
Natural Biostimulant Attenuates Salinity Stress Effects in Chili Pepper by Remodeling Antioxidant, Ion, and Phytohormone Balances, and Augments Gene Expression


 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Trial Location and Climatic Conditions
2.2. Plant Material, Experimental Layout, and Cultural Practices
2.3. Preparation and Analysis of Bee-Honey (HB)
2.4. Evaluation of Plant Parameters for Growth and Yield
2.5. Evaluation of Photosynthetic Efficiency
2.6. Leafy Tissue Integrity Assessment
2.7. Evaluation of Non-Enzymatic Antioxidant Contents
2.8. Evaluation of Antioxidant Enzymes
2.9. Extraction and Evaluation of Phytohormone Levels
2.10. Molecular Analysis and Gene Expression
2.10.1. Isolation of Total RNA and Synthesis of cDNA
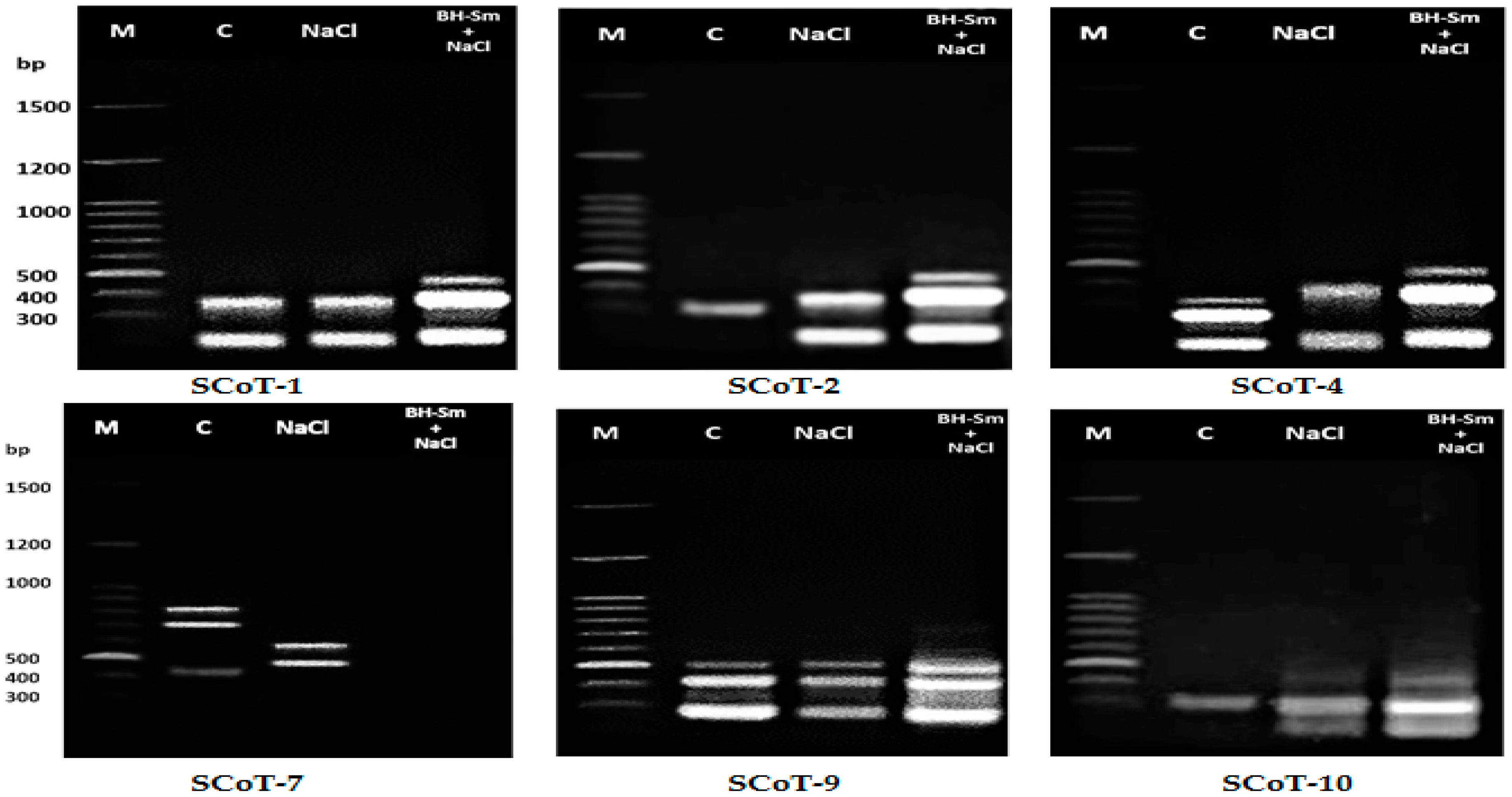
2.10.2. cDNA-SCoT Amplification and Detection
2.10.3. Patterns of Banding Related to the Expression of cDNA-SCoT
2.11. Data Analysis
3. Results
3.1. Growth and Fruit Yield Parameters
3.2. Photosynthetic Efficiency
3.3. Leaf Relative Water Content (RWC) and Osmoprotectant Contents
3.4. K+, Na+, and Cl− Ions Contents, and the Ratio of K+/Na+
3.5. Oxidative Stress Marker Levels and Membrane Damages Related to Oxidative Stress
3.6. Non-Enzymatic Antioxidant Contents
3.7. Enzymatic Antioxidant Activities
3.8. Hormonal Content
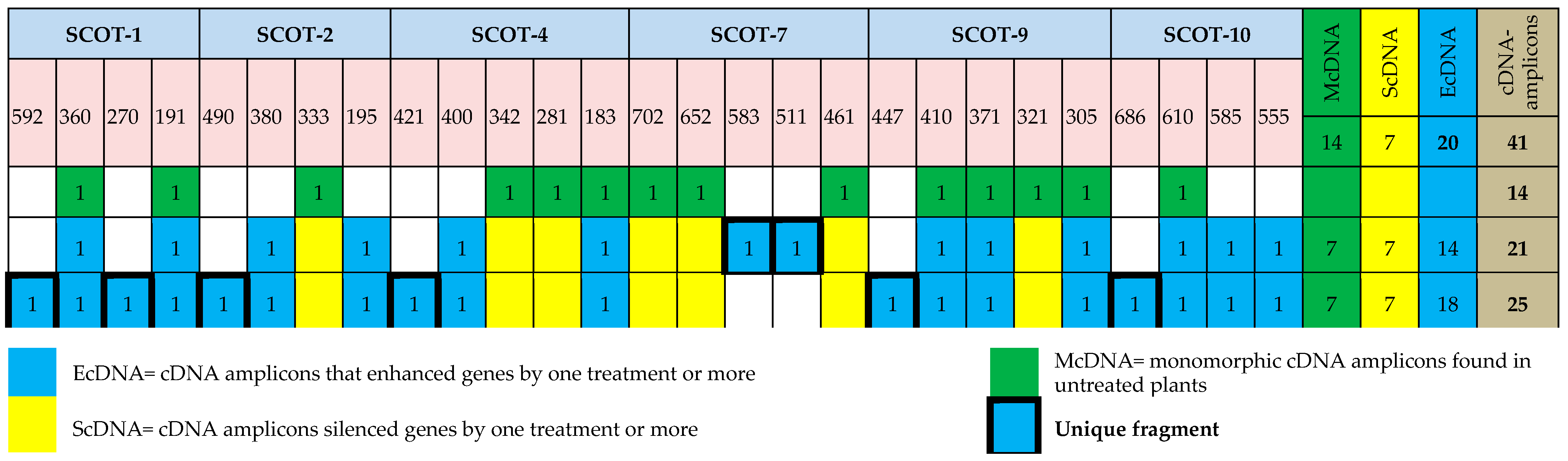
3.9. Differential Expression Analysis Using the cDNA-SCoT Technique
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mani, F. Effect of Water Stress on Some Biochemical and Physiological Characters of Chili Pepper (Capsicum frutescens L.). JECET 2015, 4, 75–83. [Google Scholar]
- Howard, L.R.; Talcott, S.T.; Brenes, C.H.; Villalon, B. Changes in phytochemical and antioxidant activity of selected pepper cultivars (Capsicum species) as influenced by maturity. J. Agric. Food Chem. 2000, 48, 1713–1720. [Google Scholar] [CrossRef] [PubMed]
- Kpinkoun, J.K.; Amoussa, A.M.; Mensah, A.C.G.; Komlan, F.A.; Kinsou, E.; Lagnika, L.; Gandonou, C.B. Effect of salt stress on flowering, fructification, and fruit nutrients concentration in a local cultivar of chili pepper (Capsicum frutescens L.). Int. J. Plant Physiol. Biochem. 2019, 11, 1–7. [Google Scholar]
- Al-Elwany, O.A.; Mohamed, G.F.; Abdehrahman, H.A.; Rady, M.M.; Abdel Latef, A.H. Exogenous glutathione-mediated tolerance to deficit irrigation stress in salt-affected Capsicum frutescence (L.) plants is connected with higher antioxidant content and proper ion homeostasis. Not. Bot. Horti Agrobot. Cluj-Napoca 2020, 48, 1957–1979. [Google Scholar] [CrossRef]
- Abdel Latef, A.A.; Chaoxing, H. Does inoculation with Glomus mosseae improve salt tolerance in pepper plants? J. Plant Growth Regul. 2014, 33, 64–653. [Google Scholar] [CrossRef]
- ElSayed, A.I.; Boulila, M.; Rafudeen, M.S.; Mohamed, A.H.; Sengupta, S.; Rady, M.; Omar, A.A. Melatonin Regulatory Mechanisms and Phylogenetic Analyses of Melatonin Biosynthesis Related Genes Extracted from Peanut under Salinity Stress. Plants 2020, 9, 854. [Google Scholar] [CrossRef] [PubMed]
- Rady, M.M.; Desoky, E.-S.M.; Ahmed, S.M.; Majrashi, A.; Ali, E.F.; Arnaout, S.M.A.I.; Selem, E. Foliar Nourishment with Nano-Selenium Dioxide Promotes Physiology, Biochemistry, Antioxidant Defenses, and Salt Tolerance in Phaseolus vulgaris. Plants 2021, 10, 1189. [Google Scholar] [CrossRef] [PubMed]
- Alam, H.; Khattak, J.Z.K.; Ksiksi, T.S.; Saleem, M.H.; Fahad, S.; Sohail, H.; Ali, Q.; Zamin, M.; El-Esawi, M.A.; Saud, S.; et al. Negative impact of long-term exposure of salinity and drought stress on native Tetraena mandavillei L. Physiol. Plant. 2021, 172, 1336–1351. [Google Scholar] [CrossRef] [PubMed]
- Mumtaz, S.; Saleem, M.H.; Hameed, M.; Batool, F.; Parveen, A.; Amjad, S.F.; Mahmood, A.; Arfan, M.; Ahmed, S.; Yasmin, H.; et al. Anatomical adaptations and ionic homeostasis in aquatic halophyteCyperus laevigatusL. under high salinities. Saudi J. Biol. Sci. 2021, 28, 2655–2666. [Google Scholar] [CrossRef]
- Hassan, A.; Amjad, S.F.; Saleem, M.H.; Yasmin, H.; Imran, M.; Riaz, M.; Ali, Q.; Joyia, F.A.; Mobeen, A.S.; Ali, S.; et al. Foliar application of ascorbic acid enhances salinity stress tolerance in barley (Hordeum vulgare L.) through modulation of morpho-physio-biochemical attributes, ions uptake, osmoprotectants, and stress response genes expression. Saudi J. Biol. Sci. 2021, 28, 4276–4290. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afzal, J.; Saleem, M.H.; Batool, F.; Elyamine, A.M.; Rana, M.S.; Shaheen, A.; El-Esawi, M.A.; Javed, M.T.; Ali, Q.; Ashraf, M.A.; et al. Role of Ferrous Sulfate (FeSO4) in Resistance to Cadmium Stress in Two Rice (Oryza sativa L.) Genotypes. Biomolecules 2020, 10, 1693. [Google Scholar] [CrossRef]
- De Pinto, M.C.; Locato, V.; De Gara, L. Redox regulation in plant programmed cell death. Plant. Cell Environ. 2012, 35, 234–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, A.; Saleem, M.H.; Parveen, A.; Mobeen, A.S.; Shakoor, A.; Ali, Q.; Chattha, M.S.; El-Sheikh, M.A.; Ali, S. Oxidative stress alleviation as indicated by enzymatic and nonenzymatic antioxidants and osmoregulators in barley (Hordeum vulgare l.) Under salt (NaCl) stress by ascorbic acid (ASA). Pak. J. Bot. 2020, 54, 1. [Google Scholar] [CrossRef]
- Yasmin, H.; Bano, A.; Wilson, N.L.; Nosheen, A.; Naz, R.; Hassan, M.N.; Ilyas, N.; Saleem, M.H.; Noureldeen, A.; Ahmad, P.; et al. Drought-tolerant Pseudomonas sp. showed differential expression of stress-responsive genes and induced drought tolerance in Arabidopsis thaliana. Physiol. Plant. 2021, 1–15. [Google Scholar] [CrossRef]
- De Azevedo, A.D.N.; Prisco, J.T.; Enéas-Filho, J.; de Abreu, C.E.B.; Gomes-Filho, E. Effect of salt stress on antioxidative enzymes and lipid peroxidation in leaves and roots of salt-tolerant and salt-sensitive maize genotypes. Environ. Exp. Bot. 2006, 56, 87–94. [Google Scholar] [CrossRef]
- Abdel Latef, A.A.; Mostofa, M.G.; Rahman, M.M.; Abdel-Farid, I.B.; Tran, L.S.P. Extracts from yeast and carrot roots enhance maize Performance under seawater-induced salt stress by altering physio-biochemical characteristics of stressed plants. J. Plant Growth Regul. 2019, 38, 966–979. [Google Scholar] [CrossRef]
- Alharby, H.F.; Al-Zahrani, H.S.; Hakeem, K.R.; Alsamadany, H.; Desoky, E.-S.M.; Rady, M.M. Silymarin-Enriched Biostimulant Foliar Application Minimizes the Toxicity of Cadmium in Maize by Suppressing Oxidative Stress and Elevating Antioxidant Gene Expression. Biomolecules 2021, 11, 465. [Google Scholar] [CrossRef] [PubMed]
- Desoky, E.-S.M.; Elrys, A.S.; Mansour, E.; Eid, R.S.M.; Selem, E.; Rady, M.M.; Ali, E.F.; Mersal, G.A.M.; Semida, W.M. Application of biostimulants promotes growth and productivity by fortifying the antioxidant machinery and suppressing oxidative stress in faba bean under various abiotic stresses. Sci. Hortic. 2021, 288, 110340. [Google Scholar] [CrossRef]
- Zaki, S.S.; Rady, M.M. Moringa oleifera leaf extract improves growth, physiochemical attributes, antioxidant defence system and yields of salt-stressed Phaseolus vulgaris L. plants. Int. J. ChemTech Res. 2015, 8, 120–134. [Google Scholar]
- Rady, M.M.; Abd El-Mageed, T.A.; Abdurrahman, H.A.; Mahdi, A.H. Humic acid application improves field Performance of cotton (Gossypium barbadense L.) under saline conditions. J. Anim. Plant Sci. 2016, 26, 487–493. [Google Scholar]
- Semida, W.M.; Abd El-Mageed, T.A.; Hemida, K.A.; Rady, M.M. Natural bee-honey based biostimulants confer salt tolerance in onion via modulation of the antioxidant defence system. J. Hortic. Sci. Biotechnol. 2019, 94, 632–642. [Google Scholar] [CrossRef]
- Rady, M.M.; Bhavya, V.C.; Howladar, S.M. Common bean (Phaseolus vulgaris L.) seedlings overcome NaCl stress as a result of presoaking in Moringa oleifera leaf extract. Sci. Hortic. 2013, 162, 63–70. [Google Scholar] [CrossRef]
- Ertani, A.; Nardi, S.; Altissimo, A. Long-term research activity on the biostimulant properties of natural origin compounds—A review. Acta Hortic. 2013, 1009, 181–188. [Google Scholar] [CrossRef]
- White, J.W. Physical Characteristics of Honey. In Honey: A Comprehensive Survey; Carne, E., Ed.; Heinemann: London, UK, 1975; pp. 207–239. [Google Scholar]
- Afshar, R.K.; Chaichi, M.R.; Jovini, M.A.; Jahanzad, E.; Hashemi, M. Accumulation of silymarin in milk thistle seeds under drought stress. Planta 2015, 242, 539–543. [Google Scholar] [CrossRef]
- Abo-Doma, A.; Azzam, C.R. Hunting of some differentially expressed genes under salt stress in wheat. Egypt. J. Plant Breed 2007, 11, 233–244. [Google Scholar]
- Azzam, C.R.; Edris, S.; Mansour, A.A. Changes in wheat P5CS gene expression in response to salt stress in wheat. Egypt. J. of Genet. Cytol. 2009, 38, 375–386. [Google Scholar]
- Azzam, C.R.; Abd-Elnaby, Z.M.; Salem, A.K. Influence of Agro-Ecological Conditions on Gene Expression, Yield and Yield Components of the Mono-Cut (Fahl) Type of Berseem. Egypt. J. Plant Breed 2012, 16, 135–159. [Google Scholar] [CrossRef]
- Ismael, R.R.; Ahmad, E.S.; Abdel-Fattah, M.; Omar, M.N.A.; Azzam, C.R.; Zahran, H.H. Effect of plant growth-promoting rhizobacteria (PGPR) on growth and symbiotic nitrogen fixation of Vicia faba plants under salt stress. Plant Sci. J. 2018, 7, 1–19. [Google Scholar]
- Azzam, C.R.; Abd-Elnaby, Z.M.; Mohamed, N.A. Salt Tolerance Associated with Molecular Markers in Alfalfa. J. Biosci. Appl. Res. 2019, 5, 416–428. [Google Scholar] [CrossRef]
- Khaled, K.A.; El-Demardash, I.S.; Amer, E.A.M. Genetic Polymorphism among Some Sugarcane Germplasm Collections as revealed by RAPD and ISSR analyses. Life Sci. J. 2015, 12, 159–167. [Google Scholar]
- AL-Taweel, S.K.; Abdel-Aziz, R.M.; Rabea, K.; Khaled, K. Studying cDNA SCoT in Response to Salinity Stress in Stevia Rebaudiana Bertoni. SABRAO J. Breed. Genet. 2019, 51, 281–294. [Google Scholar]
- Khaled, K.A.; El-Arabi, N.I.; Sabry, N.M.; El-Sherbiny, S. Sugarcane Genotypes Assessment under Drought Condition Using Amplified Fragment Length Polymorphism. Biotechnology 2018, 17, 120–127. [Google Scholar] [CrossRef] [Green Version]
- Luo, C.; He, X.H.; Hu, Y.; Yu, H.X.; Ou, S.J.; Fang, Z.B. Oligo-dT anchored cDNA–SCoT: A novel differential display method for analyzing differential gene expression in response to several stress treatments in mango (Mangifera indica L.). Gene 2014, 548, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.H.; Zhang, B.Q.; Zeng, X.P.; Chen, H.; Yang, L.T.; LI, Y.-R.; Chen, B.S. cDNA- SCoT analysis of differentially expressed genes in sugarcane induced by Leifsonia xyli subsp. Xyli. Acta Agron. Sin. 2013, 39, 1119–1126. [Google Scholar] [CrossRef]
- Al-Janabi, A.S.; AL-Rawi, T.K. Effect of irrigation times and termination on gene expression of OeDGAT1, OeFAD2.1, and OeFAD6 and oil quality and quantity in olive cultivars. SABRAO J. Breed. Genet. 2018, 50, 329–343. [Google Scholar] [CrossRef]
- Rady, M.M.; Rehman, H. Supplementing organic biostimulants into growing media enhances growth and nutrient uptake of tomato transplants. Sci. Hortic. 2016, 203, 192–198. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 17th ed.; The Association of Official Analytical Chemists: Gaithersburg, MD, USA, 2000; Methods 925.10, 65.17, 974.24, 992.16. [Google Scholar]
- Bogdanov, S.; Baumann, E. Determination of honey sugars with HPLC. Mitt. Gebiete. Lebensm. Hyg. 1988, 79, 198–206. [Google Scholar]
- Chapman, H.D.; Pratt, P.F. Methods of Analysis for Soil, Plants, and Water; The University of California, Division of Agricultural Science: Berkeley, CA, USA, 1961. [Google Scholar]
- Mukherjee, S.P.; Choudhuri, M.A. Implications of water stress-induced changes in the levels of endogenous ascorbic acid and hydrogen peroxide in Vigna seedlings. Physiol. Plant. 1983, 58, 166–170. [Google Scholar] [CrossRef]
- Lee, S.C.; Kim, J.H.; Jeong, S.M.; Kim, D.R.; Ha, J.U.; Nam, K.C. Effect of far-infrared radiation on the antioxidant activity of rice hulls. J. Agric. Food Chem. 2003, 51, 4400–4403. [Google Scholar] [CrossRef] [PubMed]
- Konrad, M.L.F.; Silva, J.A.B.; Furlani, P.R.; Machado, E.C. Trocas gasosas e fluorescência da clorofila em seis cultivares de cafeeiro sob estresse de alumínio. Bragantia 2005, 64, 30–37. [Google Scholar] [CrossRef]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolutions. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Avron, M. Photophosphorylation by swiss-chard chloroplasts. Biochim. Biophys. Acta 1960, 40, 257–272. [Google Scholar] [CrossRef]
- Nagy, Z.; Daood, H.; Koncsek, A.; Molnár, H.; Helyes, L. The simultaneous determination of capsaicinoids, tocopherols, and carotenoids in pungent pepper powder. J. Liq. Chromatogr. Relat. Technol. 2017, 40, 199–209. [Google Scholar] [CrossRef]
- Osman, A.S.; Rady, M.M. Effect of humic acid as an additive to growing media to enhance the production of eggplant and tomato transplants. J. Hortic. Sci. Biotechnol. 2014, 89, 237–244. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldeen, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Irigoyen, J.J.; Emerich, D.W.; Sanchez-Diaz, M. Water stress-induced changes in the concentrations of proline and total soluble sugars in nodulated alfalfa (Medicago sativa) plants. Plant Physiol. 1992, 8, 455–460. [Google Scholar] [CrossRef]
- Bessieres, M.A.; Gibon, Y.; Lefeuvre, J.C.; Larher, F. A single-step purification for glycine betaine determination in plant extracts by isocratic HPLC. J. Agric. Food Chem. 1999, 47, 3718–3722. [Google Scholar] [CrossRef] [PubMed]
- Subbarao, G.V.; Wheeler, R.M.; Stutte, G.W.; Levine, L.H. How far can sodium substitute for potassium in red beet? J. Plant Nutr. 1999, 22, 1745–1761. [Google Scholar] [CrossRef] [PubMed]
- Wilde, S.A.; Corey, R.B.; Lyer, J.G.; Voigt, G.K. Soil and Plant Analysis for Tree Culture, 3rd ed.; Oxford and IBM Publishers: New Delhi, India, 1985; pp. 93–106. [Google Scholar]
- Higinbotham, N.; Bud, E.; Foster, R.J. Mineral ion contents and cell trans-membranes electro-potentials of peas and oat seedling tissues. Plant Physiol. 1967, 24, 37–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubis, J. Exogenous spermidine differentially alters activities of some scavenging system enzymes, H2O2, and superoxide radical levels in water-stressed cucumber leaves. J. Plant Physiol. 2008, 165, 397–406. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation isolated chloroplasts: Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Rady, M.M. Effect of 24-epibrassinolide on growth, yield, antioxidant system, and cadmium content of bean (Phaseolus vulgaris L.) plants under salinity and cadmium stress. Sci. Hortic. 2011, 129, 232–237. [Google Scholar] [CrossRef]
- Kampfenkel, K.; Van Montagu, M. Extraction and determination of ascorbate and dehydroascorbate from plant tissue. Anal. Biochem. 1995, 225, 165–167. [Google Scholar] [CrossRef]
- Griffth, O.W. Determination of glutathione and glutathione disulfide using glutathione reductase and 2 vinyl pyridine. Anal. Biochem. 1980, 106, 207–212. [Google Scholar] [CrossRef]
- Konings, E.J.; Roomans, H.H.; Beljaars, P.R. Liquid chromatographic determination of tocopherols and tocotrienols in margarine, infant foods, and vegetables. J. AOAC Int. 1996, 79, 902–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ching, L.S.; Mohamed, S. Alpha-tocopherol content in 62 edible tropical plants. J. Agric. Food Chem. 2001, 49, 3101–3105. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, K.; Wu, X.; Liu, R.H. Antioxidant activity of apple peels. J. Agric. Food Chem. 2003, 51, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Ali, Q.; Ali, S.; Iqbal, N.; Javed, M.T.; Rizwan, M.; Khaliq, R.; Shahid, S.; Perveen, R.; Alamri, S.A.; Alyemeni, M.N.; et al. Alpha-tocopherol fertigation confers growth physio-biochemical and qualitative yield enhancement in field-grown water deficit wheat (Triticum aestivum L.). Sci. Rep. 2019, 9, 12924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arampatzis, D.A.; Karkanis, A.C.; Tsiropoulos, N.G. Impact of Plant Density and Mepiquat Chloride on Growth, Yield, and Silymarin Content of Silybum marianum Grown under Mediterranean Semi-Arid Conditions. Agronomy 2019, 9, 669. [Google Scholar] [CrossRef] [Green Version]
- Arampatzis, D.A.; Karkanis, A.C.; Tsiropoulos, N.G. Silymarin content and antioxidant activity of seeds of wild Silybum marianum populations growing in Greece. Ann. Appl. Biol. 2019, 174, 61–73. [Google Scholar] [CrossRef] [Green Version]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Chance, B.; Maehly, A.C. Assay of catalases and peroxidases. Methods Enzymol. 1955, 2, 764–775. [Google Scholar]
- Chandlee, J.M.; Scandalios, J.G. Analysis of variants affecting the catalase developmental program in maize scutellum. Theor. Appl. Genet. 1984, 69, 71–77. [Google Scholar] [CrossRef]
- Asada, K. Production and Scavenging of Reactive Oxygen Species in Chloroplasts and Their Functions. Plant Physiol. 2006, 141, 391–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Terra, L.A.; Soares, C.P.; Meneses, C.H.S.G.; Sfeir, T.Z.M.; Souza, E.M.; Silveira, V.; Vidal, S.M.; Baldani, J.I.; Schwab, S. Transcriptome and proteome profiles of the diazotroph Nitrospirillum amazonense strain CBAmC in response to the sugarcane apoplast fluid. Plant Soil 2020, 451, 145–168. [Google Scholar] [CrossRef]
- Novák, O.; Hauserová, E.; Amakorová, P.; Doležal, K.; Strnad, M. Cytokinin profiling in plant tissues using ultra-Performance liquid chromatography-electrospray tandem mass spectrometry. Phytochemistry 2008, 69, 2214–2224. [Google Scholar] [CrossRef] [PubMed]
- Siegrist, J.; Orober, M.; Buchenauer, H. β-Aminobutyric acid-mediated enhancement of resistance in tobacco to tobacco mosaic virus depends on the accumulation of salicylic acid. Physiol. Mol. Plant Pathol. 2000, 56, 95–106. [Google Scholar] [CrossRef] [Green Version]
- Metwally, A.; Finkemeier, I.; Georgi, M.; Dietz, K.J. Salicylic acid alleviates the cadmium toxicity in barley seedlings. Plant Physiol. 2003, 132, 272–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeevaart, J.A. Changes in the levels of abscisic acid and its metabolites in excised leaf blades of Xanthium strumarium during and after water stress. Plant Physiol. 1980, 66, 672–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adhikari, S.; Saha, S.; Bandyopadhyay, T.K.; Ghosh, P. Efficiency of ISSR marker for characterization of Cymbopogon germplasm and their suitability in molecular barcoding. Plant Syst. Eval. 2015, 301, 439–450. [Google Scholar] [CrossRef]
- Gorji, A.M.; Poczai, P.; Polgar, Z.; Taller, J. Efficiency of arbitrarily amplified dominant markers (SCOT, ISSR, and RAPD) for diagnostic fingerprinting in tetraploid potato. Am. J. Pot. Res. 2011, 88, 226–237. [Google Scholar] [CrossRef]
- Prevost, A.; Wilkinson, M.J. A new system of comparing PCR primers applied to ISSR fingerprinting of potato cultivars. Theor. Appl. Genet. 1999, 98, 107–112. [Google Scholar] [CrossRef]
- Azzam, C.R.; Al-Taweel, S.K.; Abdel-Aziz, R.M.; Rabea, K.M.; Abou-Sreea, A.I.B.; Rady, M.M.; Ali, E.F. Salinity Effects on Gene Expression, Morphological, and Physio-Biochemical Responses of Stevia rebaudiana Bertoni In Vitro. Plants 2021, 10, 820. [Google Scholar] [CrossRef]
- Azzam, C.R.; Omran, S.E.H. The promotive effect of PDB biofertilizer on growth, enzymatic activity, and biochemical changes of sunflower (Helianthus annuus L.) plants sprayed with micronutrients. In Proceedings of the 3rd Conference of Recent Technologies in Agriculture, Giza, Egypt, 14–16 November 2005; Volume 2, pp. 255–267. [Google Scholar]
- Omran, S.E.H.; Azzam, C.R. Protein profiles, chemical components and yield of sesame (Sesamum indicum L.) as affected by phosphorous fertilization and foliar application of chelated zinc and calcium. In Proceedings of the 3rd Conference of Recent Technologies in Agriculture, Giza, Egypt, 14–16 November 2005; Volume 2, pp. 239–254. [Google Scholar]
- Ali, M.K.M.; Hamza, J.H. Effect of GA3 on germination characteristics and seedling growth under salt stress in maize. Iraqi J. Agric. Sci. 2014, 45, 6–17. [Google Scholar]
- Abdulradh, M.; Jbara, O.; Shideed, K.; Dhehibi, B. Salinity Impacts on Environmental Efficiency of Wheat farms in Central of Iraq. Iraqi J. Agric. Sci. 2018, 49, 288–294. [Google Scholar]
- Rady, M.M.; Hemida, K.A. Sequenced application of ascorbate-proline-glutathione improves salt tolerance in maize seedlings. Ecotoxicol. Environ. Saf. 2016, 133, 252–259. [Google Scholar] [CrossRef]
- Sairam, R.K.; Tyagi, A. Physiology and molecular biology of salinity stress tolerance in plants. Curr. Sci. 2004, 86, 407–421. [Google Scholar]
- Abd El-Mageed, T.A.; Semida, W.M.; Howladar, S.M.; Zaki, S.S.; Rady, M.M. Hydrogen Peroxide Improves the Antioxidant Defence System in Salt Stressed-Allium cepa Plants. Plant 2016, 4, 91–100. [Google Scholar] [CrossRef]
- Semida, W.M.; Abd El-Mageed, T.A.; Howladar, S.M.; Rady, M.M. Foliar-applied ɑ-tocopherol enhances salt tolerance in onion plants by improving antioxidant defence system. Aust. J. Crop Sci. 2016, 10, 1030–1039. [Google Scholar] [CrossRef]
- Semida, W.M.; Hemida, K.A.; Rady, M.M. Sequenced ascorbate-proline-glutathione seed treatment elevates cadmium tolerance in cucumber transplants. Ecotoxicol. Environ. Saf. 2018, 154, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Rosa, M.; Prado, C.; Podazza, G.; Interdonato, R.; González, J.A.; Hilal, M.; Prado, F.E. Soluble sugars–Metabolism, sensing and abiotic stress:A complex network in the life of plants. Plant Signal. Behav. 2009, 4, 388–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maini, P. The experience of the first biostimulant, based on amino acids and peptides: A short retrospective review on the laboratory researches and the practical results. Fertil. Agrorum 2006, 1, 29–43. [Google Scholar]
- Vernieri, P.; Borghesi, E.; Tognoni, F.; Ferrante, A.; Serra, G.; Piaggesi, A. Use of biostimulants for reducing nutrient solution concentration in floating system. Acta Hortic. 2006, 718, 477–484. [Google Scholar] [CrossRef]
- Abd El-Mageed, T.A.; Semida, W.M.; Rady, M.M. Moringa leaf extract as biostimulant improves water use efficiency, physio-biochemical attributes of squash plants under deficit irrigation. Agric. Water Manag. 2017, 193, 46–54. [Google Scholar] [CrossRef]
- Lemoine, R.; La Camera, S.; Atanassova, R.; Dédaldéchamp, F.; Allario, T.; Pourtau, N.; Bonnemain, J.-L.; Laloi, M.; Coutos-Thévenot, P.; Maurousset, L.; et al. Source-to-sink transport of sugar and regulation by environmental factors—A review. Front. Plant Sci. 2013, 4, 272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rady, M.M.; Taha, R.S.; Mahdi, A. Proline enhances growth, productivity, and anatomy of two varieties of Lupinus termis L. grown under salt stress. S. Afr. J. Bot. 2016, 102, 221–227. [Google Scholar] [CrossRef]
- Semida, W.M.; Rady, M.M. Pre-soaking in 24-epibrassinolide or salicylic acid improves seed germination, seedling growth, and antioxidant capacity in Phaseolus vulgaris L. grown under NaCl stress. J. Hortic. Sci. Biotechnol. 2014, 89, 338–344. [Google Scholar] [CrossRef]
- Rehman, H.; Alharby, H.F.; Bamagoos, A.A.; Abdelhamid, M.T.; Rady, M.M. Sequenced application of glutathione as an antioxidant with organic biostimulant improves physiological and metabolic adaptation to salinity in wheat. Plant Physiol. Biochem. 2021, 158, 43–52. [Google Scholar] [CrossRef]
- Desoky, E.M.; Elrys, A.S.; Rady, M.M. Integrative Moringa and Licorice extracts application improves Performance and reduces fruit contamination content of pepper plants grown on heavy metals-contaminated saline soil. Ecotoxicol. Environ. Saf. 2019, 169, 50–60. [Google Scholar] [CrossRef]
- Rady, M.M.; Elrys, A.S.; Abo El-Maati, M.F.; Desoky, E.M. Interplaying roles of silicon and proline effectively improve salt and cadmium stress tolerance in Phaseolus vulgaris plant. Plant Physiol. Biochem. 2019, 139, 558–568. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Jabborova, D.; Mamadalieva, N. Salt-tolerant Pseudomonas extremorientalis Able to Stimulate Growth of Silybum marianum under Salt Stress. Med. Aromat. Plant Sci. Biotechnol. 2013, 7, 7–10. [Google Scholar]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef] [Green Version]
- Gupta, K.; Sengupta, A.; Chakraborty, M.; Gupta, B. Hydrogen peroxide and polyamines act as double-edged swords in plant abiotic stress responses. Front. Plant Sci. 2016, 7, 1343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.M.; Li, Y.R.; Yang, L.T. cDNA-SCoT: A novel rapid method for analysis of gene differential expression in sugarcane and other plants. Aust. J. Crop Sci. 2013, 7, 659–664. [Google Scholar]
- Sawant, S.V.; Singh, P.K.; Gupta, S.K.; Madnala, R.; Tuli, R. Conserved nucleotide sequences in highly expressed genes in plants. J. Genet. 1999, 78, 123–131. [Google Scholar] [CrossRef]
- Al-Taweel, S.K.; Azzam, C.R.; Khaled, K.A.; Abdel-Aziz, R.M. Improvement of stevia (Stevia rebaudiana Bertoni) and steviol glycoside through traditional breeding and biotechnological approaches. SABRAO J. Breed.Genet. 2021, 53, 88–111. [Google Scholar]
- He, L.; Tang, R.; Jiang, J.; Xiong, F.; Huang, Z.; Wu, H.; GAO, Z.; Zhong, R.; He, X.; Han, Z. Rapid gene expression change in a novel synthesized allopolyploid population of cultivated peanut × Arachis doigoi cross by cDNA-SCoT and HFO-TAG technique. J. Integr. Agric. 2017, 16, 1093–1102. [Google Scholar] [CrossRef] [Green Version]
- Abd EL-Maksoud, R.M.; Ibrahim, S.D.; Faheem, M.M.; Amer, M.N. Analysis of differential gene expression in response to drought stress in wheat (Triticum aestivum L.) using cDNA-SCoT. Egypt. J. Genet. Cytol. 2018, 47, 263–277. [Google Scholar]
- Brudzynski, K.; Sjaarda, C.P. Colloidal structure of honey and its influence on antibacterial activity. Compr. Rev. Food Sci. Food Saf. 2021, 20, 2063–2080. [Google Scholar] [CrossRef]
- Brudzynski, K. A current perspective on hydrogen peroxide production in honey. A review. Food Chem. 2020, 332, 127229. [Google Scholar] [CrossRef] [PubMed]
- Brudzynski, K.; Miotto, D.; Kim, L.; Sjaarda, C.; Maldonado-Alvarez, L.; Fukś, H. Active macromolecules of honey form colloidal particles essential for honey antibacterial activity and hydrogen peroxide production. Sci. Rep. 2017, 7, 7637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brudzynski, K.; Abubaker, K.; Wang, T. Powerful bacterial killing by buckwheat honey is concentration–dependent, involves complete DNA degradation and requires hydrogen peroxide. Front. Microbiol. 2012, 3, 242. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Component | Unit | Value | Reference |
|---|---|---|---|
| Moisture | % | 17.8 | [39] |
| pH | - | 3.8 | |
| Osmoprotectants: | |||
| Proline | μmol kg−1 FW | 359.8 | [39] |
| Total soluble sugars | % | 82.0 | |
| Sugar fractions: | |||
| Fructose | μmol kg−1 FW | 212.0 | [40] |
| Glucose | 168.7 | ||
| Maltose | 12.0 | ||
| Sucrose | 19.0 | ||
| Mineral nutrients: | |||
| Potassium (K) | mmol kg−1 FW | 7.98 | [41] |
| Magnesium (Mg) | 3.14 | ||
| Calcium (Ca) | 1.68 | ||
| Phosphorus (P) | 1.04 | ||
| Iron (Fe) | 0.99 | ||
| Manganese (Mn) | 0.84 | ||
| Zinc (Zn) | 0.08 | ||
| Copper (Cu) | 0.06 | ||
| Iodine (I) | 0.62 | ||
| Antioxidants: | |||
| Ascorbic acid (AsA; Vitamin C) | μmol kg−1 FW | 72.2 | [42] |
| DPPH radical-scavenging activity | % | 88.2 | [43] |
| SCoT Primers | Sequence (5′-3′) | %GC | Molecular Size Range |
|---|---|---|---|
| 1 | CAACAATGGCTACCACCA | 50 | 191:592 |
| 2 | CAACAATGGCTACCACCC | 56 | 395:680 |
| 4 | CAACAATGGCTACCACCT | 50 | 183:421 |
| 7 | CAACAATGGCTACCACGG | 56 | 461:702 |
| 9 | CAACAATGGCTACCAGCA | 50 | 281:640 |
| 10 | CAACAATGGCTACCAGCC | 56 | 555:762 |
| Treatment | Growth Characteristics | |||||
| Shoot FW(g plant−1) | Shoot DW(g plant−1) | Root FW(g plant−1) | Root DW(g plant−1) | |||
| Control | 81.2 ± 5.1 b | 50.2 ± 3.2 b | 23.5 ± 2.1 c | 12.7 ± 1.1 c | ||
| HB | 85.7 ± 5.9 ab | 56.04 ± 0.51 ab | 28.4 ± 3.5 b | 14.5 ± 0.33 b | ||
| HB-Sm | 91.4 ± 6.2 a | 58.8 ± 0.55 a | 31.4 ± 3.8 a | 16.5 ± 0.32 a | ||
| Salinity (S) | 34.1 ± 2.4 d | 23.2 ± 0.22 d | 11.3 ± 1.9 e | 5.2 ± 0.18 e | ||
| S + HB | 51.2 ± 3.7 c | 34.5 ± 0.38 c | 15.2 ± 2.7 d | 8.1 ± 0.22 d | ||
| S + HB-Sm | 77.8 ± 4.9 b | 49.2 ± 0.44 b | 22.8 ± 3.2 c | 12.5 ± 0.22 c | ||
| Yield Parameters | ||||||
| No. of fruits plant−1 | Fresh fruit weights (g plant−1) | Dry fruit weights (g plant−1) | ||||
| Control | 8.2 ± 0.7 b | 14.8 ± 1.5 b | 5.4 ± 0.3 c | |||
| HB | 8.7 ± 0.9 ab | 15.4 ± 1.5 ab | 6.1 ± 0.4 b | |||
| HB-Sm | 9.4 ± 1.0 a | 15.9 ± 1.6 a | 7.1 ± 0.6 a | |||
| Salinity (S) | 4.3 ± 0.5 d | 7.2 ± 0.8 d | 2.1 ± 0.2 e | |||
| S + HB | 6.1 ± 0.6 c | 10.8 ± 0.9 c | 4.1 ± 0.3 d | |||
| S + HB-Sm | 7.8 ± 0.8 b | 14.4 ± 1.2 b | 5.2 ± 0.4 c | |||
| Treatment | T. Chls (mg g−1 FW) | T. Carot (mg g−1 FW) | iCE (µmol m−2 s−1) | Fv/Fm | PhA |
|---|---|---|---|---|---|
| Control | 2.48 ± 0.06 c | 0.62 ± 0.02 c | 0.26 ± 0.01 c | 0.82 ± 0.02 b | 44.8 ± 0.8 c |
| HB | 2.69 ± 0.08 b | 0.69 ± 0.02 b | 0.30 ± 0.01 b | 0.87 ± 0.03 a | 49.9 ± 1.1 b |
| HB-Sm | 2.92 ± 0.09 a | 0.78 ± 0.03 a | 0.33 ± 0.01 a | 0.89 ± 0.03 a | 54.7 ± 1.4 a |
| Salinity (S) | 0.97 ± 0.03 e | 0.35 ± 0.01 e | 0.12 ± 0.00 e | 0.44 ± 0.01 d | 24.2 ± 0.3 e |
| S + HB | 1.92 ± 0.04 d | 0.51 ± 0.01 d | 0.21 ± 0.01 d | 0.73 ± 0.02 c | 36.8 ± 0.6 d |
| S + HB-Sm | 2.39 ± 0.05 c | 0.69 ± 0.02 b | 0.25 ± 0.01 c | 0.80 ± 0.02 b | 43.5 ± 0.7 c |
| Treatment | RWC (%) | Proline Content (μmol g−1 DW) | T.S. Sugars Content (mg g−1 DW) | GB Content (μmol g−1 DW) | Choline Content (μmol g−1 DW) |
|---|---|---|---|---|---|
| Control | 79.8 ± 5.5 b | 94.2 ± 2.5 e | 12.3 ± 0.2 e | 122.4 ± 3.2 f | 198.2 ± 5.2 a |
| HB | 86.9 ± 5.8 a | 105.8 ± 2.8 d | 15.8 ± 0.3 d | 148.6 ± 4.4 e | 176.4 ± 4.0 b |
| HB-Sm | 91.8 ± 7.2 a | 126.6 ± 2.9 c | 19.2 ± 0.4 c | 174.5 ± 4.6 d | 155.3 ± 4.0 b |
| Salinity (S) | 47.2 ± 3.7 d | 129.8 ± 3.2 c | 21.2 ± 0.5 c | 192.6 ± 5.4 c | 129.6 ± 2.6 c |
| S + HB | 70.4 ± 5.2 c | 162.4 ± 3.5 b | 24.4 ± 0.6 b | 214.8 ± 6.7 b | 105.5 ± 2.2 d |
| S + HB-Sm | 78.4 ± 6.0 b | 180.7 ± 3.8 a | 27.7 ± 0.6 a | 240.7 ± 7.4 a | 82.2 ± 1.4 e |
| Treatment | K+ Ion Content (mg g−1 DW) | Na+ Ion Content (mg g−1 DW) | K+/Na+ Ratio | Cl− Ion Content (mg g−1 DW) |
|---|---|---|---|---|
| Control | 2.42 ± 0.05 c | 0.32 ± 0.01 d | 7.56 ± 0.16 c | 0.74 ± 0.02 d |
| HB | 2.70 ± 0.06 b | 0.29 ± 0.01 de | 9.31 ± 0.20 b | 0.72 ± 0.02 d |
| HB-Sm | 2.98 ± 0.06 a | 0.25 ± 0.01 e | 11.92 ± 0.23 a | 0.63 ± 0.01 e |
| Salinity (S) | 1.04 ± 0.02 e | 9.84 ± 0.18 a | 0.11 ± 0.01 f | 12.80 ± 0.10 a |
| S + HB | 2.12 ± 0.04 d | 3.58 ± 0.05 b | 0.59 ± 0.02 e | 4.22 ± 0.08 b |
| S + HB-Sm | 2.40 ± 0.05 bc | 1.04 ± 0.03 c | 2.31 ± 0.05 d | 1.54 ± 0.04 c |
| Treatment | O2•− Level (A580 g−1 FW) | H2O2 Level (μmol g‒1 FW) | MDA Level (A532–600 g−1 FW) | IL (%) | SI (%) |
|---|---|---|---|---|---|
| Control | 0.55 ± 0.01 d | 1.92 ± 0.03 d | 0.18 ± 0.002 d | 6.96 ± 0.30 c | 73.8 ± 4.4 c |
| HB | 0.48 ± 0.01 e | 1.68 ± 0.03 e | 0.14 ± 0.002 e | 6.12 ± 0.24 d | 79.8 ± 4.8 b |
| HB-Sm | 0.41 ± 0.01 f | 1.24 ± 0.03 f | 0.11 ± 0.001 f | 5.42 ± 0.20 e | 85.2 ± 5.5 a |
| Salinity (S) | 1.18 ± 0.02 a | 5.74 ± 0.09 a | 0.42 ± 0.009 a | 24.42 ± 1.31 a | 38.4 ± 2.2 e |
| S + HB | 0.75 ± 0.02 b | 3.02 ± 0.05 b | 0.31 ± 0.006 b | 9.22 ± 0.44 b | 60.2 ± 3.3 d |
| S + HB-Sm | 0.60 ± 0.01 c | 2.18 ± 0.04 c | 0.22 ± 0.002 c | 7.10 ± 0.32 c | 72.3 ± 3.8 c |
| Treatment | AsA Content (μmol g−1 FW) | GSH Content (μmol g−1 FW) | ToC (μmol g−1 DW) | Phenolic Content (mg kg−1 FW) | Capsaicin (mg kg−1 DW) | Sm Content (mg kg−1 DW) |
|---|---|---|---|---|---|---|
| Control | 3.20 ± 0.04 e | 1.31 ± 0.02 f | 46.2 ± 0.4 e | 41.8 ± 0.3 e | 0.84 ± 0.02 d | 25.4 ± 0.4 f |
| HB | 3.58 ± 0.04 d | 1.55 ± 0.02 e | 47.4 ± 0.4 d | 41.9 ± 0.3 e | 0.82 ± 0.02 d | 28.7 ± 0.4 e |
| HB-Sm | 3.96 ± 0.05 c | 1.76 ± 0.03 d | 56.3 ± 0.5 c | 43.3 ± 0.3 d | 0.85 ± 0.02 d | 32.2 ± 0.5 d |
| Salinity (S) | 4.08 ± 0.05 c | 1.98 ± 0.03 c | 58.6 ± 0.6 c | 52.2 ± 0.4 c | 0.94 ± 0.03 c | 37.6 ± 0.6 c |
| S + HB | 4.58 ± 0.06 b | 2.32 ± 0.04 b | 64.8 ± 0.6 b | 57.4 ± 0.5 b | 1.06 ± 0.04 b | 43.2 ± 0.6 b |
| S + HB-Sm | 4.95 ± 0.06 a | 2.62 ± 0.04 a | 71.6 ± 0.6 a | 62.8 ± 0.6 a | 1.22 ± 0.05 a | 51.4 ± 0.8 a |
| Treatment | Activity of SOD (Units mg−1 Protein) | Activity of CAT (Units mg−1 Protein) | Activity of POD (Units mg−1 Protein) | Activity of APX (Units mg−1 Protein) |
|---|---|---|---|---|
| Control | 32.2 ± 0.48 f | 4.94 ± 0.06 f | 14.6 ± 0.19 f | 12.2 ± 0.15 f |
| HB | 35.6 ± 0.55 e | 6.12 ± 0.09 e | 16.9 ± 0.28 e | 13.9 ± 0.18 e |
| HB-Sm | 39.2 ± 0.58 d | 7.68 ± 0.11 d | 18.8 ± 0.32 d | 15.8 ± 0.20 d |
| Salinity (S) | 44.0 ± 0.64 c | 9.62 ± 0.14 c | 22.2 ± 0.38 c | 17.6 ± 0.25 c |
| S + HB | 47.8 ± 0.74 b | 11.44 ± 0.18 b | 25.4 ± 0.42 b | 20.4 ± 0.30 b |
| S + HB-Sm | 52.3 ± 0.84 a | 13.58 ± 0.22 a | 29.8 ± 0.48 a | 24.8 ± 0.36 a |
| Treatment | IAA Content | GAs Content | Cytokinin Content | Salicylic Acid | ABA Content |
|---|---|---|---|---|---|
| (ng g−1 FW) | |||||
| Control | 15.8 ± 0.02 b | 32.1 ± 0.05 b | 17.2 ± 0.03 c | 148 ± 1.8 e | 5.12 ± 0.01 c |
| HB | 17.2 ± 0.03 a | 35.6 ± 0.06 a | 19.8 ± 0.03 b | 169 ± 1.9 d | 3.23 ± 0.01 d |
| HB-Sm | 17.5 ± 0.03 a | 37.0 ± 0.07 a | 22.8 ± 0.04 a | 192 ± 2.4 c | 2.14 ± 0.00 e |
| Salinity (S) | 9.2 ± 0.01 d | 15.7 ± 0.02 d | 8.6 ± 0.02 e | 198 ± 2.5 c | 12.82 ± 0.02 a |
| S + HB | 14.5 ± 0.02 c | 28.2 ± 0.04 c | 13.8 ± 0.03 d | 226 ± 2.9 b | 6.24 ± 0.01 b |
| S + HB-Sm | 16.1 ± 0.03 b | 31.8 ± 0.05 b | 16.9 ± 0.03 c | 255 ± 3.2 a | 5.18 ± 0.01 c |
| Primer Name | cDNA Amplicons | P% | PIC | Rp | ||||
|---|---|---|---|---|---|---|---|---|
| bp Range | McDNA | ScDNA | EcDNA | Total | ||||
| SCoT-1 | 191:592 | 0 | 2 | 4 | 6 | 100.0 | 0.387 | 3.774 |
| SCoT-2 | 195:490 | 0 | 2 | 3 | 5 | 100.0 | 0.387 | 3.774 |
| SCoT-4 | 183:421 | 3 | 4 | 5 | 12 | 75.0 | 0.333 | 2.220 |
| SCoT-7 | 461:702 | 3 | 6 | 2 | 11 | 72.7 | 0.375 | 2.664 |
| SCoT-9 | 305:447 | 3 | 0 | 14 | 17 | 82.4 | 0.311 | 4.440 |
| SCoT-10 | 555:686 | 1 | 0 | 10 | 11 | 91.0 | 0.296 | 2.220 |
| Parameters | Treatments | |||||
|---|---|---|---|---|---|---|
| Control | HB | HB-Sm | Salinity (S) | S + HB | S + HB-Sm | |
| Shoot FW | 81.2 b | +5.54 ab | +12.56 a | −58.00 d | −36.95 c | −4.18 b |
| Shoot DW | 50.2 b | +11.63 ab | +17.13 a | −53.78 d | −31.27 c | −1.99 b |
| Root FW | 23.5 c | +20.85 b | +33.62 a | − 51.91 e | −35.32 d | −2.98 c |
| Root DW | 12.7 c | +14.17 b | +29.92 a | −59.06 e | −36.22 d | −1.57 c |
| No. of fruits plant−1 | 8.2 b | +6.10 ab | +14.63 a | −47.56 d | − 25.61 c | −4.88 b |
| Fresh fruit weights | 14.8 b | +4.05 ab | +7.43 a | −51.35 d | −27.03 c | −2.70 b |
| Dry fruit weights | 5.4 c | +12.96 b | +31.48 a | −61.11 e | −24.07 d | −3.70 c |
| T. Chls. | 2.48 c | +8.47 b | +17.74 a | −60.89 e | −22.58 d | −3.63 c |
| T. Carot. | 0.62 c | +11.29 b | +25.81 a | −43.55 e | − 17.74 d | +11.29 b |
| iEC | 0.26 c | +15.38 b | +26.92 a | −53.85 e | −19.23 d | −3.85 c |
| Fv/Fm | 0.82 b | +6.10 a | +8.54 a | −46.34 d | −10.98 c | −2.44 b |
| Photochemical activity | 44.8 c | +11.38 b | +22.10 a | −45.98 e | − 17.86 d | −2.90 c |
| RWC (%) | 79.8 b | +8.90 a | +15.04 a | −40.85 d | −11.78 c | −1.75 b |
| Proline content | 94.2 e | +12.31 d | +34.39 c | +37.79 c | +72.40 b | +91.83 a |
| T.S. sugars content | 12.3 e | +28.46 d | +56.10 c | +72.36 c | +98.37 b | +125.20 a |
| GB content | 122.4 f | +21.41 e | +42.57 d | +57.35 c | +75.49 b | +96.65 a |
| Choline content | 198.2 a | −11.00 b | −21.64 b | −34.61 c | −46.77 d | −58.53 e |
| K+ ion content | 2.42 c | +11.57 b | +23.14 a | −57.02 e | −12.40 d | −0.83 bc |
| Na+ ion content | 0.32 d | −9.38 de | −21.88 e | +2975.00 a | +1018.75 b | +225.00 c |
| K+/Na+ ratio | 7.56 c | +23.15 b | +57.67 a | −98.54 f | −92.20 e | −69.44 d |
| Cl− ion content | 0.74 d | −2.70 d | −14.86 e | +1629.73 a | +470.27 b | +108.11 c |
| O2•‒ level | 0.55 d | −12.73 e | −25.45 f | +114.55 a | +36.36 b | +9.09 c |
| H2O2 level | 1.92 d | −12.50 e | −35.42 f | +198.96 a | +57.29 b | +13.54 c |
| MDA level | 0.18 d | −22.22 e | −38.89 f | +133.33 a | +72.22 b | +22.22 c |
| IL (%) | 6.96 c | −12.07 d | −22.13 e | +250.86 a | +32.47 b | +2.01 c |
| SI (%) | 73.8 c | +8.13 b | +15.45 a | −47.97 e | −18.43 d | −2.03 c |
| AsA content | 3.20 e | +11.88 d | +23.75 c | +27.50 c | +43.13 b | +54.69 a |
| GSH content | 1.31 f | +18.32 e | +34.35 d | +51.15 c | +77.10 b | +100.00 a |
| α-Tocopherol | 46.2 e | +2.60 d | +21.86 c | +26.84 c | +40.26 b | +54.98 a |
| Phenolic content | 41.8 e | +0.24 e | +3.59 d | +24.88 c | +37.32 b | +50.24 a |
| Capsaicin content | 0.84 d | −2.38 d | +1.19 d | +11.90 c | +26.19 b | +45.24 a |
| Sm content | 25.4 f | +12.99 e | +26.77 d | +48.03 c | +70.08 b | +102.36 a |
| SOD activity | 32.2 f | +10.56 e | +21.74 d | +36.65 c | +48.45 b | +62.42 a |
| CAT activity | 4.94 f | +23.89 e | +55.47 d | +94.74 c | +131.58 b | +174.90 a |
| POD activity | 14.6 f | +15.75 e | +28.77 d | +52.05 c | +73.97 b | +104.11 a |
| APX activity | 12.2 f | +13.93 e | +29.51 d | +44.26 c | +67.21 b | +103.28 a |
| IAA content | 15.8 b | +8.86 a | +10.76 a | −41.77 d | −8.23 c | +1.90 b |
| GAs content | 32.1 b | +10.90 a | +15.26 a | −51.09 d | −12.15 c | −0.93 b |
| Cytokinin content | 17.2 c | +15.12 b | +32.56 a | −50.00 e | −19.77 d | −1.74 c |
| Salicylic acid | 148 e | +14.19 d | +29.73 c | +33.78 c | +52.70 b | +72.30 a |
| ABA content | 5.12 c | −36.91 d | −58.20 e | +150.39 a | +21.88 b | +1.17 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abou-Sreea, A.I.B.; Azzam, C.R.; Al-Taweel, S.K.; Abdel-Aziz, R.M.; Belal, H.E.E.; Rady, M.M.; Abdel-Kader, A.A.S.; Majrashi, A.; Khaled, K.A.M. Natural Biostimulant Attenuates Salinity Stress Effects in Chili Pepper by Remodeling Antioxidant, Ion, and Phytohormone Balances, and Augments Gene Expression. Plants 2021, 10, 2316. https://doi.org/10.3390/plants10112316
Abou-Sreea AIB, Azzam CR, Al-Taweel SK, Abdel-Aziz RM, Belal HEE, Rady MM, Abdel-Kader AAS, Majrashi A, Khaled KAM. Natural Biostimulant Attenuates Salinity Stress Effects in Chili Pepper by Remodeling Antioxidant, Ion, and Phytohormone Balances, and Augments Gene Expression. Plants. 2021; 10(11):2316. https://doi.org/10.3390/plants10112316
Chicago/Turabian StyleAbou-Sreea, Alaa I. B., Clara R. Azzam, Sudad K. Al-Taweel, Ranya M. Abdel-Aziz, Hussein E. E. Belal, Mostafa M. Rady, Atef A. S. Abdel-Kader, Ali Majrashi, and Khaled A. M. Khaled. 2021. "Natural Biostimulant Attenuates Salinity Stress Effects in Chili Pepper by Remodeling Antioxidant, Ion, and Phytohormone Balances, and Augments Gene Expression" Plants 10, no. 11: 2316. https://doi.org/10.3390/plants10112316
APA StyleAbou-Sreea, A. I. B., Azzam, C. R., Al-Taweel, S. K., Abdel-Aziz, R. M., Belal, H. E. E., Rady, M. M., Abdel-Kader, A. A. S., Majrashi, A., & Khaled, K. A. M. (2021). Natural Biostimulant Attenuates Salinity Stress Effects in Chili Pepper by Remodeling Antioxidant, Ion, and Phytohormone Balances, and Augments Gene Expression. Plants, 10(11), 2316. https://doi.org/10.3390/plants10112316