
Wheat and Cereal Rye Inter-Row Living Mulches Interfere with Early Season Weeds in Soybean


Abstract
:1. Introduction
2. Materials and Methods
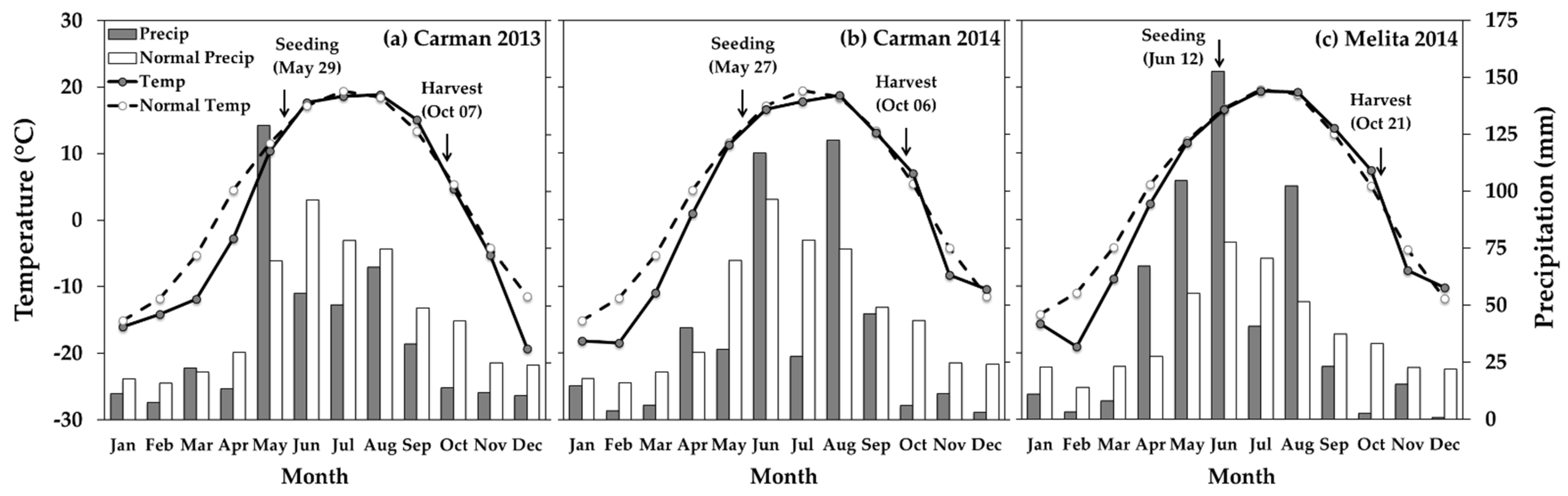
2.1. Experimental Sites
2.2. Experimental Design and Treatment Structure
2.3. Data Collection
2.3.1. Soybean
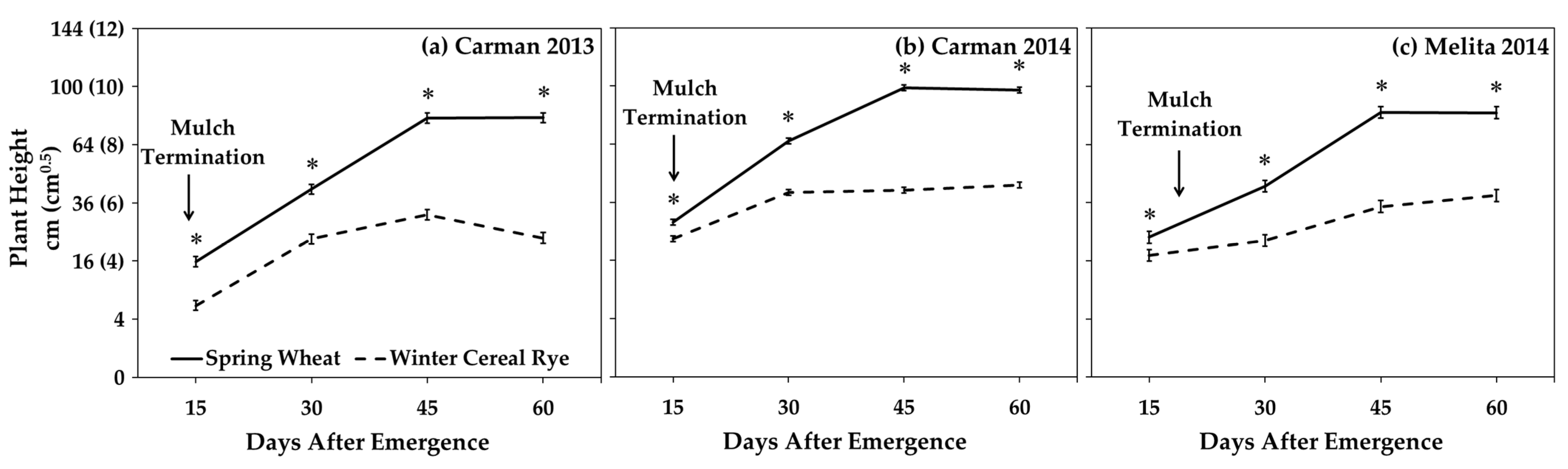
2.3.2. Inter-Row Mulches
2.3.3. Volunteer Canola
2.4. Statistical Analysis
3. Results
3.1. Soybean
3.2. Volunteer Canola
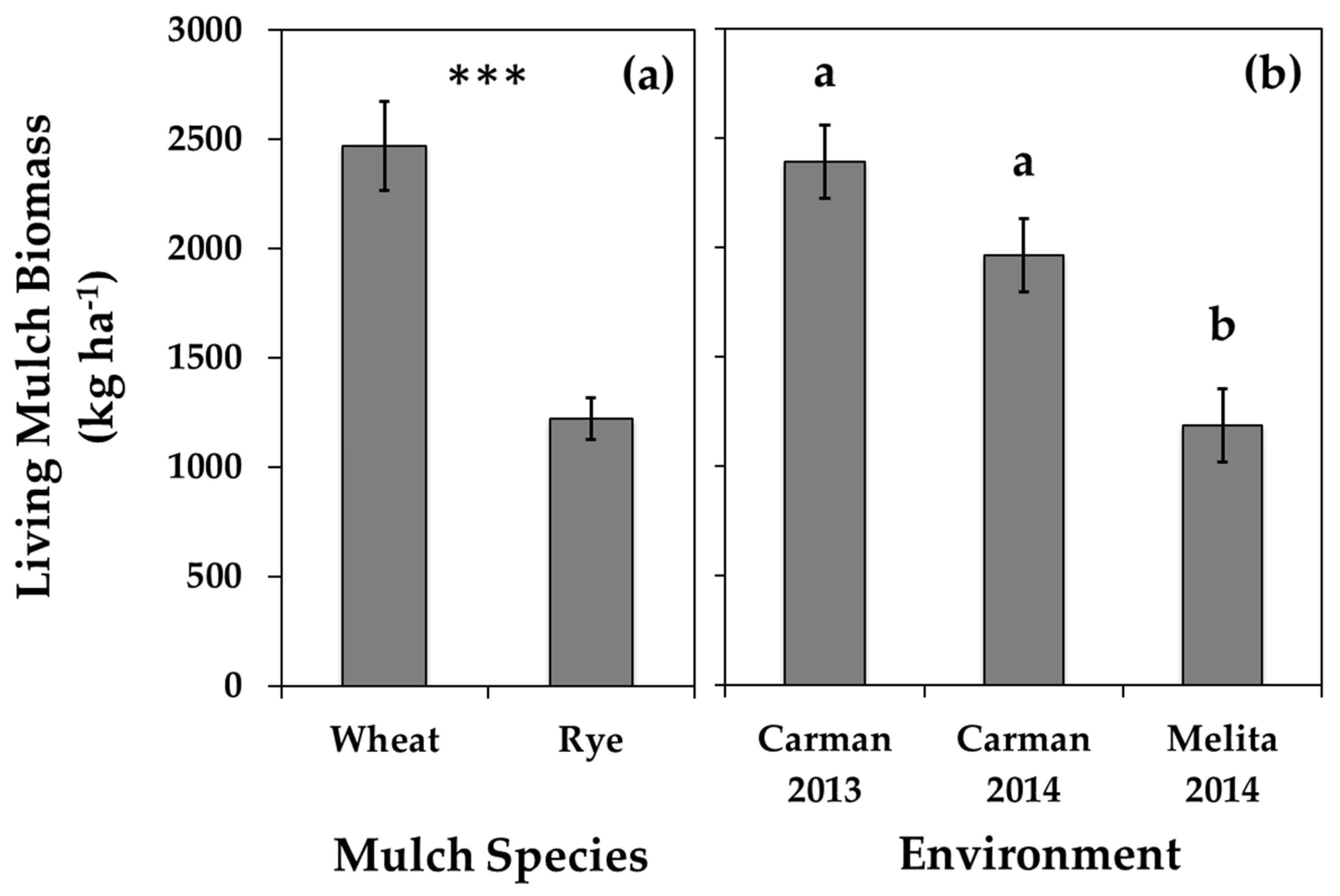
3.3. Inter-Row Mulches
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Geddes, C.M.; Gulden, R.H. Candidate tools for integrated weed management in soybean at the northern frontier of production. Weed Sci. 2018, 66, 662–672. [Google Scholar] [CrossRef]
- Mierau, A.; Kurtenbach, M.E.; Johnson, E.N.; Gulden, R.H.; Weber, J.D.; May, W.E.; Willenborg, C.J. Herbicide programs for control of glyphosate-resistant canola (Brassica napus) in glyphosate-resistant soybean. Weed Technol. 2020, 34, 540–546. [Google Scholar] [CrossRef]
- Statistics Canada. Table 32-10-0359-01—Estimated Areas, Yield, Production, Average Farm Price and Total Farm Value of Principal Field Crops, in Metric and Imperial Units (Database). Available online: www.statcan.gc.ca (accessed on 26 June 2021).
- Cavalieri, A.; Harker, K.N.; Hall, L.M.; Gulden, R.H. Evaluation of the causes of on-farm harvest losses in canola in the northern Great Plains. Crop Sci. 2016, 56, 2005–2015. [Google Scholar] [CrossRef]
- Gulden, R.H.; Shirtliffe, S.J.; Thomas, A.G. Secondary seed dormancy prolongs persistence of volunteer canola in western Canada. Weed Sci. 2003, 51, 904–913. [Google Scholar] [CrossRef]
- Haile, T.A.; Holzapfel, C.B.; Shirtliffe, S.J. Canola genotypes and harvest methods affect seedbank addition. Agron. J. 2014, 106, 236–242. [Google Scholar] [CrossRef]
- Geddes, C.M.; Gulden, R.H. Early autumn soil disturbance decreases persistence of volunteer summer-annual oilseed rape (Brassica napus). Weed Res. 2017, 57, 182–192. [Google Scholar] [CrossRef]
- Gregoire, P.; Rosset, J.D.; Gulden, R.H. Volunteer Brassica napus (L.) interference with soybean [Glycine max (L.) Merr.]: Management thresholds, plant growth, and seed return. Can. J. Plant Sci. 2021, 101, 556–567. [Google Scholar] [CrossRef]
- Beckie, H.J.; Warwick, S.I. Persistence of an oilseed rape transgene in the environment. Crop Prot. 2010, 29, 509–512. [Google Scholar] [CrossRef]
- Hartwig, N.L.; Ammon, H.U. Cover crops and living mulches. Weed Sci. 2002, 50, 688–699. [Google Scholar] [CrossRef]
- Teasdale, J.R. Contribution of cover crops to weed management in sustainable agricultural systems. J. Prod. Agric. 1996, 9, 475–479. [Google Scholar] [CrossRef]
- Norsworthy, J.K.; McClelland, M.; Griffith, G.; Bangarwa, S.K.; Still, K. Evaluation of cereal and Brassicaceae cover crops in conservation-tillage, enhanced, glyphosate-resistant cotton. Weed Technol. 2011, 25, 6–13. [Google Scholar] [CrossRef]
- Price, A.J.; Balkcom, K.S.; Duzy, L.M.; Kelton, J.A. Herbicide and cover crop residue integration for Amaranthus control in conservation agriculture cotton and implications for resistance management. Weed Technol. 2012, 26, 490–498. [Google Scholar] [CrossRef]
- Wiggins, M.S.; Hayes, R.M.; Steckel, L.E. Evaluating cover crops and herbicides for glyphosate-resistant palmer amaranth (Amaranthus palmeri) control in cotton. Weed Technol. 2016, 30, 415–422. [Google Scholar] [CrossRef]
- Bhaskar, V.; Westbrook, A.S.; Bellinder, R.R.; DiTommaso, A. Integrated management of living mulches for weed control: A review. Weed Technol. 2021, in press. [Google Scholar] [CrossRef]
- Liebman, M.; Dyck, E. Crop rotation and intercropping strategies for weed management. Ecol. Appl. 1993, 3, 92–122. [Google Scholar] [CrossRef]
- Harper, J.L. Population Biology of Plants, 4th ed.; Academic Press: New York, NY, USA, 1977; p. 306. [Google Scholar]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S.; Vivanco, J.M. The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barney, J.N.; Sparks, J.P.; Greenberg, J.; Whitlow, T.H.; Guenther, A. Biogenic volatile organic compounds from an invasive species: Impacts on plant-plant interactions. Plant Ecol. 2009, 203, 195–205. [Google Scholar] [CrossRef] [Green Version]
- Krogh, S.S.; Mensz, S.J.M.; Nielsen, S.T.; Mortensen, A.G.; Christophersen, C.; Fomsgaard, I.S. Fate of benzoxazinone allelochemicals in soil after incorporation of wheat and rye sprouts. J. Agric. Food Chem. 2006, 54, 1064–1074. [Google Scholar] [CrossRef]
- Inderjit. Soil microorganisms: An important determinant of allelopathic activity. Plant Soil 2005, 274, 227–236. [Google Scholar] [CrossRef]
- Geddes, C.M.; Cavalieri, A.; Daayf, F.; Gulden, R.H. The allelopathic potential of hairy vetch (Vicia villosa Roth.) mulch. Am. J. Plant Sci. 2015, 6, 2651–2663. [Google Scholar] [CrossRef] [Green Version]
- Ateh, C.M.; Doll, J.D. Spring-planted winter rye (Secale cereale) as a living mulch to control weeds in soybean (Glycine max). Weed Technol. 1996, 10, 347–353. [Google Scholar] [CrossRef]
- Bernstein, E.R.; Posner, J.L.; Stoltenberg, D.E.; Hedtcke, J.L. Organically managed no-tillage rye-soybean systems: Agronomic, economic, and environmental assessment. Agron. J. 2011, 103, 1169–1179. [Google Scholar] [CrossRef]
- Smith, A.N.; Reberg-Horton, S.C.; Place, G.T.; Meijer, A.D.; Arellano, C.; Mueller, J.P. Rolled rye mulch for weed suppression in organic no-tillage soybeans. Weed Sci. 2021, 59, 224–231. [Google Scholar] [CrossRef]
- Nurse, R.E.; Mensah, R.; Robinson, D.W.; Leroux, G.D. Adzuki bean [Vigna angularis (Willd.) Ohwi & Ohashi], oilseed radish (Raphanus sativus L.), and cereal rye (Secale cereale L.) as living mulches with and without herbicides to control annual grasses in sweet corn (Zea mays L.). Can. J. Plant Sci. 2019, 99, 152–158. [Google Scholar]
- Uchino, H.; Iwama, K.; Jitsuyama, Y.; Yudate, T.; Nakamura, S. Yield losses of soybean and maize by competition with interseeded cover crops and weeds in organic-based cropping systems. Field Crops Res. 2009, 113, 342–351. [Google Scholar] [CrossRef] [Green Version]
- Vollmer, K.M.; Besançon, T.W.; Carr, B.L.; Van Gessel, M.J.; Scott, B.A. Spring-seeded cereal rye suppresses weeds in watermelon. Weed Technol. 2020, 34, 42–47. [Google Scholar] [CrossRef]
- Thelen, K.D.; Mutch, D.R.; Martin, T.E. Utility of interseeded winter cereal rye in organic soybean production systems. Agron. J. 2004, 96, 281–284. [Google Scholar] [CrossRef]
- Brainard, D.C.; Backer, J.; Noyes, D.C.; Myers, N. Rye living mulch effects on soil moisture and weeds in asparagus. HortScience 2012, 47, 58–63. [Google Scholar] [CrossRef] [Green Version]
- Liebman, M.; Mohler, C.L.; Staver, C.P. Ecological Management of Agricultural Weeds; Cambridge University Press: Cambridge, UK, 2001; p. 532. [Google Scholar]
- Leeson, J.Y.; Gailtier, J.; Grenkow, L. Manitoba Weed Survey of Annual Crops in 2016; Weed Survey Series Publ. 17-2; Agriculture and Agri-Food Canada: Saskatoon, SK, Canada, 2017; p. 203. [Google Scholar]
- Mason, H.E.; Spaner, D. Competitive ability of wheat in conventional and organic management systems: A review of the literature. Can. J. Plant Sci. 2006, 86, 333–343. [Google Scholar] [CrossRef]
- Littell, R.C.; Milken, G.A.; Stroup, W.W.; Wolfinger, R.D.; Schabenberger, O. SAS for Mixed Models, 2nd ed.; SAS Institute: Cary, NC, USA, 2006; p. 834. [Google Scholar]
- Lund, R.E. Tables for an approximate test for outliers in linear models. Technometrics 1975, 17, 473–476. [Google Scholar] [CrossRef]
- Luschei, E.C.; Jackson, R.D. Research methodologies and statistical approaches for multitactic systems. Weed Sci. 2005, 53, 393–403. [Google Scholar] [CrossRef]
- Onofri, A.; Carbonell, E.A.; Piepho, H.P.; Mortimer, A.M.; Cousens, R.D. Current statistical issues in Weed Research. Weed Res. 2010, 50, 5–24. [Google Scholar] [CrossRef]
- Saxton, A. A macro for converting mean separation output to letter groupings in proc mixed. In Proceedings of the 23rd SAS Users Group International, Nashville, TN, USA, 22–25 March 1998; SAS Institute: Cary, NC, USA, 1998. [Google Scholar]
- Environment Canada. Past Weather and Climate: Historical Data. Available online: www.climate.weather.gc.ca (accessed on 24 August 2021).
- Blackshaw, R.E.; Brandt, R.N.; Janzen, H.H.; Entz, T.; Grant, C.A.; Derksen, C.A. Differential response of weed species to added nitrogen. Weed Sci. 2003, 51, 532–539. [Google Scholar] [CrossRef]
- Wells, M.S.; Reberg-Horton, S.C.; Smith, A.N.; Grossman, J.M. The reduction of plant-available nitrogen by cover crop mulches and subsequent effects on soybean performance and weed interference. Agron. J. 2013, 105, 539–545. [Google Scholar] [CrossRef]
- Barnes, J.P.; Putnam, A.R. Role of benzoxazinones in allelopathy by rye (Secale cereale L.). J. Chem. Ecol. 1987, 13, 889–906. [Google Scholar] [CrossRef]
- Mirsky, S.B.; Ryan, M.R.; Teasdale, J.R.; Curran, W.S.; Reberg-Horton, C.S.; Spargo, J.T.; Wells, M.S.; Keene, C.L.; Moyer, J.W. Overcoming weed management challenges in cover crop-based organic rotational no-till soybean production in the eastern United States. Weed Technol. 2013, 27, 193–203. [Google Scholar] [CrossRef]
- Evans, R.; Lawley, Y.; Entz, M.H. Fall-seeded cereal cover crops differ in ability to facilitate low-till organic bean (Phaseolus vulgaris) production in a short-season growing environment. Field Crops Res. 2016, 191, 91–100. [Google Scholar] [CrossRef]
- Halde, C.; Gulden, R.H.; Entz, M.H. Selecting cover crop mulches for organic rotational no-till systems in Manitoba, Canada. Agron. J. 2014, 106, 1193–1204. [Google Scholar] [CrossRef]
- Martin, S.G.; Van Acker, R.C.; Friesen, L.F. Critical period of weed control in spring canola. Weed Sci. 2001, 49, 326–333. [Google Scholar] [CrossRef]
- Van Acker, R.C.; Swanton, C.J.; Weise, S.F. The critical period of weed control in soybean [Glycine max (L.) Merr.]. Weed Sci. 1993, 41, 194–200. [Google Scholar] [CrossRef]
- Knezevic, S.Z.; Datta, A. The critical period for weed control: Revisiting data analysis. Weed Sci. 2015, 63, 188–202. [Google Scholar] [CrossRef] [Green Version]
- Geddes, C.M. (Agriculture and Agri-Food Canada, Lethbridge, Alberta, Canada). Unpublished Observation. 2021. [Google Scholar]
- Knezevic, S.Z.; Evans, S.P.; Mainz, M. Row spacing influences the critical timing for weed removal in soybean (Glycine max). Weed Technol. 2003, 17, 666–673. [Google Scholar] [CrossRef]
- Rosset, J.D.; Gulden, R.H. Cultural weed management practices shorten the critical weed-free period for soybean grown in the Northern Great Plains. Weed Sci. 2020, 68, 79–91. [Google Scholar] [CrossRef]
- Bullied, W.J.; Marginet, A.M.; Van Acker, R.C. Conventional- and conservation-tillage systems influence emergence periodicity of annual weed species in canola. Weed Sci. 2003, 51, 886–897. [Google Scholar] [CrossRef]
- Maun, M.A.; Barrett, C.H. The biology of Canadian weeds. 77. Echinochloa crus-galli (L.) Beauv. Can. J. Plant Sci. 1986, 66, 739–759. [Google Scholar] [CrossRef]
- Manitoba Agricultural Services Corporation, Crop Dollar Values. 2021. Available online: https://www.masc.mb.ca/masc.nsf/crop_dollar_values.html (accessed on 15 October 2021).
- Mierau, A.; Johnson, E.N.; Gulden, R.H.; Weber, J.D.; May, W.E.; Willenborg, C.J. Minimizing competition between glyphosate-resistant volunteer canola (Brassica napus) and glyphosate-resistant soybean: Impact of soybean planting date and rate. Weed Technol. 2020, 34, 220–228. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spatial Arrangement | Herbicide Regime | Mulch Type b | Soybean Yield a (kg ha−1) | |||
|---|---|---|---|---|---|---|
| Carman 2013 | Carman 2014 | Melita 2014 | Combined | |||
| 38 cm | 2080 | 1010 A | 1320 | 1410 | ||
| 76 cm | 1800 | 650 B | 1130 | 1260 | ||
| Clodinafop | Unseeded | 1920 ab | 680 | 1170 | 1260 bc | |
| Clodinafop | Wheat (L) | 1490 b | 680 | 1160 | 1110 c | |
| Clodinafop | Rye (L) | 1480 b | 580 | 1050 | 1040 c | |
| Glyphosate | Unseeded | 2140 a | 920 | 1320 | 1460 ab | |
| Glyphosate | Wheat (T) | 2330 a | 1120 | 1510 | 1650 a | |
| Glyphosate | Rye (T) | 2270 a | 990 | 1120 | 1460 ab | |
| Spatial Arrangement | Herbicide Regime | Soybean Biomass a (kg ha−1) | |||
|---|---|---|---|---|---|
| Carman 2013 | Carman 2014 | Melita 2014 | Combined | ||
| 38 cm | - b | - | - | 3080 A | |
| 76 cm | - | - | - | 2530 B | |
| Clodinafop | 3490 b | 1200 | 2470 | 2380 b | |
| Glyphosate | 5450 a | 1690 | 2520 | 3220 a | |
| Herbicide Regime | Mulch Type | Volunteer Canola Plant Fecundity a (Seeds Plant−1) | |||
|---|---|---|---|---|---|
| Carman 2013 | Carman 2014 | Melita 2014 | Combined | ||
| Clodinafop | 270 B | 330 B | 550 | 380 B | |
| Glyphosate | 590 A | 870 A | 890 | 780 A | |
| Unseeded | - b | - | - | 700 a | |
| Wheat | - | - | - | 530 b | |
| Rye | - | - | - | 520 b | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Geddes, C.M.; Gulden, R.H. Wheat and Cereal Rye Inter-Row Living Mulches Interfere with Early Season Weeds in Soybean. Plants 2021, 10, 2276. https://doi.org/10.3390/plants10112276
Geddes CM, Gulden RH. Wheat and Cereal Rye Inter-Row Living Mulches Interfere with Early Season Weeds in Soybean. Plants. 2021; 10(11):2276. https://doi.org/10.3390/plants10112276
Chicago/Turabian StyleGeddes, Charles M., and Robert H. Gulden. 2021. "Wheat and Cereal Rye Inter-Row Living Mulches Interfere with Early Season Weeds in Soybean" Plants 10, no. 11: 2276. https://doi.org/10.3390/plants10112276
APA StyleGeddes, C. M., & Gulden, R. H. (2021). Wheat and Cereal Rye Inter-Row Living Mulches Interfere with Early Season Weeds in Soybean. Plants, 10(11), 2276. https://doi.org/10.3390/plants10112276