
Inhibition of Melanoma Cells A375 by Carotenoid Extract and Nanoemulsion Prepared from Pomelo Leaves



Abstract
:1. Introduction
2. Results and Discussion
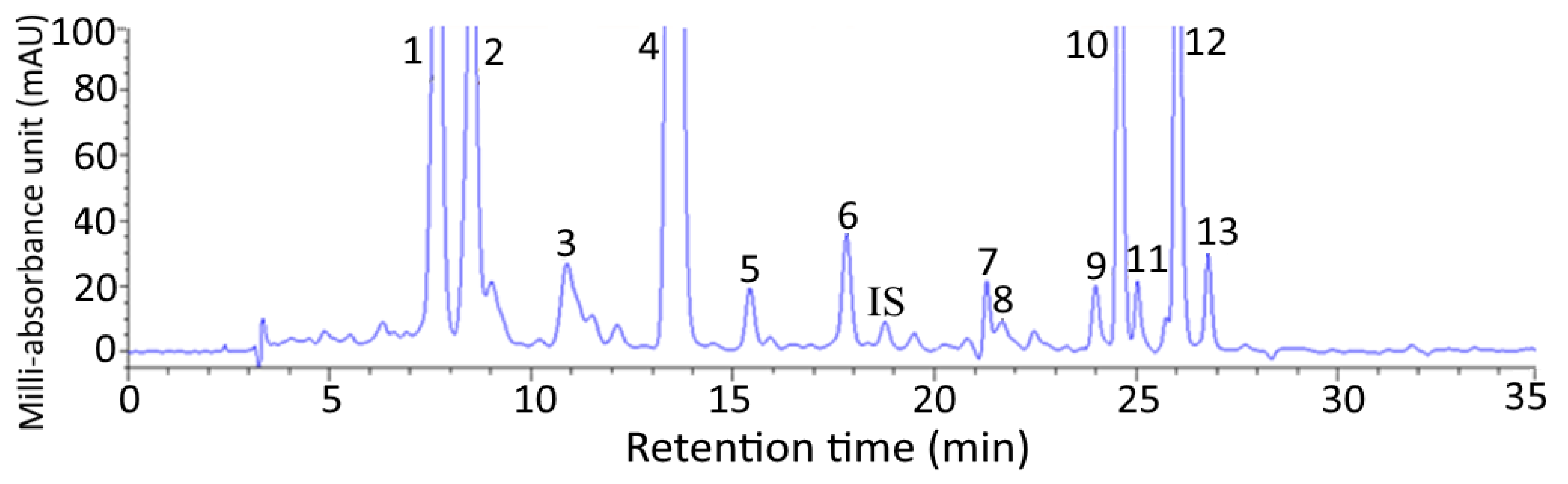
2.1. HPLC Analysis of Carotenoids in Pomelo Leaves
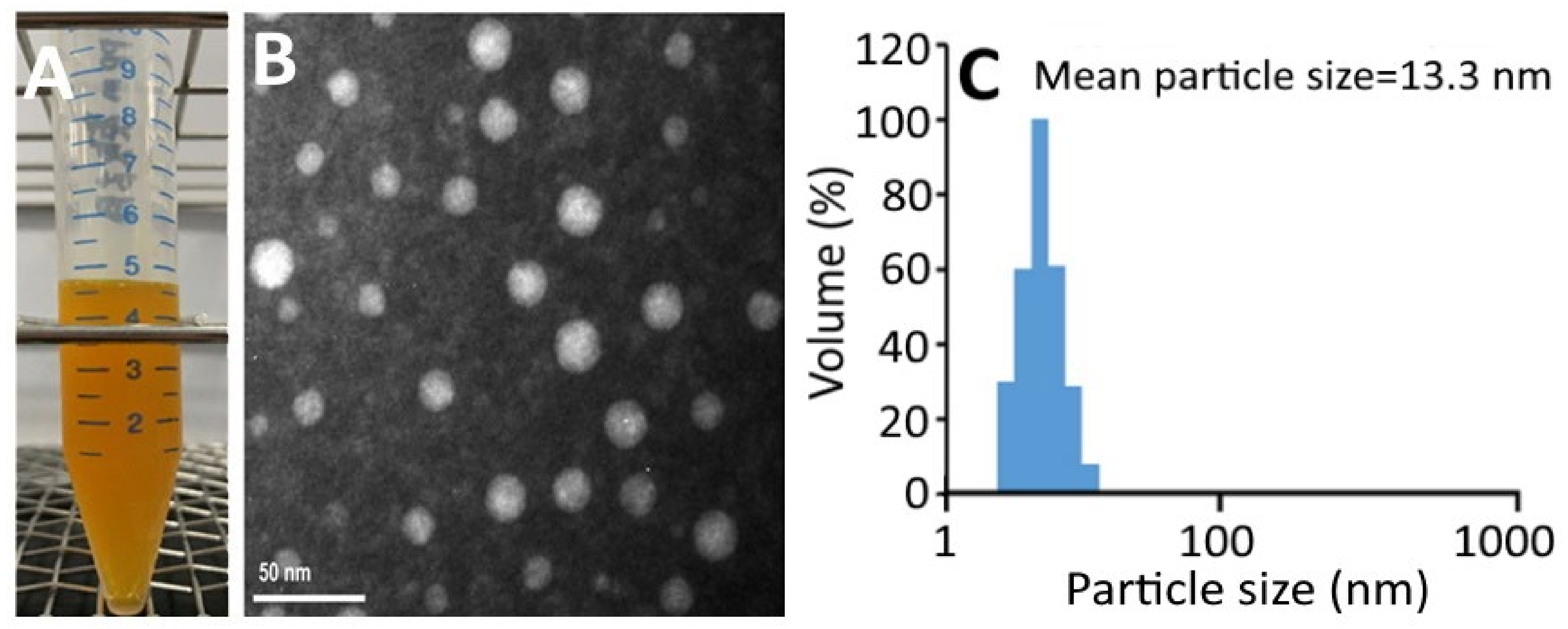
2.2. Nanoemulsion Characteristics
2.3. Tolerance of Sample Solvent and Blank Nanoemulsion towards Cells
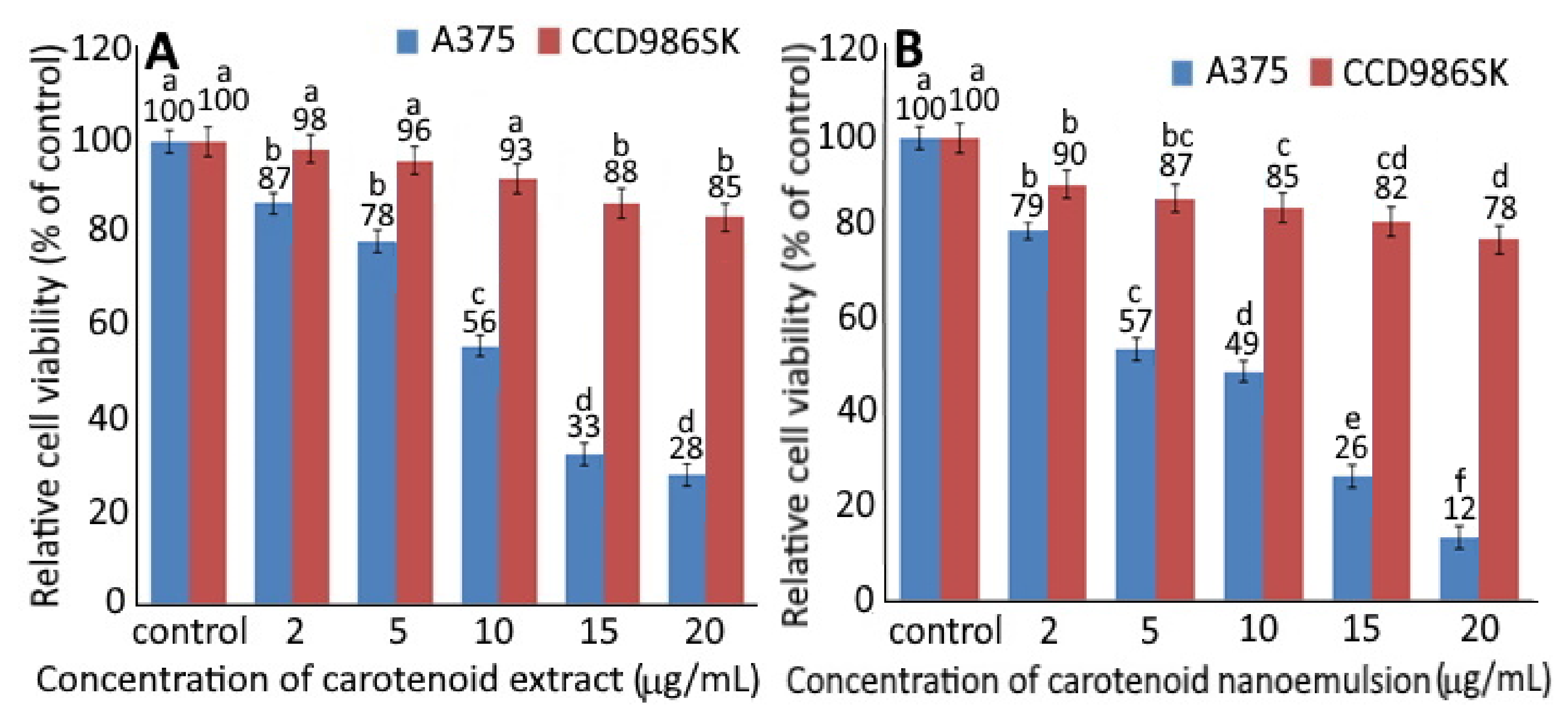
2.4. MTT Assay
2.5. Cell-Cycle Analysis
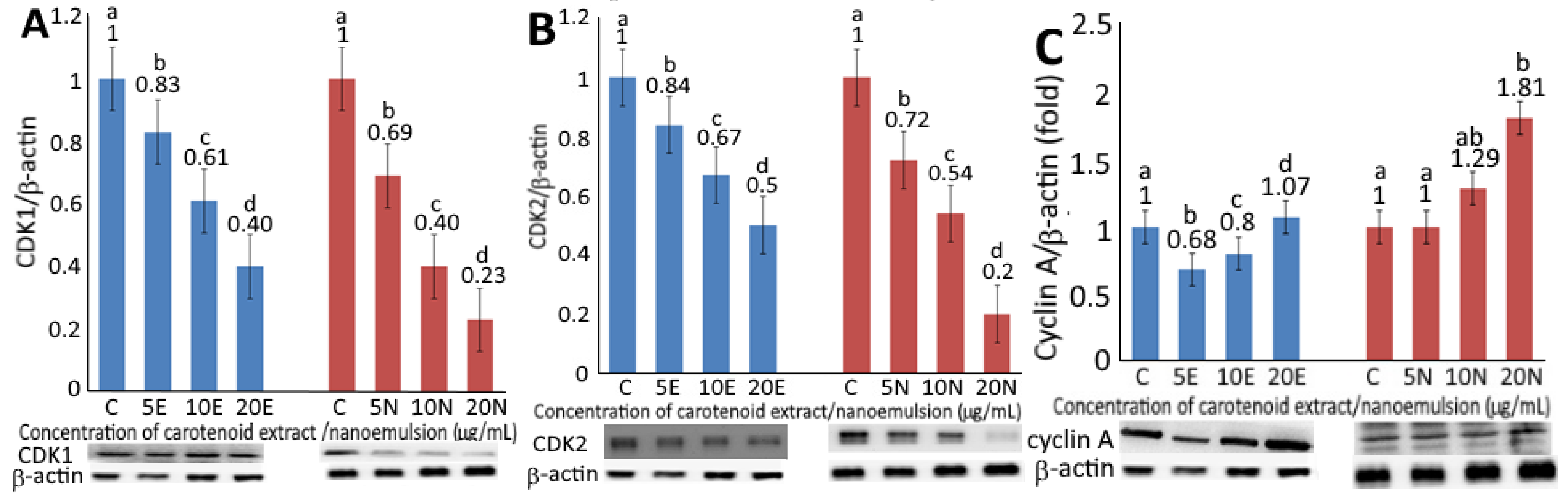
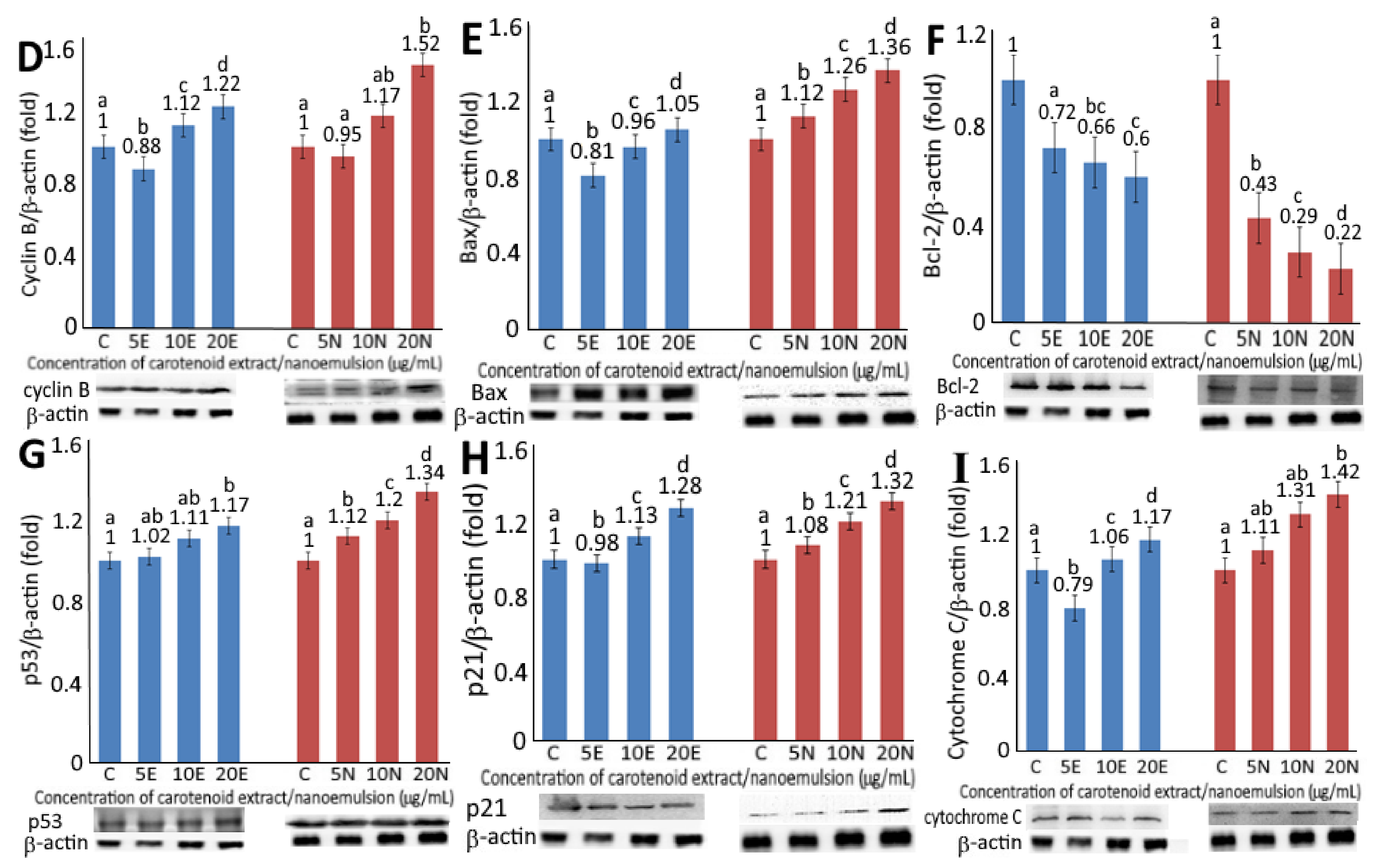
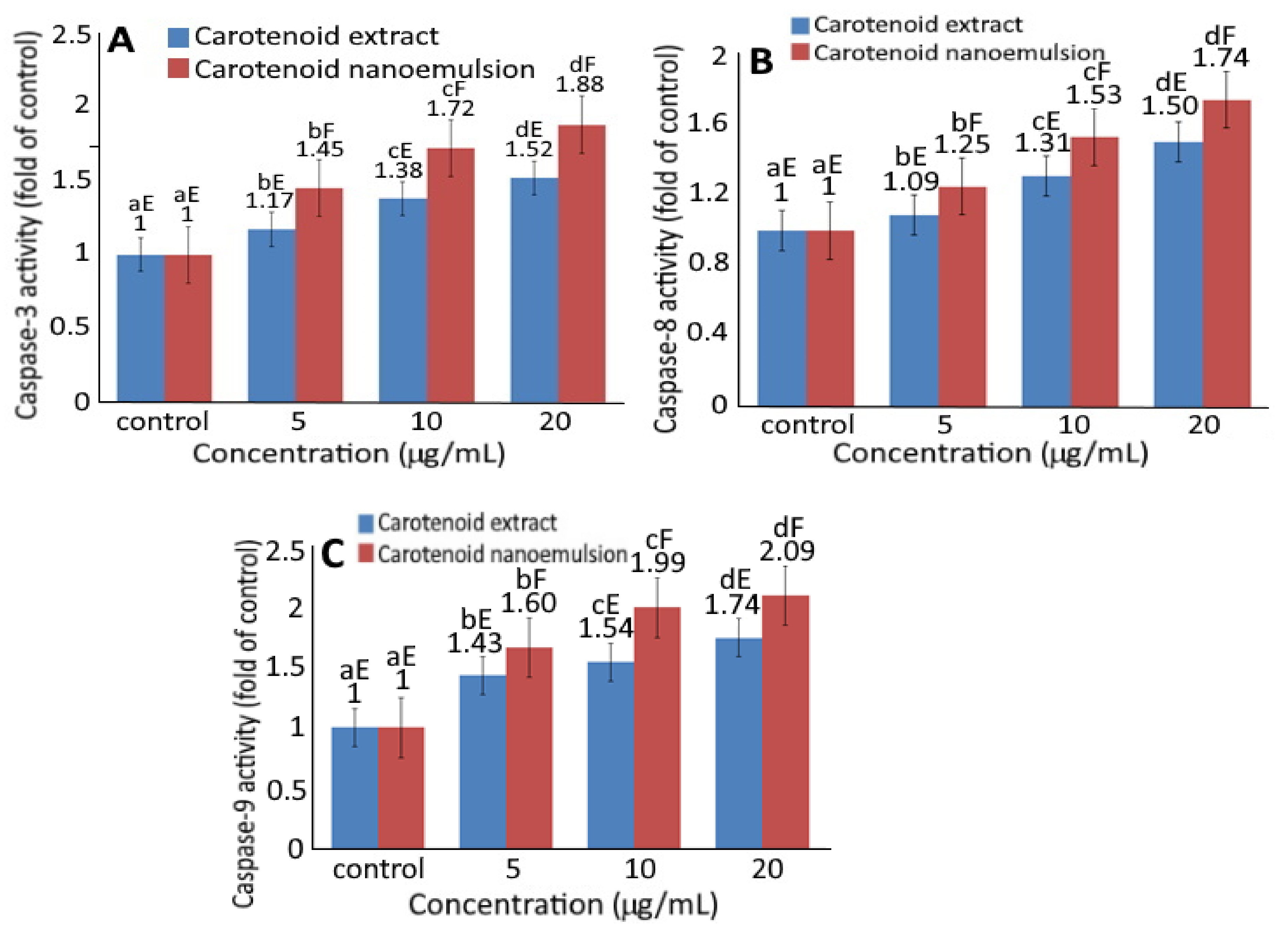
2.6. Expression of Cell-Cycle and Apoptosis-Related Protein
3. Materials and Methods
3.1. Materials
3.2. Cell Culture and Reagents
3.3. Western Blotting Antibody
3.4. Instrumentation
3.5. Extraction of Carotenoids from Pomelo Leaves
3.6. Preparation of Carotenoids from Pomelo Leaves by Open-Column Chromatography
3.7. HPLC Analysis of Carotenoids
3.8. Preparation of Carotenoid Nanoemulsion
3.9. Determination of Carotenoid Nanoemulsion Characteristics
3.10. Stability of Carotenoid Nanoemulsion
3.11. Cell Culture
3.12. MTT Assay
3.13. Cell-Cycle Analysis
3.14. Western Blotting
3.15. Activities of Caspase-3, -8 and -9
3.16. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Ruiz-Iglesias, P.; Estruel-Amades, S.; Camps-Bossacoma, M.; Massot-Cladera, M.; Franch, À.; Pérez-Cano, F.J.; Castell, M. Influence of hesperidin on systemic immunity of rats following an intensive training and exhausting exercise. Nutrients 2020, 12, 1291. [Google Scholar] [CrossRef] [PubMed]
- Ho, N.-H.; Inbaraj, B.S.; Chen, B.-H. Utilization of microemulsions from Rhinacanthus nasutus (L.) Kurz to improve carotenoid bioavailability. Sci. Rep. 2016, 6, 25426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perveen, R.; Suleria, H.A.R.; Anjum, F.M.; Butt, M.S.; Pasha, I.; Ahmad, S. Tomato (Solanum lycopersicum) carotenoids and lycopenes chemistry; metabolism, absorption, nutrition, and allied health claims—A comprehensive review. Crit. Rev. Food Sci. Nutr. 2015, 55, 919–929. [Google Scholar] [CrossRef] [PubMed]
- Milani, A.; Basirnejad, M.; Shahbazi, S.; Bolhassani, A. Carotenoids: Biochemistry, pharmacology and treatment. Br. J. Pharmacol. 2017, 174, 1290–1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.-M.; Yen, Y.-T.; Huang, C.-S.; Hu, M.-L. Growth inhibitory efficacy of lycopene and β-carotene against androgen-independent prostate tumor cells xenografted in nude mice. Mol. Nutr. Food Res. 2011, 55, 606–612. [Google Scholar] [CrossRef]
- Gloria, N.F.; Soares, N.; Brand, C.; Oliveira, F.L.; Borojevic, R.; Teodoro, A.J. Lycopene and beta-carotene induce cell-cycle arrest and apoptosis in human breast cancer cell lines. Anticancer Res. 2014, 34, 1377–1386. [Google Scholar]
- Huang, R.F.; Wei, Y.J.; Inbaraj, B.S.; Chen, B.H. Inhibition of colon cancer cell growth by nanoemulsion carrying gold nanoparticles and lycopene. Int. J. Nanomed. 2015, 10, 2823–2846. [Google Scholar] [CrossRef] [Green Version]
- Hsu, H.J.; Huang, R.F.; Kao, T.H.; Inbaraj, B.S.; Chen, B.H. Preparation of carotenoid extracts and nanoemulsions from Lycium barbarum L. and their effects on growth of HT-29 colon cancer cells. Nanotechnology 2017, 28, 135103. [Google Scholar] [CrossRef]
- Sridhar, K.; Inbaraj, B.S.; Chen, B.-H. Recent advances on nanoparticle based strategies for improving carotenoid stability and biological activity. Antioxidants 2021, 10, 713. [Google Scholar] [CrossRef]
- McClements, D.J. Nanoemulsions versus microemulsions: Terminology, differences, and similarities. Soft Matter 2012, 8, 1719–1729. [Google Scholar] [CrossRef]
- Lin, B.-J.; Chen, W.-H.; Budzianowski, W.M.; Hsieh, C.-T.; Lin, P.-H. Emulsification analysis of bio-oil and diesel under various combinations of emulsifiers. Appl. Energy 2016, 178, 746–757. [Google Scholar] [CrossRef]
- Bonifácio, B.V.; Silva, P.B.d.; Ramos, M.A.D.S.; Negri, K.M.S.; Bauab, T.M.; Chorilli, M. Nanotechnology-based drug delivery systems and herbal medicines: A review. Int. J. Nanomed. 2014, 9, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Inbaraj, B.S.; Lu, H.; Hung, C.F.; Wu, W.B.; Lin, C.L.; Chen, B.H. Determination of carotenoids and their esters in fruits of Lycium barbarum Linnaeus by HPLC–DAD–APCI–MS. J. Pharm. Biomed. Anal. 2008, 47, 812–818. [Google Scholar] [CrossRef] [PubMed]
- De Faria, A.F.; Hasegawa, P.N.; Chagas, E.A.; Pio, R.; Purgatto, E.; Mercadante, A.Z. Cultivar influence on carotenoid composition of loquats from Brazil. J. Food Compos. Anal. 2009, 22, 196–203. [Google Scholar] [CrossRef]
- Delgado-Pelayo, R.; Gallardo-Guerrero, L.; Hornero-Méndez, D. Chlorophyll and carotenoid pigments in the peel and flesh of commercial apple fruit varieties. Food Res. Int. 2014, 65, 272–281. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.-L.; Chen, B.-H.; Kao, T.-H.; Shiau, C.-Y. Carotenoids composition in Scutellaria barbata D. Don as detected by high performance liquid chromatography-diode array detection-mass spectrometry-atmospheric pressure chemical ionization. J. Funct. Foods 2014, 8, 100–110. [Google Scholar] [CrossRef]
- Song, J.; Li, D.; He, M.; Chen, J.; Liu, C. Comparison of carotenoid composition in immature and mature grains of corn (Zea mays L.) varieties. Int. J. Food Prop. 2016, 19, 351–358. [Google Scholar] [CrossRef]
- Schex, R.; Lieb, V.M.; Jiménez, V.M.; Esquivel, P.; Schweiggert, R.M.; Carle, R.; Steingass, C.B. HPLC-DAD-APCI/ESI-MSn analysis of carotenoids and α-tocopherol in Costa Rican Acrocomia aculeata fruits of varying maturity stages. Food Res. Int. 2018, 105, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Petry, F.C.; Mercadante, A.Z. New method for carotenoid extraction and analysis by HPLC-DAD-MS/MS in freeze-dried citrus and mango pulps. J. Braz. Chem. Soc. 2018, 29, 205–215. [Google Scholar] [CrossRef]
- Yang, C.C.; Hung, C.F.; Chen, B.H. Preparation of coffee oil-algae oil-based nanoemulsions and the study of their inhibition effect on UVA-induced skin damage in mice and melanoma cell growth. Int. J. Nanomed. 2017, 12, 6559–6580. [Google Scholar] [CrossRef] [Green Version]
- Qian, C.; Decker, E.A.; Xiao, H.; McClements, D.J. Physical and chemical stability of β-carotene-enriched nanoemulsions: Influence of pH, ionic strength, temperature, and emulsifier type. Food Chem. 2012, 132, 1221–1229. [Google Scholar] [CrossRef]
- Liu, M.H.; Li, Y.F.; Chen, B.H. Preparation of chlorophyll nanoemulsion from pomelo leaves and tts inhibition effect on melanoma cells A375. Plants 2021, 10, 1664. [Google Scholar] [CrossRef]
- Dutta, S.; Surapaneni, B.K.; Bansal, A. Marked Inhibition of Cellular Proliferation in the normal human esophageal epithelial cells and human esophageal squamous cancer cells in culture by carotenoids: Role for prevention and early treatment of esophageal cancer. Asian Pac. J. Cancer Prev. APJCP 2018, 19, 3251–3256. [Google Scholar] [CrossRef]
- Wang, S.K.; Yang, L.; Wang, T.T.; Huang, G.L.; Yang, L.G.; Sun, G.J. Inhibition of proliferation and induction of apoptosis by the combination of β-carotene and 1,25-dihydroxyvitamin D3 in human esophageal cancer EC9706 cells. Asian Pac. J. Cancer Prev. APJCP 2012, 13, 6327–6332. [Google Scholar] [CrossRef] [Green Version]
- Albino, A.P.; Juan, G.; Traganos, F.; Reinhart, L.; Connolly, J.; Rose, D.P.; Darzynkiewicz, Z. Cell cycle arrest and apoptosis of melanoma cells by docosahexaenoic acid: Association with decreased pRb phosphorylation. Cancer Res. 2000, 60, 4139. [Google Scholar] [PubMed]
- Mohd, M.; Saima, W. Herbal based polymeric nanoparticles as a therapeutic remedy for breast cancer. Anti-Cancer Agents Med. Chem. 2021, 21, 433–444. [Google Scholar]
- Koklesova, L.; Liskova, A.; Samec, M.; Buhrmann, C.; Samuel, S.M.; Varghese, E.; Ashrafizadeh, M.; Najafi, M.; Shakibaei, M.; Büsselberg, D.; et al. Carotenoids in Cancer Apoptosis—The Road from Bench to Bedside and Back. Cancers 2020, 12, 2425. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Liu, W.; Zhou, X.; Long, C.; Kuang, X.; Hu, J.; Tang, Y.; Liu, L.; He, J.; Huang, Z.; et al. Protective effect of lutein on ARPE-19 cells upon H2O2-induced G2/M arrest. Mol. Med. Rep. 2017, 16, 2069–2074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, J.; Song, M.-H.; Oh, J.-W.; Keum, Y.-S.; Saini, R.K. Pro-oxidant actions of carotenoids in triggering apoptosis of cancer cells: A review of emerging evidence. Antioxidants 2020, 9, 532. [Google Scholar] [CrossRef]
- Hwang, D.; Kim, M.; Park, H.; Jeong, M.I.; Jung, W.; Kim, B. Natural products and acute myeloid leukemia: A review highlighting mechanisms of action. Nutrients 2019, 11, 1010. [Google Scholar] [CrossRef] [Green Version]
- SAS. SAS Procedures and SAS/Graph User’s Guide; Version 6; SAS Institute Inc.: Gary, NC, USA, 2014. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No. | Compound | tR (min) | Retention Factor (k) a | Separation Factor (α) b | Content (μg/g) d | m/z Found | m/z Reported | λmax (nm, Online) j | λmax (nm, Reported) j | Q-Ratio Found | Q-Ratio Reported |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | All-trans-neoxanthin | 7.67 | 1.28 | 1.20 (1,2) c | 309.2 | 601.5 [M+H], 583 [M+H-18] | 601.5 [M+H], 583 [M+H-18] f | 328, 418, 440, 470 | 330, 416, 442, 470 k | 0.08 | - |
| 2 | All-trans-violaxanhin | 8.53 | 1.53 | 1.47 (2,3) | 208.5 | 601.5 [M+H], 583 [M+H-18] | 601.5 [M+H], 583 [M+H-18] f | 328, 412, 436, 464 | 329, 416, 439, 469 g | 0.07 | - |
| 3 | 13-or 13’-cis- lutein | 10.88 | 2.25 | 1.17 (3,4) | 81.04 | 551.5 [M+H-18] | 551.5 [M+H-18] g | 334, 426, 448, 470 | 334, 418, 441, 470 g | 0.35 | 0.38 l |
| 4 | All-trans-lutein | 12.13 | 2.62 | 1.16 (4,5) | 2922.3 | 551.4 [M+H-18] | 551.4 [M+H-18] g | 332, 440, 466 | 332, 445, 473 g | 0.07 | 0.07 l |
| 5 | 9-or 9’cis-lutein | 13.54 | 3.04 | 1.18 (5,6) | 9.63 | 551.4 [M+H], 533.4 [M+H-18-18] | 551.4 [M+H], 533.4 [M+H-18-18] g | 332, 446, 474 | 332, 440, 468 g | 0.06 | 0.08 l |
| 6 | All-trans-zeaxanthin | 17.81 | 3.60 | 1.20 (6,IS) | 54.67 | 569.4 [M+H], 551.4 [M+H-18] | 569.4 [M+H],551.4 [M+H-18] g | 340, 452, 478 | 339, 452, 478 k | 0.07 | 0.06 l |
| IS e | All-trans-canthaxanthin | 19.21 | 4.32 | 1.24 (IS,7) | - | - | 478 | - | - | - | |
| 7 | All-trans-β-cryptoxanthin | 21.29 | 5.36 | 1.02 (7,8) | 18.63 | 553.4 [M+H], 535.4 [M+H-18] | 553.4 [M+H], 535.4 [M+H-18] h | 456, 482 | 428, 450, 477 k | 0.14 | 0.16 m |
| 8 | 9-or 9’-cis-β-cryptoxanthin | 21.66 | 5.47 | 1.13 (8,9) | 5.93 | 553.4 [M+H] | 553.4 [M+H] h | 336, 442, 468 | 332, 410, 440, 468 k | 0.12 | 0.11 m |
| 9 | 13-or 13’-cis-β -carotene | 23.99 | 6.16 | 1.03 (9,10) | 12.64 | 537.4 [M+H] g | 537.4 [M+H] g | 342, 448 | 332, 416, 440, 470 k | 0.42 | 0.43 m |
| 10 | All-trans-α-carotene | 24.56 | 6.33 | 1.02 (10,11) | 142.6 | 537.3 [M+H] | 537.3 [M+H] i | 450, 478 | 338, 418, 440, 472 k | 0.08 | - |
| 11 | 9-or 9’-cis-α-carotene | 25.03 | 6.47 | 1.05 (11,12) | 9.9 | 537.3 [M+H] | - | 444, 472 | 440, 468 k | 0.10 | - |
| 12 | All-trans-β-carotene | 26.01 | 6.77 | 1.03 (12,13) | 168.3 | 537.4 [M+H] | 537.4 [M+H] g | 456, 482 | 442, 452, 476 k | 0.08 | 0.09 m |
| 13 | 9-or 9’-cis-β-carotene | 26.78 | 7.00 | 1.03 (13,14) | 22.23 | 537.4 [M+H] | 537.4 [M+H] g | 452, 476 | 446, 448, 472 k | 0.13 | 0.13 m |
| Concentration (µg/mL) | Sub-G1 (%) | G0/G1 (%) | S (%) | G2/M (%) |
|---|---|---|---|---|
| control | 3.310.06 B | 52.83 A | 17.65 A | 26.27 A |
| carotenoid extract | ||||
| 5 | 4.03 B | 49.36 B | 16.23 B | 28.15 B |
| 10 | 4.25 B | 47.78 C | 15.86 B | 31.36 C |
| 20 | 3.91 B | 46.48 C | 15.23 B | 35.27 D |
| carotenoid nanoemulsion | ||||
| 5 | 4.36 B | 45.27 C | 15.57 B | 32.86 C |
| 10 | 4.18 B | 44.53 C | 16.14 B | 36.72 D |
| 20 | 4.98 B | 42.35 D | 14.92 C | 41.95 E |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, M.-H.; Li, Y.-F.; Chen, B.-H. Inhibition of Melanoma Cells A375 by Carotenoid Extract and Nanoemulsion Prepared from Pomelo Leaves. Plants 2021, 10, 2129. https://doi.org/10.3390/plants10102129
Liu M-H, Li Y-F, Chen B-H. Inhibition of Melanoma Cells A375 by Carotenoid Extract and Nanoemulsion Prepared from Pomelo Leaves. Plants. 2021; 10(10):2129. https://doi.org/10.3390/plants10102129
Chicago/Turabian StyleLiu, Man-Hai, Yi-Fen Li, and Bing-Huei Chen. 2021. "Inhibition of Melanoma Cells A375 by Carotenoid Extract and Nanoemulsion Prepared from Pomelo Leaves" Plants 10, no. 10: 2129. https://doi.org/10.3390/plants10102129
APA StyleLiu, M. -H., Li, Y. -F., & Chen, B. -H. (2021). Inhibition of Melanoma Cells A375 by Carotenoid Extract and Nanoemulsion Prepared from Pomelo Leaves. Plants, 10(10), 2129. https://doi.org/10.3390/plants10102129