
Double Haploid Development and Assessment of Androgenic Competence of Balkan Pepper Core Collection in Bulgaria

 , and
, and
Abstract
:1. Introduction
2. Results
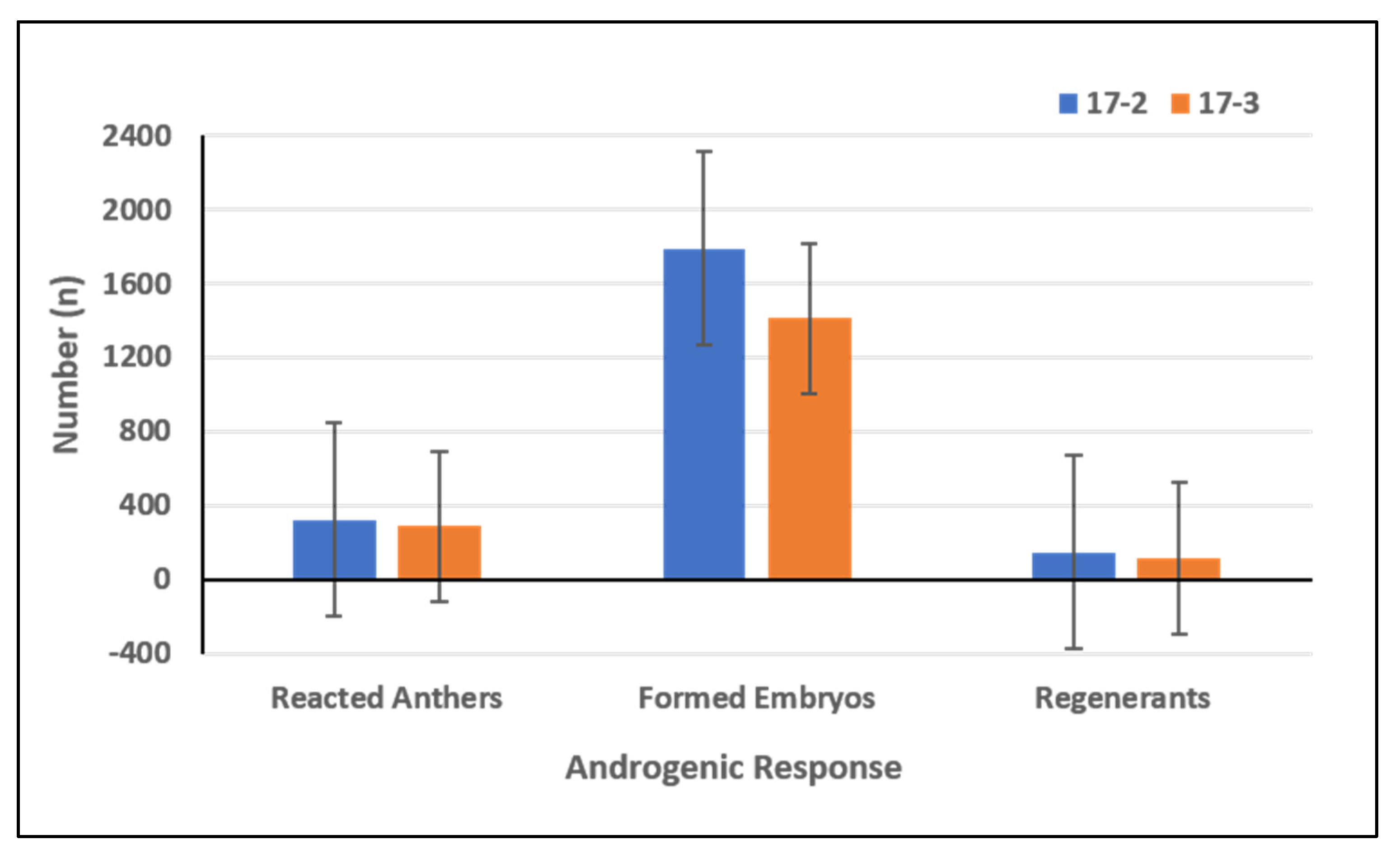
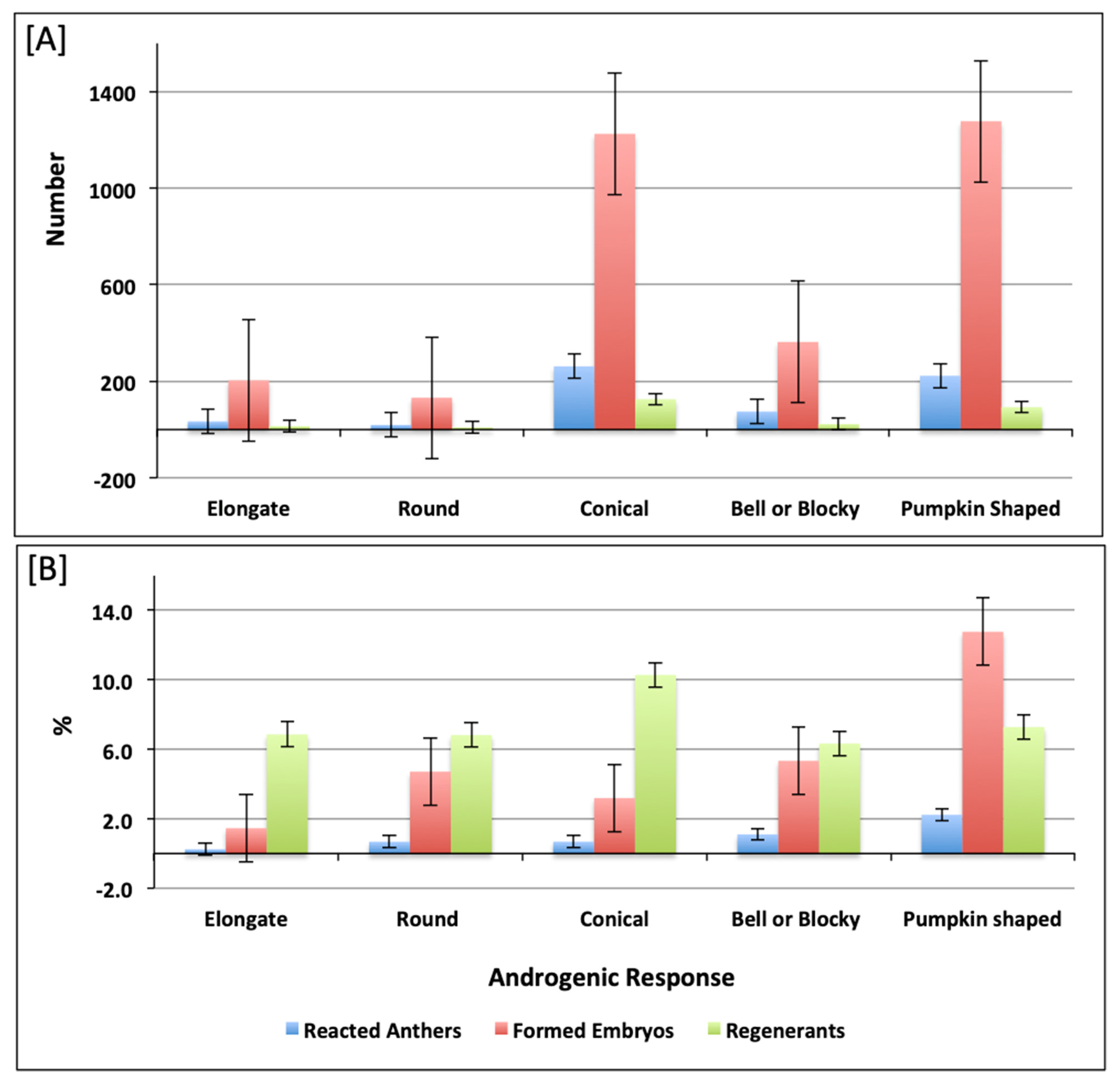
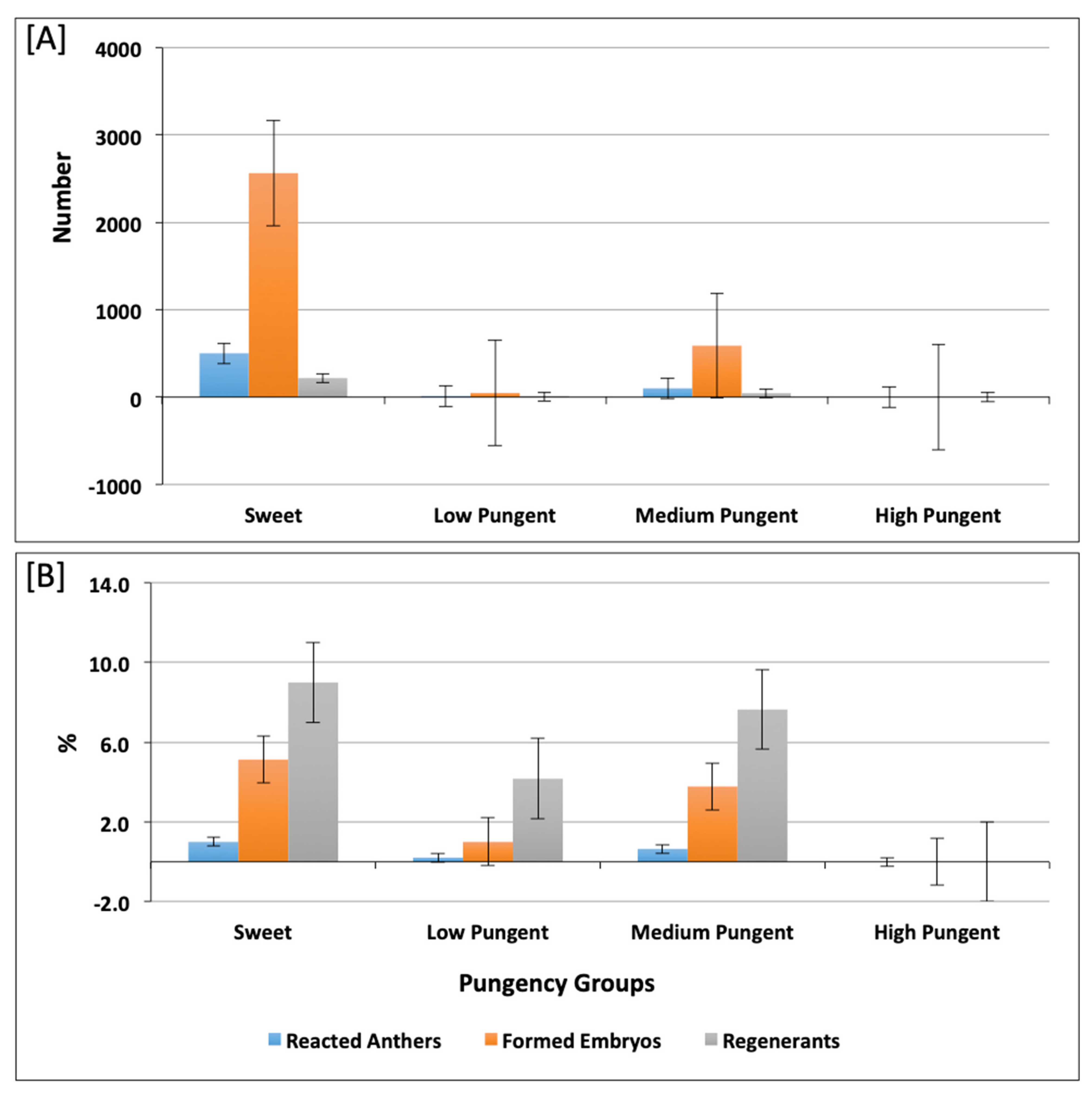
2.1. Assessment of Androgenic Competence
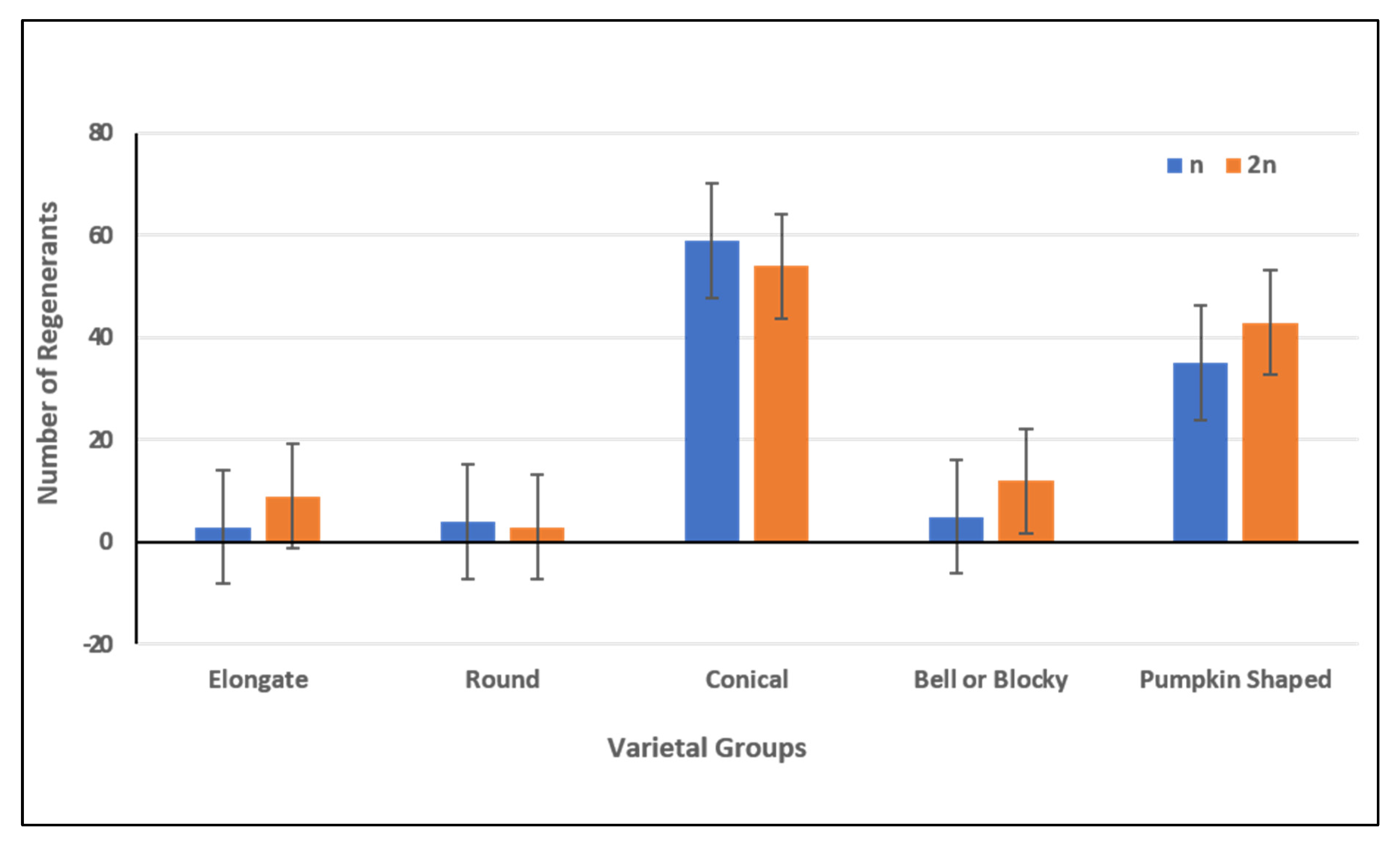
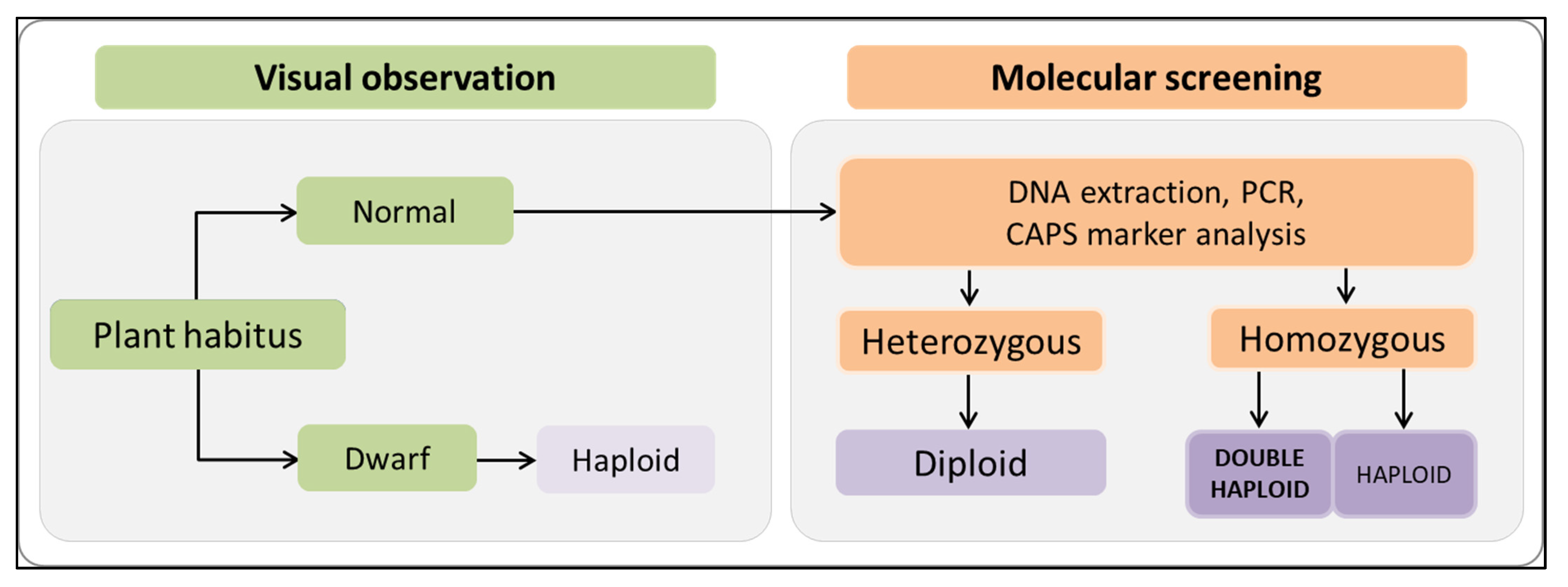
2.2. Discrimination of Haploid and Doubled Haploid Regenerants
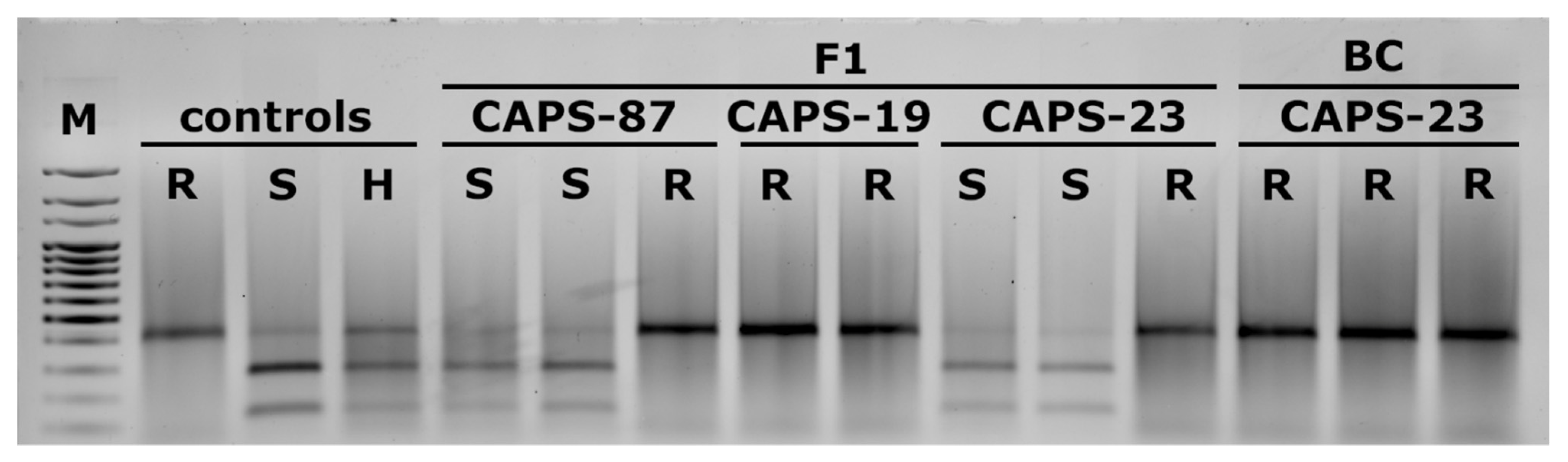
2.3. Identification of Resistance in Doubled Haploid Individuals by Polymerase Chain Reaction (PCR)
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Anther Culture
- 17-2–0.1 mgL−1 2,4-D
- 17-3–0.3 mgL−1 2,4-D
4.3. Flow Cytometry to Analyze Ploidy Level
4.4. DNA Extraction and PCR Analysis
4.5. CAPS Marker Resolving
4.6. Chloroplast Number Counting
4.7. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bosland, P.W.; Votava, E.J. Peppers: Vegetable and Spice Capsicums; CABI Publishing: New York, NY, USA, 2000. [Google Scholar]
- Krasteva, L.; Stoilova, T.; Varbanova, K.; Neykov, S. European Landraces On-Farm Conservation, Management; Vetelainen, M.V., Negri, N.M., Eds.; Biodiversity International: Rome, Italy, 2009; p. 53. [Google Scholar]
- Nankar, A.N.; Todorova, V.; Tringovska, I.; Pasev, G.; Radeva-Ivanova, V.; Ivanova, V.; Kostova, D. A step towards Balkan Capsicum annuum L. core collection: Phenotypic and biochemical characterization of 180 accessions for agronomic, fruit quality, and virus resistance traits. PLoS ONE 2020, 15, e0237741. [Google Scholar] [CrossRef] [PubMed]
- Nankar, A.N.; Tringovska, I.; Grozeva, S.; Todorova, V.; Kostova, D. Application of high-throughput phenotyping tool Tomato Analyzer to characterize Balkan Capsicum fruit diversity. Sci. Hortic. 2019, 260, 108862. [Google Scholar] [CrossRef]
- Krasteva, L.; Pandeva, R.; Rodeva, R.; Todorova, V.; Neykov, S.; Uzundzhalieva, K.; Velcheva, N.; Cvikić, D.; Tome, E.; Ilieva, V. Pepper as a target object of SEE-ERA.NET PROJECT. Acta Hortic. 2012, 960, 151–158. [Google Scholar] [CrossRef]
- Todorova, V.; Djinovic, I. Assessment of Serbian pepper varieties grown in conditions of South Bulgaria. Genetik 2017, 49, 161–172. [Google Scholar] [CrossRef] [Green Version]
- Popova, T.; Kintzios, S.; Grozeva, S.; Rodeva, V. Pepper (Capsicum annuum L.) anther culture—Fundamental research and practical applications. Turk. J. Biol. 2016, 40, 719–726. [Google Scholar] [CrossRef]
- Germanà, M.A. Anther culture for haploid and doubled haploid production. Plant Cell Tiss Org. 2011, 104, 283–300. [Google Scholar] [CrossRef]
- Ochoa-Alejo, N. Anther Culture of Chili Pepper (Capsicum spp.). In Plant Cell Culture Protocols. Methods in Molecular Biology (Methods and Protocols); Loyola-Vargas, V., Ochoa-Alejo, N., Eds.; Humana Press: Totowa, NJ, USA, 2012; Volume 877. [Google Scholar] [CrossRef]
- Forster, B.P.; Heberle-Bors, E.; Kasha, K.J.; Touraev, A. The resurgence of haploids in higher plants. Trends Plant Sci. 2007, 12, 368–375. [Google Scholar] [CrossRef]
- Murovec, J.; Bohanec, B. Haploids and doubled haploids in plant breeding. In Plant Breeding; Abdurakhmonov, I., Ed.; In Tech: Rijeka, Croatia, 2013; pp. 87–105. [Google Scholar]
- Seguı´-Simarro, J.M.; Corral-Martínez, P.; Parra-Vega, V.; González-García, B. Androgenesis in recalcitrant solanaceous crops. Plant Cell Rep. 2011, 30, 765–778. [Google Scholar] [CrossRef]
- Keles, D.; Pınar, H.; Ata, A.; Taskın, H.; Yıldız, S.; Büyükalaca, S. Effect of pepper types on obtaining spontaneous doubled haploid plants via anther culture. HortScience 2015, 50, 1671–1676. [Google Scholar] [CrossRef]
- Grozeva, S.; Todorova, V.; Nankar, A.N. Creation of pepper doubled haploids and morphological characterization of androgenic plants. Euphytica 2021, 217, 113. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Sun, C.S.; Wang, C.C.; Chien, N.J. The induction of pollen plantlets of Triticale and Capsicum annuum anther culture. Sci. Sin. 1973, 16, 147–151. [Google Scholar]
- George, L.; Narayanaswamy, S. Haploid Capsicum through experimental androgenesis. Protoplasma 1973, 78, 467–480. [Google Scholar] [CrossRef]
- Sibi, M.; Dumas de Vaulx, R.; Chambonnet, D. Obtaining haploid plants by in vitro androgenesis in red pepper (Capsicum annuum L.). Ann. L’amelioration Plantes 1979, 29, 583–606. [Google Scholar]
- Dumas de Vaulx, R.; Chambonnet, D.; Pochard, E. In vitro anther culture in red pepper (Capsicum annuum L.): Improvement of the rate of plant production in different genotypes by treatments at 35°C. Agronomie 1981, 1, 859–864. [Google Scholar] [CrossRef]
- Dumas de Vaulx, R. Haploidy and pepper breeding: A review. Capsicum Newsl. 1989, 13–17. [Google Scholar]
- Barroso, P.A.; Rêgo, M.M.; Rêgo, E.R.; Soares, W.S. Embryogenesis in the anthers of different ornamental pepper (Capsicum annuum L.) genotypes. Genet. Mol. Res. 2015, 14, 13349–13363. [Google Scholar] [CrossRef]
- Ari, E.; Bedir, H.; Yildirim, S.; Yildirim, T. Androgenic responses of 64 ornamental pepper (Capsicum annuum L.) genotypes 3 to shed-microspore culture in autumn season. Turk. J. Biol. 2016, 40, 706–717. [Google Scholar] [CrossRef]
- Popova, T.; Grozeva, S.; Todorova, V.; Stankova, G.; Anachkov, N.; Rodeva, V. Effects of low temperature, genotype and culture media on in vitro androgenic answer of pepper (Capsicum annuum L.). Acta Physiol. Plant 2016, 38, 1–11. [Google Scholar] [CrossRef]
- Parra-Vega, V.; Seguı´-Simarro, J.M. Anther culture in pepper (Capsicum annuum L.). In In vitro Embryogenesis in Higher Plants, Vol 1359 of the Series Methods in Molecular Biology; Germana, M., Lambardi, M., Eds.; Springer Science+Business Media: New York, NY, USA, 2016; pp. 467–474. [Google Scholar] [CrossRef]
- Ata, A.; Keleş, D.; Taşkin, H.; Büyükalaca, S. Effects of season, genotype and nutrient medium on pepper anther culture and microspore development. Turk. J. Agric. For. 2018, 43, 123–137. [Google Scholar] [CrossRef]
- Parra-Vega, V.; Renau-Morata, B.; Sifres, A.; Seguí-Simarro, J.M. Stress treatments and in vitro culture conditions influence microspore embryogenesis and growth of callus from anther walls of sweet pepper (Capsicum annuum L.). Plant Cell Tiss Org. Cult. 2013, 112, 353–360. [Google Scholar] [CrossRef]
- Olszewska, D.; Kisiała, A.; Niklas-Nowak, A.; Nowaczyk, P. Study of in vitro anther culture in selected genotypes of genus Capsicum. Turk. J. Biol. 2014, 38, 118–124. [Google Scholar] [CrossRef]
- Boukema, I.W. Allelism of genes controlling resistance to TMV in Capsicum L. Euphytica 1980, 29, 433–439. [Google Scholar] [CrossRef]
- Boukema, I.W. Resistance to TMV in Capsicum chacoense Huns is governed by an allele of the L-Locus. Capsicum. Newsl. 1983, 3, 47–48. [Google Scholar]
- Matsunaga, H.; Saito, T.; Hirai, M.; Nunome, T.; Yoshida, T. DNA Markers Linked to Pepper Mild Mottle Virus (PMMoV) Resistant Locus (L4) in Capsicum. J. Jpn. Soc. Hortic. Sci. 2003, 72, 218–220. [Google Scholar] [CrossRef]
- Yang, H.B.; Liu, W.Y.; Kang, W.H.; Jahn, M.; Kang, B.C. Development of SNP markers linked to the L locus in Capsicum spp. by a comparative genetic analysis. Mol. Breed. 2009, 24, 433–446. [Google Scholar] [CrossRef]
- Yang, H.B.; Liu, W.Y.; Kang, W.H.; Kim, J.H.; Cho, H.J.; Yoo, J.H.; Kang, B.C. Development and validation of L allele-specific markers in Capsicum. Mol. Breed. 2012, 30, 819–829. [Google Scholar] [CrossRef]
- Xu, X.; Li, P.; Zou, C.; Lu, Y.; Xie, C.; Zhang, X.; Prasanna, B.M.; Olsen, M.S. Enhancing genetic gain in the era of molecular breeding. J. Exp. Bot. 2017, 68, 2641–2666. [Google Scholar] [CrossRef] [PubMed]
- Mitykó, J.; Andrásfalvy, A.; Csilléry, G.; Fári, M. Anther-culture response in different genotypes and F1 hybrids of pepper (Capsicum annuum L.). Plant Breed. 1995, 114, 78–80. [Google Scholar] [CrossRef]
- Irikova, T.; Grozeva, S.; Popov, P.; Rodeva, V.; Todorovska, E. In vitro response of pepper anther culture (Capsicum annuum L.) depending on genotype, culture medium and duration of cultivation. Biotechnol. Biotechnol. Equip. 2011, 25, 2604–2609. [Google Scholar] [CrossRef]
- Taskin, H.; Buyukalaka, S.; Keles, D.; Ekbik, E. Induction of microspore-derived embryos by anther culture in selected pepper genotypes. Afr. J. Biotechnol. 2011, 10, 17116–17121. [Google Scholar]
- Luitel, B.; Kang, W. In vitro Androgenic response of minipaprika (Capsicum annuum L.) genotypes in different culture media. Hortic. Environ. Biotechnol. 2013, 54, 162–171. [Google Scholar] [CrossRef]
- Cheng, Y.; Ma, R.; Jiao, Y.; Qiao, N.; Li, T. Impact of genotype, plant growth regulators and activated charcoal on embryogenesis induction in microspore culture of pepper (Capsicum annuum L.). S. Afr. J. Bot. 2013, 88, 306–309. [Google Scholar] [CrossRef] [Green Version]
- Niklas-Nowak, A.; Olszewska, D.; Kisiala, A.; Nowaczyk, P. Study of individual plant responsiveness in anther cultures of selected pepper (Capsicum spp.) genotypes. Folia Hortic. 2012, 24, 141–146. [Google Scholar] [CrossRef] [Green Version]
- Mitykó, J.; Fáry, M. Problems and results of doubled haploid plant production in pepper (Capsicum annuum L.) via anther and microscope culture. Hortic. Biotech. Vitr. Cult. Breed. Acta Hortic. 1997, 447, 281–287. [Google Scholar] [CrossRef]
- Koleva-Gudeva, L.; Gulabovski, R.; Janevik-Ivanovska, E.; Trajkova, F.; Maskimova, V. Capsaicin-inhibitory factor of somatic embryogenesis in pepper anther culture. IEC J. Biol. 2013, 9, 29–36. [Google Scholar]
- Pelliccione, S. Microspore Embryogenesis Induction in Hot Pepper Genotypes. Ph.D. Thesis, University of Granada, Granada, Spain, 2013; p. 20. [Google Scholar]
- Gémesné Juhász, A.; Petus, M.; Venczel, G.; Zatykó, L.; Gyulai, G.; Cséplö, M. variability of anther donor versus spontaneous doubled haploid descendents and colchicine induced doubled haploid sweet pepper (Capsicum annuum L.) lines. Acta Hortic. 2001, 560, 149–152. [Google Scholar] [CrossRef] [Green Version]
- Gyulai, G.; Gémesné, J.A.; Sági, Z.S.; Venczel, G.; Pintér, P.; Kristóf, Z.; Törjék, O.; Heszky, L.; Bottka, S.; Kiss, J.; et al. Doubled Haploid Development and PCR-analysis of F1 Hybrid Derived DH-R2 Paprika (Capsicum annuum L.) Lines. J. Plant Physiol. 2000, 156, 168–174. [Google Scholar] [CrossRef]
- Caranta, C.; Palloix, A.; Lefebvre, V.; Daubèze, A. QTLs for a component of partial resistance to cucumber mosaic virus in pepper: Restriction of virus installation in host-cells. Theor. Appl. Genet. 1997, 94, 431–438. [Google Scholar] [CrossRef]
- Rodeva, V.; Koleva-Gudeva, L.; Grozeva, S.; Traikova, F. Obtaining of haploids in anther culture of pepper Capsicum annuum L. and their including in the breeding process. J. Agric. Plant Sci. 2007, 7, 7–17. [Google Scholar]
- Murashige, S.; Skoog, F. A revised medium for rapid growth and biassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Gamborg, O.L.; Miller, R.A.; Ojima, K. Nutrient requirements of suspension cultures of soybean root cells. Exp. Cell. Res. 1968, 50, 151–158. [Google Scholar] [CrossRef]
- Edwards, K.; Johnstone, C.; Thompson, C. A simple and rapid method for the preparation of plant genomic DNA for PCR analysis. Nucleic Acids Res. 1991, 19, 1349. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Cultivated Anthers | Reacted Anthers | Formed Embryos | Plant Regenerants | ||||
|---|---|---|---|---|---|---|---|---|
| No. | % | No. | % | No. | % | % Towards Embryos | ||
| Accessions | 72,000 | 613 | 0.85 | 3201 | 4.45 | 265 | 0.37 | 8.28 |
| F1 | 3740 | 149 | 3.98 | 787 | 21.04 | 38 | 1.02 | 4.83 |
| BC | 1400 | 128 | 9.14 | 891 | 63.64 | 54 | 3.86 | 6.06 |
| F1 | Reacted Anthers | Formed Embryos | Plant Regenerants | ||||
|---|---|---|---|---|---|---|---|
| No | % | No | % | No | % | % Toward Embryos | |
| F1 hybrids with | |||||||
| CAPS-19 | 13 | 1.76 | 109 | 14.76 | 13 | 1.76 | 11.93 |
| CAPS-23 | 89 | 6.36 | 467 | 33.36 | 17 | 1.21 | 3.64 |
| CAPS-30 | 37 | 6.17 | 165 | 27.50 | 5 | 0.83 | 3.03 |
| CAPS-87 | 10 | 1.00 | 46 | 4.60 | 3 | 0.30 | 6.52 |
| Female parent | |||||||
| CAPS-19 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0.00 |
| CAPS-23 | 44 | 11.00 | 223 | 55.75 | 30 | 7.50 | 13.45 |
| CAPS-30 | 5 | 1.25 | 38 | 9.50 | 2 | 0.50 | 5.26 |
| CAPS-87 | 3 | 0.75 | 12 | 3.00 | 1 | 0.25 | 8.30 |
| BC progeny with CAPS-23 | |||||||
| 782 | 22 | 6.17 | 143 | 27.50 | 13 | 0.83 | 9.09 |
| 787 | 7 | 3.50 | 35 | 17.50 | 3 | 1.50 | 8.57 |
| 788 | 1 | 0.50 | 1 | 0.50 | 0 | 0.00 | 0.00 |
| 881 | 12 | 6.00 | 51 | 25.50 | 3 | 1.50 | 5.88 |
| 882 | 8 | 4.00 | 51 | 25.50 | 5 | 2.50 | 9.80 |
| 886 | 34 | 17.00 | 242 | 121.00 | 10 | 5.00 | 4.13 |
| 887 | 44 | 22.00 | 368 | 184.00 | 20 | 10.00 | 5.43 |
| Varietal Group | Accession Number | Description |
|---|---|---|
| Elongate | CAPS-5, 8, 9, 10, 13, 14, 65, 66, 67, 69, 70, 71, 72, 73, 74, 77, 78, 71A, 109, 112, 113, 123, 124, 125, 130, 132, 137, 139, 144, 149, 153, 155, 166, 174 and 149A | Elongate fruit shape at longitudinal section and fruit length is greater than width. Most fruits are narrow at the fruit base and pointed at the blossom end. |
| Round | CAPS-1, 2, 68, 110, 115, 126 and 128 | Round fruit shape at longitudinal section and fruit length is similar to width. Fruits are blunt at the blossom end. |
| Conical | CAPS-3, 6, 7, 11, 12, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 31, 32, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60,61, 62,63, 64, 75, 76, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 79A, 102, 108, 111, 116, 117, 118, 119, 120, 121, 122, 129, 138, 140, 141, 142, 145, 150, 151, 152, 154, 156, 157, 158, 159, 162, 168, 171, 173, 110A, 117A, 133A, 143A and 151B | Conical fruit shape at longitudinal section and fruit length is several times bigger than the width. Most fruits are pointed at the blossom end. |
| Bell or Blocky | CAPS-29, 30, 92, 93, 94, 95, 133, 134, 135, 136, 143, 146, 147, 148, 160, 161 and 121A | Rectangular or square fruit shape in longitudinal section and fruit length is almost equal to width. Most fruits have 3–5 apexes and are sunken, blunt or sunken and pointed at the blossom end. |
| Pumpkin shaped | CAPS-4, 33, 34, 35, 36, 37, 38, 96, 97, 98, 97A, 101, 103, 104, 105, 107, 114, 127, 131, 163, 164, 165, 169, 135A and 151A | Flattened fruit shape at longitudinal section and fruit length is smaller than width. Fruits are sunken or sunken and pointed at the blossom end. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grozeva, S.; Pasev, G.; Radeva-Ivanova, V.; Todorova, V.; Ivanova, V.; Nankar, A.N. Double Haploid Development and Assessment of Androgenic Competence of Balkan Pepper Core Collection in Bulgaria. Plants 2021, 10, 2414. https://doi.org/10.3390/plants10112414
Grozeva S, Pasev G, Radeva-Ivanova V, Todorova V, Ivanova V, Nankar AN. Double Haploid Development and Assessment of Androgenic Competence of Balkan Pepper Core Collection in Bulgaria. Plants. 2021; 10(11):2414. https://doi.org/10.3390/plants10112414
Chicago/Turabian StyleGrozeva, Stanislava, Gancho Pasev, Vesela Radeva-Ivanova, Velichka Todorova, Valentina Ivanova, and Amol N. Nankar. 2021. "Double Haploid Development and Assessment of Androgenic Competence of Balkan Pepper Core Collection in Bulgaria" Plants 10, no. 11: 2414. https://doi.org/10.3390/plants10112414
APA StyleGrozeva, S., Pasev, G., Radeva-Ivanova, V., Todorova, V., Ivanova, V., & Nankar, A. N. (2021). Double Haploid Development and Assessment of Androgenic Competence of Balkan Pepper Core Collection in Bulgaria. Plants, 10(11), 2414. https://doi.org/10.3390/plants10112414