biol. 2008, 18, 1624–1629. [Google Scholar] [CrossRef] [PubMed]
- Beermann, A.; Prühs, R.; Lutz, R.; Schröder, R. A context-dependent combination of Wnt receptors controls axis elongation and leg development in a short germ insect. Development 2011, 138, 2793–2805. [Google Scholar] [CrossRef] [PubMed]
- Bolognesi, R.; Fischer, T.D.; Brown, S.J. Loss of Tc-arrow and canonical Wnt signaling alters posterior morphology and pair-rule gene expression in the short-germ insect, Tribolium castaneum. Dev. Genes Evol. 2009, 219, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Posnien, N.; Bolognesi, R.; Fischer, T.D.; Rayl, P.; Oberhofer, G.; Kitzmann, P.; Brown, S.J.; Bucher, G. Asymmetrically expressed axin required for anterior development in Tribolium. Proc. Natl. Acad. Sci. USA 2012, 109, 7782–7786. [Google Scholar] [CrossRef] [PubMed]
- Mosca, T.J.; Luginbuhl, D.J.; Wang, I.E.; Luo, L.Q. Presynaptic LRP4 promotes synapse number and function of excitatory CNS neurons. eLife 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Liang, C.; Bates, R.; Yin, Y.M.; Xiong, W.C.; Mei, L. Wnt proteins regulate acetylcholine receptor clustering in muscle cells. Mol. Brain 2012, 5, 7. [Google Scholar] [CrossRef] [PubMed]
- Ahn, Y.; Sims, C.; Logue, J.M.; Weatherbee, S.D.; Krumlauf, R. Lrp4 and Wise interplay controls the formation and patterning of mammary and other skin appendage placodes by modulating Wnt signaling. Development 2013, 140, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.Y.; Dieckmann, M.; Herz, J.; Niemeier, A. Lrp4, a novel receptor for Dickkopf 1 and sclerostin, is expressed by osteoblasts and regulates bone growth and turnover in vivo. PLoS ONE 2009, 4, e7930. [Google Scholar] [CrossRef] [PubMed]
- Ahn, Y.; Sims, C.; Murray, M.J.; Kuhlmann, P.K.; Fuentes-Antras, J.; Weatherbee, S.D.; Krumlauf, R. Multiple modes of Lrp4 function in modulation of Wnt/beta-catenin signaling during tooth development. Development 2017, 144, 2824–2836. [Google Scholar] [CrossRef] [PubMed]
- Driever, W.; Nüsslein-Volhard, C. The Bicoid protein determines position in the Drosophila embryo in a concentration-dependent manner. Cell 1988, 54, 95–104. [Google Scholar] [CrossRef]
- Bucher, G.; Scholten, J.; Klingler, M. Parental RNAi in Tribolium (Coleoptera). Curr. Biol. 2002, 12, R85–R86. [Google Scholar] [CrossRef]
- Sarrazin, A.F.; Peel, A.D.; Averof, M. A segmentation clock with two-segment periodicity in insects. Science 2012, 336, 338–341. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Beermann, A.; Schröder, R. The dynamic expression of extraembryonic marker genes in the beetle Tribolium castaneum reveals the complexity of serosa and amnion formation in a short germ insect. Gene Expr. Patterns 2013, 13, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Schulz, C.; Schröder, R.; Hausdorf, B.; Wolff, C.; Tautz, D. A caudal homologue in the short germ band beetle Tribolium shows similarities to both, the Drosophila and the vertebrate caudal expression patterns. Dev. Genes Evol. 1998, 208, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Berns, N.; Kusch, T.; Schröder, R.; Reuter, R. Expression, function and regulation of Brachyenteron in the short germband insect Tribolium castaneum. Dev. Genes Evol. 2008, 218, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Patel, N.H.; Condron, B.G.; Zinn, K. Pair-rule expression patterns of even-skipped are found in both short- and long-germ beetles. Nature 1994, 367, 429–434. [Google Scholar] [CrossRef] [PubMed]
- Choe, C.P.; Miller, S.C.; Brown, S.J. A pair-rule gene circuit defines segments sequentially in the short-germ insect Tribolium castaneum. Proc. Natl. Acad. Sci. USA 2006, 103, 6560–6564. [Google Scholar] [CrossRef] [PubMed]
- Springer, T.A. An extracellular beta-propeller module predicted in lipoprotein and scavenger receptors, tyrosine kinases, epidermal growth factor precursor, and extracellular matrix components. J. Mol. Biol. 1998, 283, 837–862. [Google Scholar] [CrossRef] [PubMed]
- Itasaki, N.; Jones, C.M.; Mercurio, S.; Rowe, A.; Domingos, P.M.; Smith, J.C.; Krumlauf, R. Wise, a context-dependent activator and inhibitor of Wnt signalling. Development 2003, 130, 4295–4305. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, Y.; Kang, H.; Liu, W.; Liu, P.; Zhang, J.; Harris, S.E.; Wu, D. Sclerostin binds to LRP5/6 and antagonizes canonical Wnt signaling. J. Biol. Chem. 2005, 280, 19883–19887. [Google Scholar] [CrossRef] [PubMed]
- Mao, J.; Wang, J.; Liu, B.; Pan, W.; Farr, G.H., 3rd; Flynn, C.; Yuan, H.; Takada, S.; Kimelman, D.; Li, L.; et al. Low-density lipoprotein receptor-related protein-5 binds to Axin and regulates the canonical Wnt signaling pathway. Mol. Cell 2001, 7, 801–809. [Google Scholar] [CrossRef]
- Tamai, K.; Zeng, X.; Liu, C.; Zhang, X.; Harada, Y.; Chang, Z.; He, X. A mechanism for Wnt coreceptor activation. Mol. Cell 2004, 13, 149–156. [Google Scholar] [CrossRef]
- Petersen, C.P.; Reddien, P.W. Wnt signaling and the polarity of the primary body axis. Cell 2009, 139, 1056–1068. [Google Scholar] [CrossRef] [PubMed]
- Nakao, H. Anterior and posterior centers jointly regulate Bombyx embryo body segmentation. Dev. Biol. 2012, 371, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.A.; Brent, A.E.; Leaf, D.S.; Pultz, M.A.; Desplan, C. Localized maternal orthodenticle patterns anterior and posterior in the long germ wasp Nasonia. Nature 2006, 439, 728–732. [Google Scholar] [CrossRef] [PubMed]
- Schröder, R. The genes orthodenticle and hunchback substitute for bicoid in the beetle Tribolium. Nature 2003, 422, 621–625. [Google Scholar] [CrossRef] [PubMed]
- Kotkamp, K.; Klingler, M.; Schoppmeier, M. Apparent role of Tribolium orthodenticle in anteroposterior blastoderm patterning largely reflects novel functions in dorsoventral axis formation and cell survival. Development 2010, 137, 1853–1862. [Google Scholar] [CrossRef] [PubMed]
- Schinko, J.B.; Kreuzer, N.; Offen, N.; Posnien, N.; Wimmer, E.A.; Bucher, G. Divergent functions of orthodenticle, empty spiracles and buttonhead in early head patterning of the beetle Tribolium castaneum (Coleoptera). Dev. Biol. 2008, 317, 600–613. [Google Scholar] [CrossRef] [PubMed]
- Beermann, A.; Schröder, R. Sites of Fgf signalling and perception during embryogenesis of the beetle Tribolium castaneum. Dev. Genes Evol. 2008, 218, 153–167. [Google Scholar] [CrossRef] [PubMed]
- Schoppmeier, M.; Schröder, R. Maternal Torso signaling controls body axis elongation in a short germ insect. Curr. Biol. 2005, 15, 2131–2136. [Google Scholar] [CrossRef] [PubMed]
- Sander, K. Specification of the basic body pattern in insect embryogenesis. Adv. Insect Physiol. 1976, 12, 125–238. [Google Scholar]
- Lawrence, P.A. Background to bicoid. Cell 1988, 54, 1–2. [Google Scholar] [CrossRef]
- Zong, Y.; Zhang, B.; Gu, S.; Lee, K.; Zhou, J.; Yao, G.; Figueiredo, D.; Perry, K.; Mei, L.; Jin, R. Structural basis of agrin-LRP4-MuSK signaling. Genes Dev. 2012, 26, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Johnson, E.B.; Hammer, R.E.; Herz, J. Abnormal development of the apical ectodermal ridge and polysyndactyly in Megf7-deficient mice. Hum. Mol. Genet. 2005, 14, 3523–3538. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Luo, S.; Wang, Q.; Suzuki, T.; Xiong, W.C.; Mei, L. LRP4 serves as a coreceptor of agrin. Neuron 2008, 60, 285–297. [Google Scholar] [CrossRef] [PubMed]
- Lickert, H.; Domon, C.; Huls, G.; Wehrle, C.; Duluc, I.; Clevers, H.; Meyer, B.I.; Freund, J.N.; Kemler, R. Wnt/(beta)-catenin signaling regulates the expression of the homeobox gene Cdx1 in embryonic intestine. Development 2000, 127, 3805–3813. [Google Scholar] [PubMed]
- Pilon, N.; Oh, K.; Sylvestre, J.R.; Bouchard, N.; Savory, J.; Lohnes, D. Cdx4 is a direct target of the canonical Wnt pathway. Dev. Biol. 2006, 289, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Shinmyo, Y.; Mito, T.; Matsushita, T.; Sarashina, I.; Miyawaki, K.; Ohuchi, H.; Noji, S. Caudal is required for gnathal and thoracic patterning and for posterior elongation in the intermediate-germband cricket Gryllus bimaculatus. Mech. Dev. 2005, 122, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Choe, C.P.; Brown, S.J. Genetic regulation of engrailed and wingless in Tribolium segmentation and the evolution of pair-rule segmentation. Dev. Biol. 2009, 325, 482–491. [Google Scholar] [CrossRef] [PubMed]
- Banyai, L.; Sonderegger, P.; Patthy, L. Agrin binds BMP2, BMP4 and TGFbeta1. PLoS ONE 2010, 5, e10758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bull, A. Bicaudal, a genetic factor which affects the polarity of the embryo in Drosophila melanogaster. J. Exp. Zool. 1966, 161, 221–241. [Google Scholar] [CrossRef]
- Mohler, J.; Wieschaus, E.F. Dominant maternal-effect mutations of Drosophila melanogaster causing the production of double-abdomen embryos. Genetics 1986, 112, 803–822. [Google Scholar] [PubMed]
- Klomp, J.; Athy, D.; Kwan, C.W.; Bloch, N.I.; Sandmann, T.; Lemke, S.; Schmidt-Ott, U. Embryo development. A cysteine-clamp gene drives embryo polarity in the midge Chironomus. Science 2015, 348, 1040–1042. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}

| Numbers (N =) | axinRNAi_20 | axinRNAi_50 | iB-04108-1 | iB-04108-2 | axinRNAi GFP-Strain | axin/Wnt8 Double RNAi | axin/Arrow Double RNAi |
|---|---|---|---|---|---|---|---|
| WT | 93 | 91 | 44 | - | 167 | 126 | 25 |
| K-T-A | 15 | 8 | 4 | - | 7 | 19 | 1 |
| T-A (−) | 6 | - | 6 | - | 4 | 9 | 10 |
| A (−) | 7 | 5 | 15 | 1 | 9 | 12 | 34 |
| part. cuticle | 12 | 4 | 24 | 6 | 9 | 80 | 39 |
| T-A (+) | 5 | 7 | 8 | - | 3 | 5 | - |
| A (+) | 5 | 3 | 18 | 3 | 10 | 15 | 12 |
| A (+2 hg) | 9 | 6 | 11 | 11 | 6 | 62 | 21 |
| cuticles with polarity reversal | 19 | 16 | 37 | 14 | 19 | 82 | 33 |
| affected cuticles | 59 | 33 | 86 | 21 | 48 | 202 | 117 |
| all cuticles | 152 | 124 | 130 | 21 | 215 | 328 | 142 |
| empty eggs | 39 | 58 | 17 | 22 | 17 | 76 | 97 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prühs, R.; Beermann, A.; Schröder, R. The Roles of the Wnt-Antagonists Axin and Lrp4 during Embryogenesis of the Red Flour Beetle Tribolium castaneum. J. Dev. Biol. 2017, 5, 10. https://doi.org/10.3390/jdb5040010
Prühs R, Beermann A, Schröder R. The Roles of the Wnt-Antagonists Axin and Lrp4 during Embryogenesis of the Red Flour Beetle Tribolium castaneum. Journal of Developmental Biology. 2017; 5(4):10. https://doi.org/10.3390/jdb5040010
Chicago/Turabian StylePrühs, Romy, Anke Beermann, and Reinhard Schröder. 2017. "The Roles of the Wnt-Antagonists Axin and Lrp4 during Embryogenesis of the Red Flour Beetle Tribolium castaneum" Journal of Developmental Biology 5, no. 4: 10. https://doi.org/10.3390/jdb5040010
APA StylePrühs, R., Beermann, A., & Schröder, R. (2017). The Roles of the Wnt-Antagonists Axin and Lrp4 during Embryogenesis of the Red Flour Beetle Tribolium castaneum. Journal of Developmental Biology, 5(4), 10. https://doi.org/10.3390/jdb5040010