
First Detection and Molecular Characterization of Apple Stem Grooving Virus, Apple Chlorotic Leaf Spot Virus, and Apple Hammerhead Viroid in Loquat in Spain




Abstract
:1. Introduction
2. Results
2.1. Detection of Loquat Viruses by RT-PCR
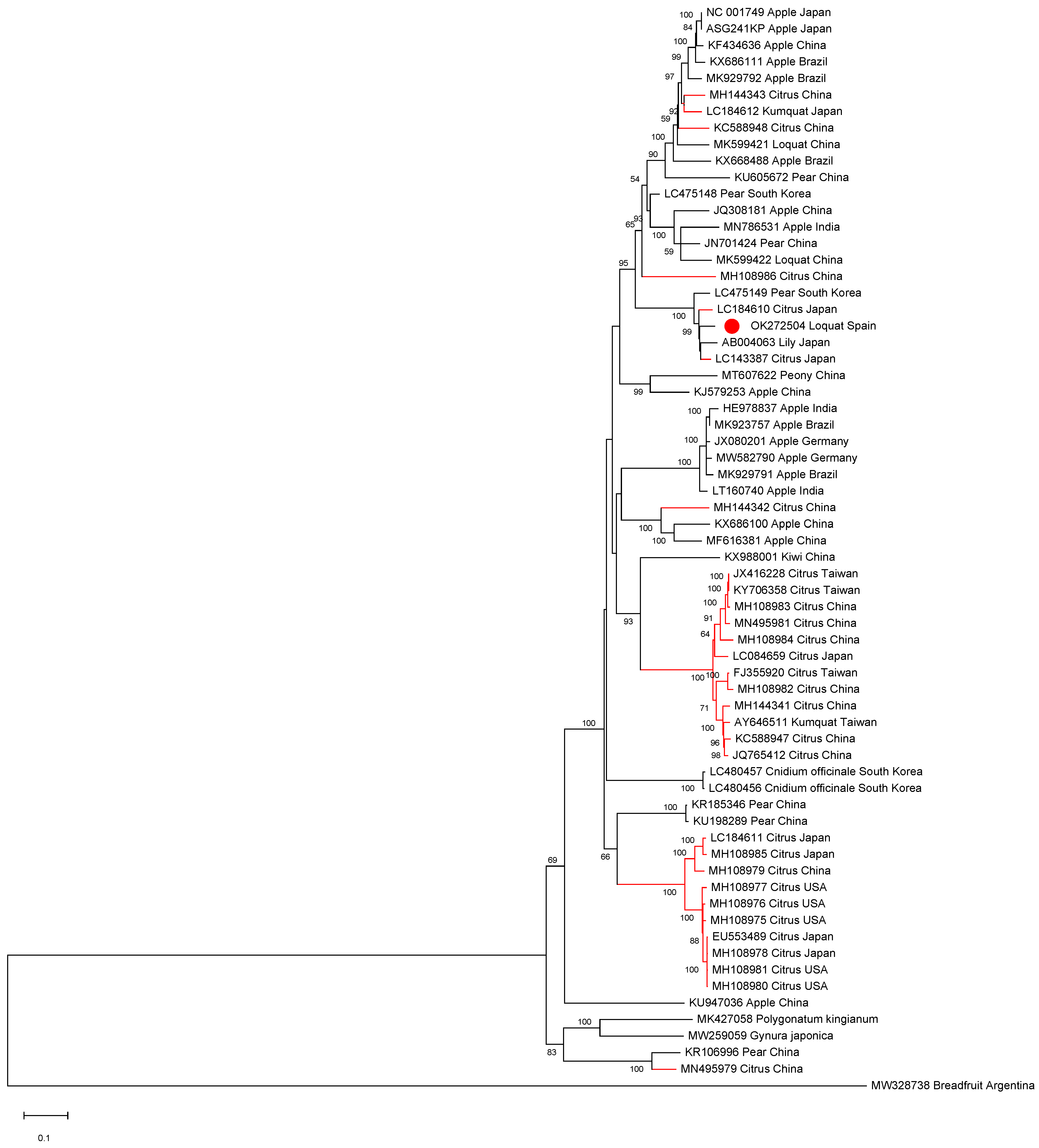
2.2. First Report of ASGV in Loquat in Spain
2.3. First Report of ACLSV in Loquat in Spain
2.4. Loquat Is a Natural Host of AHVd
3. Discussion
4. Materials and Methods
4.1. Plant Material and Sample Preparation
4.2. RNA Purification
4.3. ASGV, ACLSV, LoVA, and ASPV Detection by RT-PCR
4.4. HTS Analysis
4.5. AHVd Genome Amplification by RT-PCR
4.6. Sequence Alignment and Phylogenetic Analysis
4.7. AHVd Secondary Structure Prediction
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. The State of the World’s Biodiversity for Food and Agriculture; Bélanger, J., Pilling, D., Eds.; FAO Commission on Genetic Resources for Food and Agriculture Assessments: Rome, Italy, 2019; pp. 1–572. Available online: http://www.fao.org/3/CA3129EN/CA3129EN.pdf (accessed on 20 October 2021).
- Liu, Q.; Xuan, Z.; Wu, J.; Qiu, Y.; Li, M.; Zhang, S.; Wu, D.; Li, R.; Cao, M. Loquat is a new natural host of Apple stem grooving virus and Apple chlorotic leaf spot virus in China. Plant Dis. 2019, 103, 3290. [Google Scholar] [CrossRef]
- Morán, F.; Canales, C.; Olmos, A.; Ruiz-García, A.B. Loquat (Eriobotrya japonica) Is a New Natural Host of Apple Stem Pitting Virus. Plants 2020, 9, 1560. [Google Scholar] [CrossRef] [PubMed]
- Caballero, P.; Fernández, M.A. Loquat, production and market. Options Méditerranéennes: Série A. Séminaires Méditerranéens n. 58; First international symposium on loquat, Spain, 11-13/04/2002; Llácer, G., Badenes, M.L., Eds.; CIHEAM: Zaragoza, Spain, 2003; OM A58, 198. [Google Scholar]
- Jones, R.A.C.; Naidu, R.A. Global dimensions of plant virus diseases: Current status and future perspectives. Annu. Rev. Virol. 2019, 6, 387–409. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Yang, L.; Xuan, Z.; Wu, J.; Qiu, Y.; Zhang, S.; Wu, D.; Zhou, C.; Cao, M. Complete nucleotide sequence of loquat virus A, a member of the family Betaflexiviridae with a novel genome organization. Arch. Virol. 2020, 165, 223–226. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Plant Health (PLH). Pest categorisation of Tatter leaf virus. EFSA J. 2017, 15, 5033. [Google Scholar] [CrossRef]
- Wallace, J.M.; Drake, R.J. Tatter-leaf, a previously undescribed virus effect on citrus. Plant Dis. Rept. 1962, 46, 211–212. [Google Scholar]
- Lister, R.M.; Bancroft, J.B.; Nadakavukaren, M.J. Some sap-transmissible viruses from apple. Phytopathology 1965, 55, 859–870. [Google Scholar]
- De Sequeira, A.O. Studies on a virus causing stem grooving and graft union abnormalities in Virginia crab apple. Ann. Appl. Biol. 1967, 60, 59–66. [Google Scholar] [CrossRef]
- Waterworth, H.E.; Gilmer, R.M. Dark green epinasty of Chenopodium quinoa, a symdrome induced by a virus latent in apple and pear. Phytopathology 1969, 59, 334–338. [Google Scholar]
- Bhardwaj, P.; Hallan, V. Occurrence of Apple stem grooving virus on Rubus ellipticus, a perennial weed in India. Eur. J. Plant Pathol. 2019, 153, 311–319. [Google Scholar] [CrossRef]
- Bhardwaj, P.; Hallan, V. Molecular evidence of Apple stem grooving virus infecting Ficus palmata. Trees 2019, 33, 1–9. [Google Scholar] [CrossRef]
- Yoshikawa, N.; Sasaki, E.; Kato, M.; Takahashi, T. The nucleotide sequence of apple stem grooving virus capillovirus genome. Virology 1992, 191, 98–105. [Google Scholar] [CrossRef]
- Massart, S.; Jijakli, M.H.; Kummert, J. Apple stem grooving virus. In Virus and Virus-Like Diseases of Pome and Stone Fruits; Hadidi, A., Barba, M., Candresse, T., Jelkmann, W., Eds.; The American Phytopathological Society: Saint Paul, MN, USA, 2011; pp. 29–33. [Google Scholar]
- Tan, S.-h.; Osman, F.; Bodaghi, S.; Dang, T.; Greer, G.; Huang, A.; Hammado, S.; Shurooq, A.-H.; Campos, R.; Vidalakis, G. Full genome characterization of 12 citrus tatter leaf virus isolates for the development of a detection assay. PLoS ONE 2019, 14, e0223958. [Google Scholar] [CrossRef] [PubMed]
- Martelli, G.P.; Candresse, T.; Namba, S. Trichoviruses. A new genus of plant viruses. Arch. Virol. 1994, 134, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Al Rwahnih, M.; Turturo, C.; Minafra, A.; Salsarelli, P.; Myrta, A.; Pallás, V.; Savino, V. Molecular variability of Apple chlorotic leaf spot virus in different hosts and geographical regions. J. Plant Pathol. 2004, 86, 117–122. [Google Scholar]
- Yaegashi, H.; Yoshikawa, N.; Candresse, T. Apple chlorotic leaf spot virus in pome fruits. In Virus and Virus-Like Diseases of Pome and Stone Fruits; Hadidi, A., Barba, M., Candresse, T., Jelkmann, W., Eds.; The American Phytopathological Society: Saint Paul, MN, USA, 2011; pp. 17–21. [Google Scholar]
- Mathioudakis, M.M.; Candresse, T.; Katis, N.I. First report of apple chlorotic leaf spot virus in quince in Greece. Plant Dis. 2007, 91, 462. [Google Scholar] [CrossRef] [PubMed]
- Katsiani, A.T.; Maliogka, V.I.; Candresse, T.; Katis, N.I. Host-range studies, genetic diversity and evolutionary relationships of ACLSV isolates from ornamental, wild and cultivated Rosaceous species. Plant Pathol. 2014, 63, 63–71. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Qi, S.; Tang, N.; Zhang, X.; Chen, S.; Zhu, P.; Ma, L.; Cheng, J.; Xu, Y.; Lu, M.; et al. Discovery of replicating circular RNAs by RNA-seq and computational algorithms. PLoS Pathog. 2014, 10, e1004553. [Google Scholar] [CrossRef] [PubMed]
- Serra, P.; Messmer, A.; Sanderson, D.; James, D.; Flores, R. Apple hammerhead viroid-like is a bona fide viroid: Autonomous replication and structural features support its inclusion as a new member in the genus Pelamoviroid. Virus Res. 2018, 249, 8–15. [Google Scholar] [CrossRef] [Green Version]
- Messmer, A.; Sanderson, D.; Braun, G.; Serra, P.; Flores, R.; James, D. Molecular and phylogenetic identification of unique isolates of hammerhead viroid-like RNA from ‘Pacific Gala’ apple (Malus domestica) in Canada. Can. J. Plant Pathol. 2017, 39, 342–353. [Google Scholar] [CrossRef]
- Szostek, S.A.; Wright, A.; Harper, S. First report of apple hammerhead viroid in the US, Japan, Italy, Spain, and New Zealand. Plant Dis. 2018, 102, 2670. [Google Scholar] [CrossRef]
- Chiumenti, M.; Navarro, B.; Venerito, P.; Civita, F.; Minafra, A.; Di Serio, F. Molecular variability of apple hammerhead viroid from Italian apple varieties supports the relevance in vivo of its branched conformation stabilized by a kissing loop interaction. Virus Res. 2019, 270, 197644. [Google Scholar] [CrossRef]
- Malandraki, I.; Beris, D.; Isaioglou, I.; Olmos, A.; Varveri, C.; Vassilakos, N. Simultaneous detection of three pome fruit tree viruses by one-step multiplex quantitative RT-PCR. PLoS ONE 2017, 12, e0180877. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Yang, Z.; Hong, N.; Wang, G.; Ning, G.; Xu, W. Deep sequencing reveals a novel closterovirus associated with wild rose leaf rosette disease. Mol. Plant Pathol. 2014, 16, 449–458. [Google Scholar] [CrossRef]
- Salmon, M.; Vendrame, M.; Kummert, J.; Lepoivre, P. Detection of apple chlorotic leaf spot virus using a 5′ nuclease assay with a fluorescent 3′ minor groove binder-DNA probe. J. Virol. Methods 2002, 104, 99–106. [Google Scholar] [CrossRef]
- Alas, T.; Baloglu, S.; Caglar, B.K.; Gunes, A. Detection and characterization of citrus tatter leaf virus (CTLV) and citrus yellow vein clearing virus (CYVCV) in citrus trees from Cyprus. Saudi J. Biol. Sci. 2019, 26, 995–998. [Google Scholar] [CrossRef]
- Stecher, G.; Tamura, K.; Kumar, S. Molecular evolutionary genetics analysis (MEGA) for macOS. Mol. Biol. Evol. 2020, 37, 1237–1239. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: Oxford, UK, 2000. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Variety | Symptomatology | ASGV | ACLSV | LoVA | ASPV |
|---|---|---|---|---|---|---|
| SL73.1 | Andrés | Symptomless | − | + | − | − |
| SL73.2 | Algerie | Symptomless | + | − | − | − |
| SL73.4 | Xirlero | Symptomless | − | + | − | − |
| SL73.6 | Xirlero | Chlorotic mottling | − | + | − | + |
| SL73.11 | Juliana | Symptomless | − | + | − | − |
| SL73.12 | Algerie | Symptomless | − | + | − | − |
| SL73.16 | Algerie | Symptomless | − | + | − | − |
| SL73.18 | Algerie | Symptomless | − | + | − | − |
| SL73.20 | Algerie | Symptomless | − | + | − | − |
| SL73.32 | Xirlero | Symptomless | + | − | − | − |
| SL73.33 | Xirlero | Chlorotic mottling | − | + | − | − |
| SL73.40 | Crisanto Amadeo | Symptomless | − | + | − | − |
| SL73.41 | Crisanto Amadeo | Symptomless | − | + | − | − |
| SL73.42 | Xirlero | Chlorotic mottling | − | + | − | − |
| SL73.43 | Xirlero | Chlorotic mottling | − | + | − | − |
| SL73.54 | Xirlero | Symptomless | + | + | − | + |
| SL73.55 | Xirlero | Symptomless | − | + | − | − |
| SL73.56 | Xirlero | Symptomless | − | + | − | − |
| SL73.57 | Algerie | Symptomless | − | + | − | − |
| SL73.61 | Algerie | Symptomless | − | + | − | − |
| SL73.69 | Xirlero | Chlorotic mottling | − | + | − | − |
| SL73.71 | Xirlero | Symptomless | − | + | − | − |
| SL73.72 | Toni tomaca | Symptomless | − | + | − | − |
| SL73.76 | Algerie | Symptomless | − | + | − | + |
| SL73.78 | Algerie | Symptomless | − | + | − | + |
| SL73.79 | Algerie | Symptomless | − | + | − | + |
| SL73.82 | Toni tomaca | Symptomless | − | + | − | − |
| SL73.83 | Toni tomaca | Symptomless | + | − | − | + |
| SL73.84 | Toni tomaca | Symptomless | + | + | − | + |
| SL73.87 | Toni tomaca | Symptomless | + | + | − | − |
| SL73.89 | Xirlero | Symptomless | − | + | − | + |
| Virus | Primer | Sequence (5′–3′) | Size (bp) | Reference |
|---|---|---|---|---|
| ASGV | qASG-F | GGGACTTGCCACYAACATTT | 72 | [27] |
| qASG-R | CACCCAAGGGCTYTTTTCAA | |||
| ASG-P | AGAAATGGCCCAAAGC | |||
| ASGV-499-1 | CCCGCTGTTGGATTTGATACACCTC | 499 | [28] | |
| ASGV-499-2 | GGAATTTCACACGACTCCTAACCCTCC | |||
| ACLSV | ACLSV 5F | GCCTACAAATTAGGTGAGAGGCTC | 288 | [29] |
| ACLSV 8R | TTCCAATGGATCATGAGGTC | |||
| MGB26 | FAM-ATTCACATCTTGAAATT-MGB | |||
| LoVA | LoVA-F | TAATGAAGTCCAAGGAAGCACC | 453 | [6] |
| LoVA-R | GGTCTCATTTCTGAAACCTCGA | |||
| ASPV | qASP-FS | CGCTTTCTACGCGAAGCATGT | 385 | [3] |
| ASPV-RS | TTGGGATCAACTTTATTAAAAGCATAA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Canales, C.; Morán, F.; Olmos, A.; Ruiz-García, A.B. First Detection and Molecular Characterization of Apple Stem Grooving Virus, Apple Chlorotic Leaf Spot Virus, and Apple Hammerhead Viroid in Loquat in Spain. Plants 2021, 10, 2293. https://doi.org/10.3390/plants10112293
Canales C, Morán F, Olmos A, Ruiz-García AB. First Detection and Molecular Characterization of Apple Stem Grooving Virus, Apple Chlorotic Leaf Spot Virus, and Apple Hammerhead Viroid in Loquat in Spain. Plants. 2021; 10(11):2293. https://doi.org/10.3390/plants10112293
Chicago/Turabian StyleCanales, Celia, Félix Morán, Antonio Olmos, and Ana Belén Ruiz-García. 2021. "First Detection and Molecular Characterization of Apple Stem Grooving Virus, Apple Chlorotic Leaf Spot Virus, and Apple Hammerhead Viroid in Loquat in Spain" Plants 10, no. 11: 2293. https://doi.org/10.3390/plants10112293
APA StyleCanales, C., Morán, F., Olmos, A., & Ruiz-García, A. B. (2021). First Detection and Molecular Characterization of Apple Stem Grooving Virus, Apple Chlorotic Leaf Spot Virus, and Apple Hammerhead Viroid in Loquat in Spain. Plants, 10(11), 2293. https://doi.org/10.3390/plants10112293