
Contrasting Hydraulic Efficiency and Photosynthesis Strategy in Differential Successional Stages of a Subtropical Forest in a Karst Region


Abstract
:1. Introduction
2. Materials and Methods
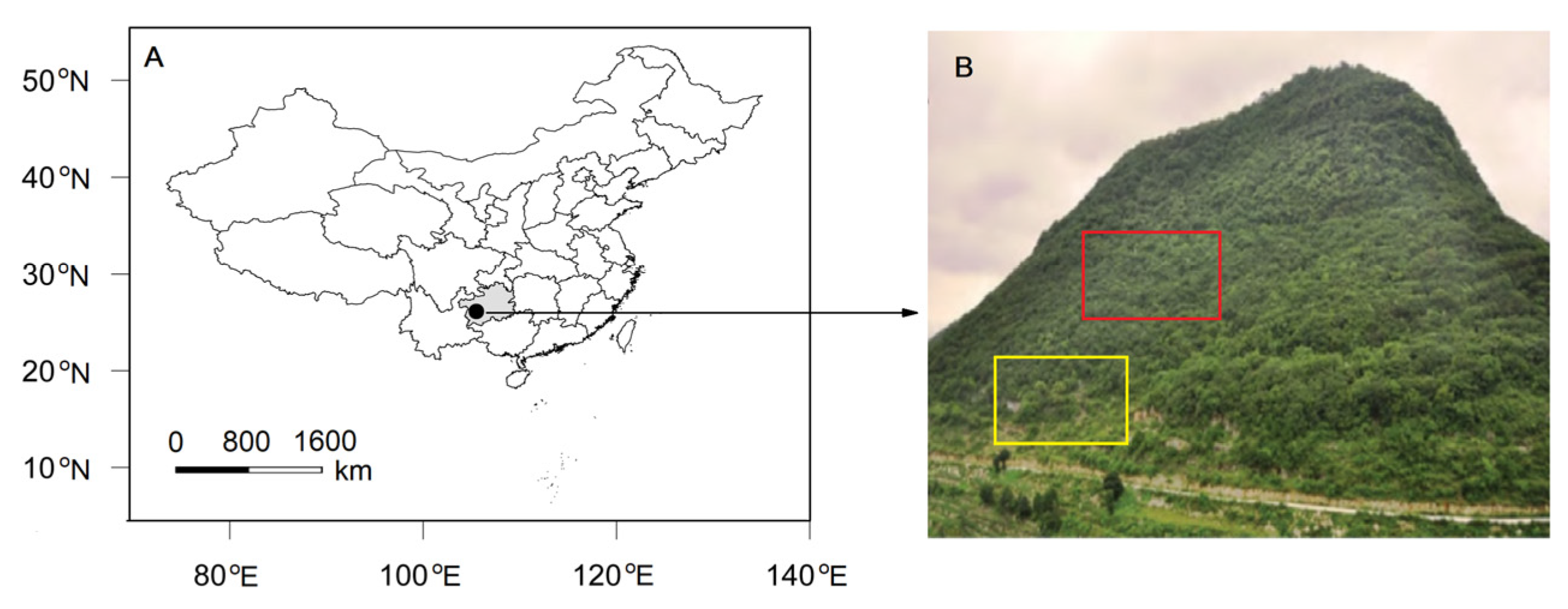
2.1. Study Site and Species
2.2. Gas Exchange Measurement
2.3. Stem Hydraulic Conductivity, Sapwood Density, Specific Leaf Area, and Leaf Area and Sapwood Area Ratio
2.4. Pressure–Volume Relations
2.5. Statistical Analyses
3. Results
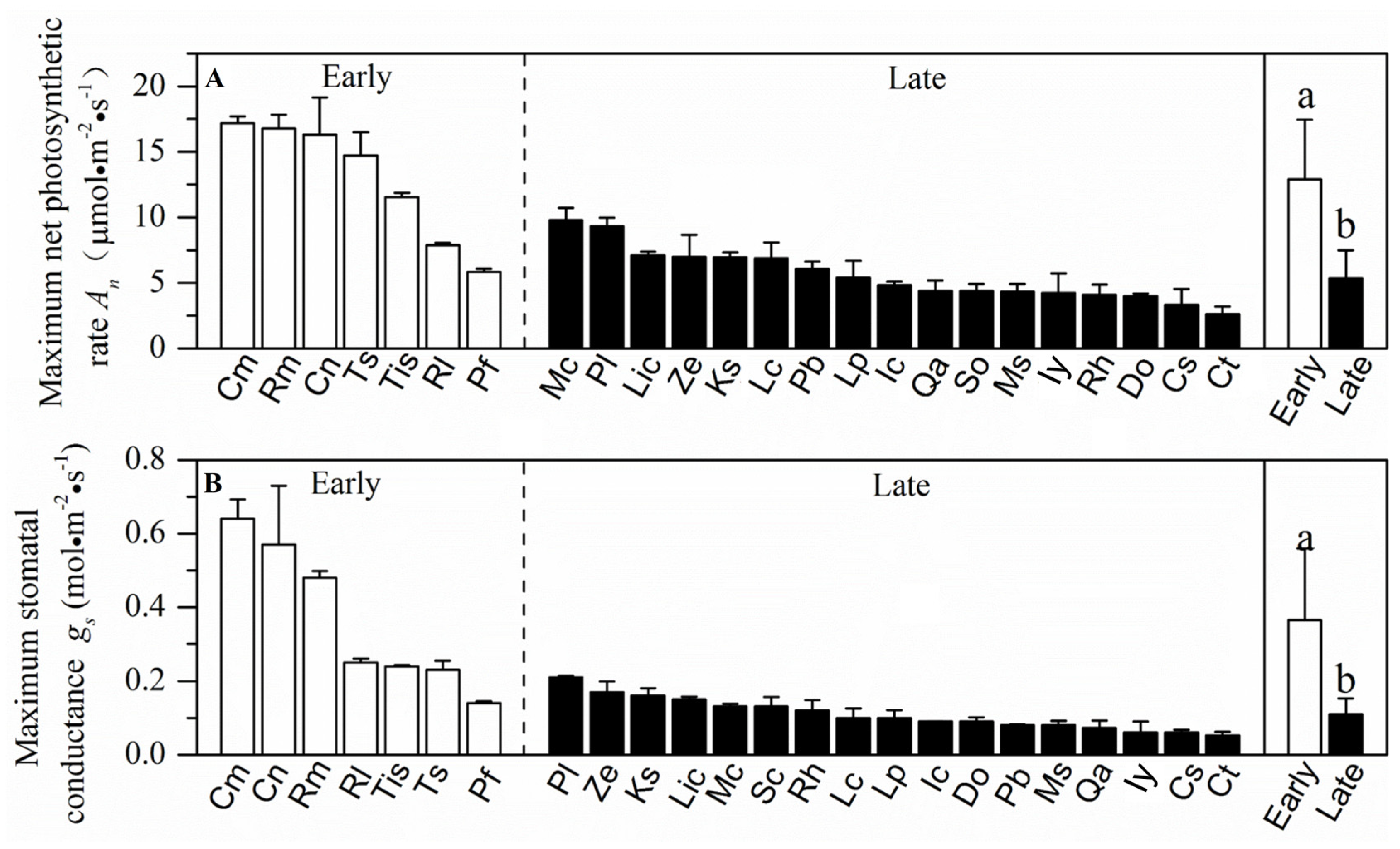
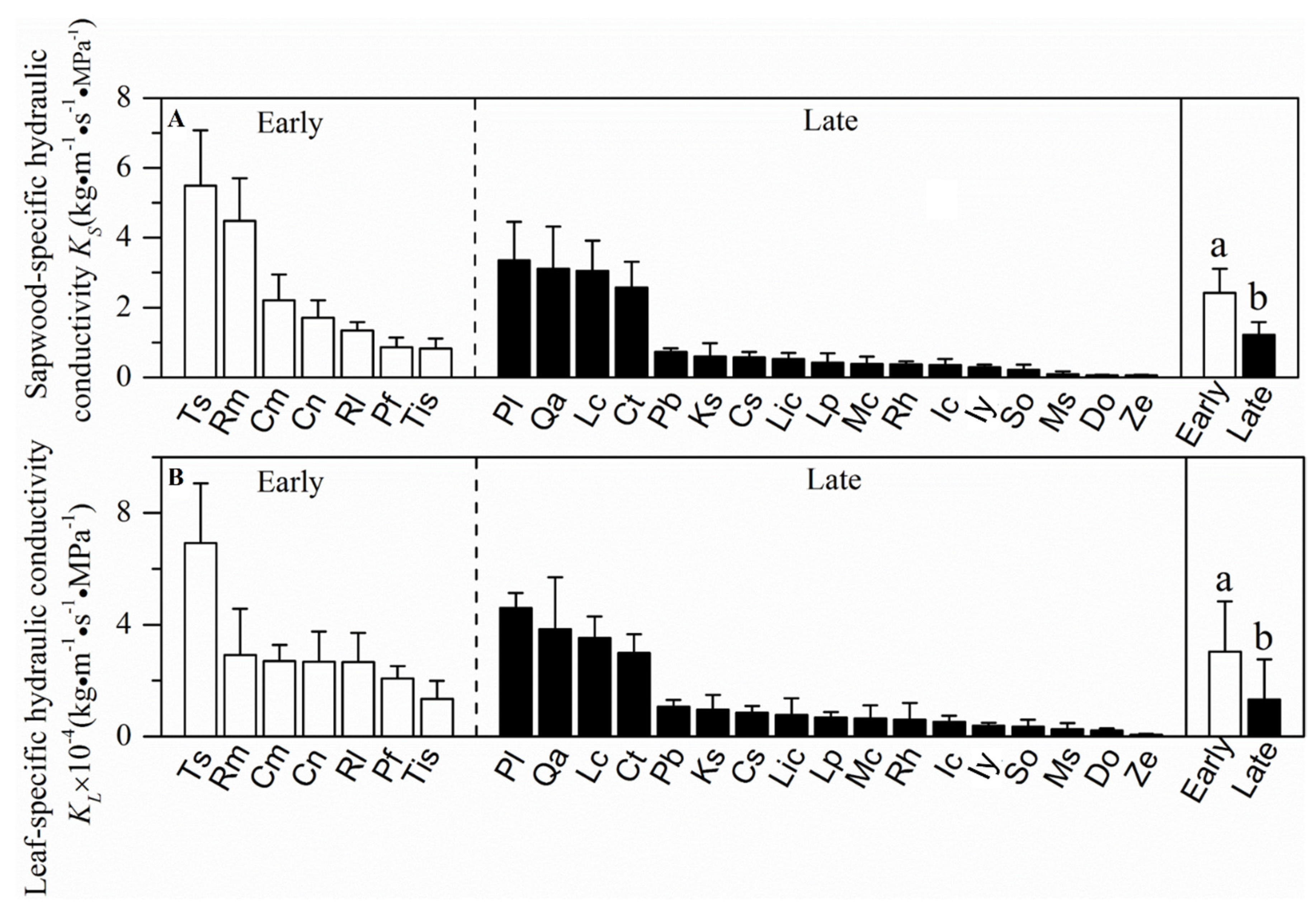
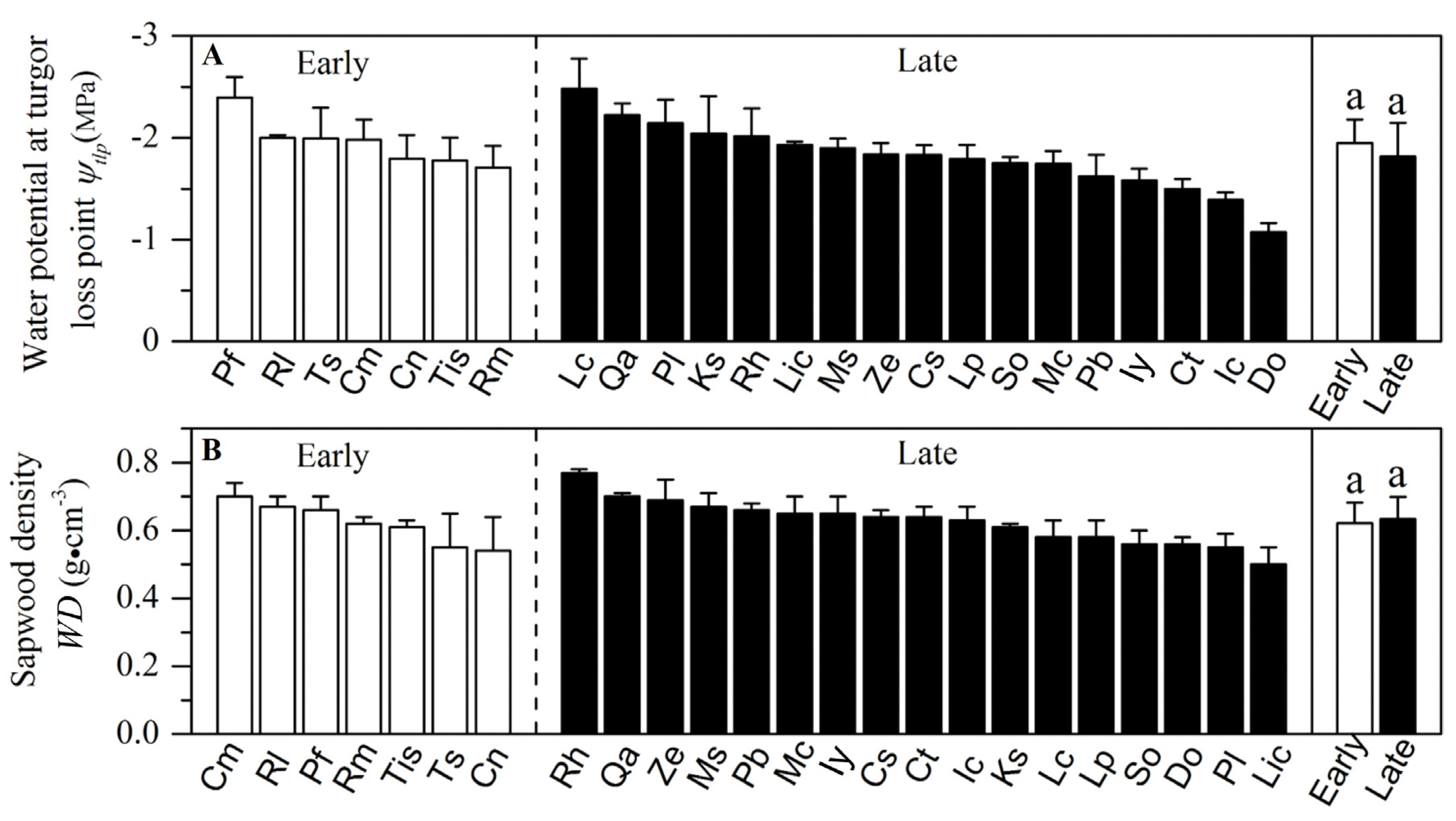
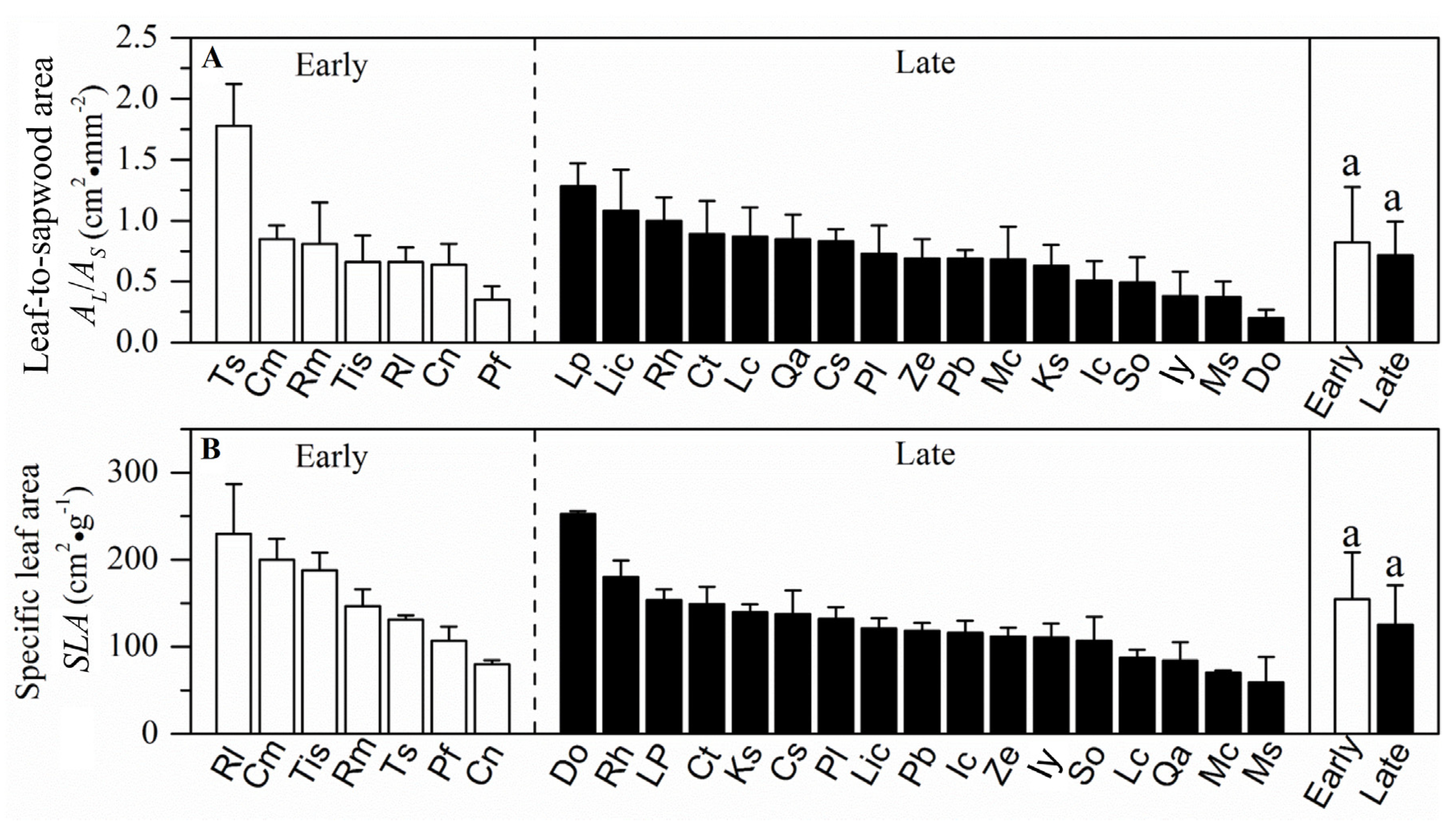
3.1. Resource Acquisition and Drought Tolerance Traits in a Forest at Different Successional Stages
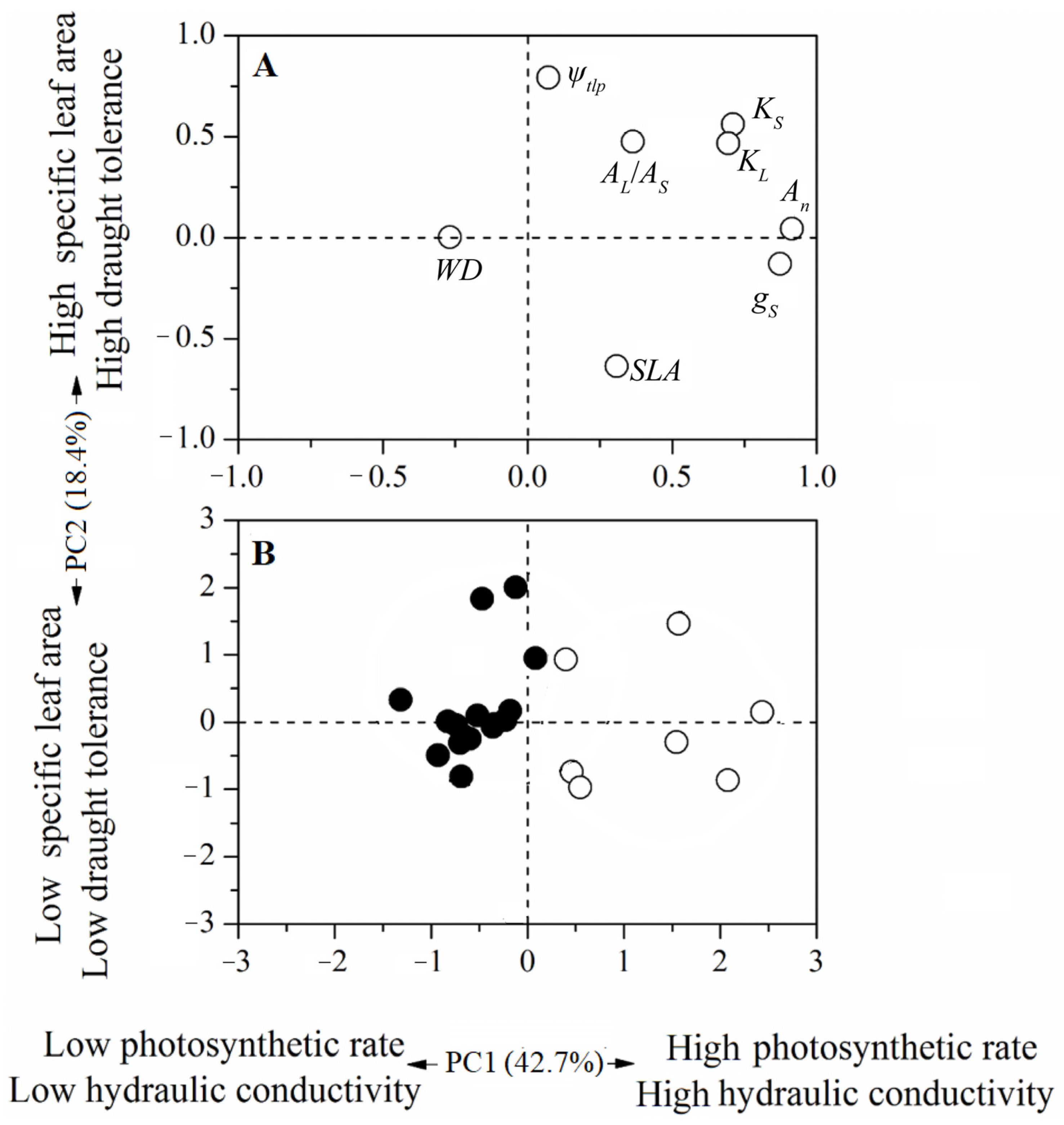
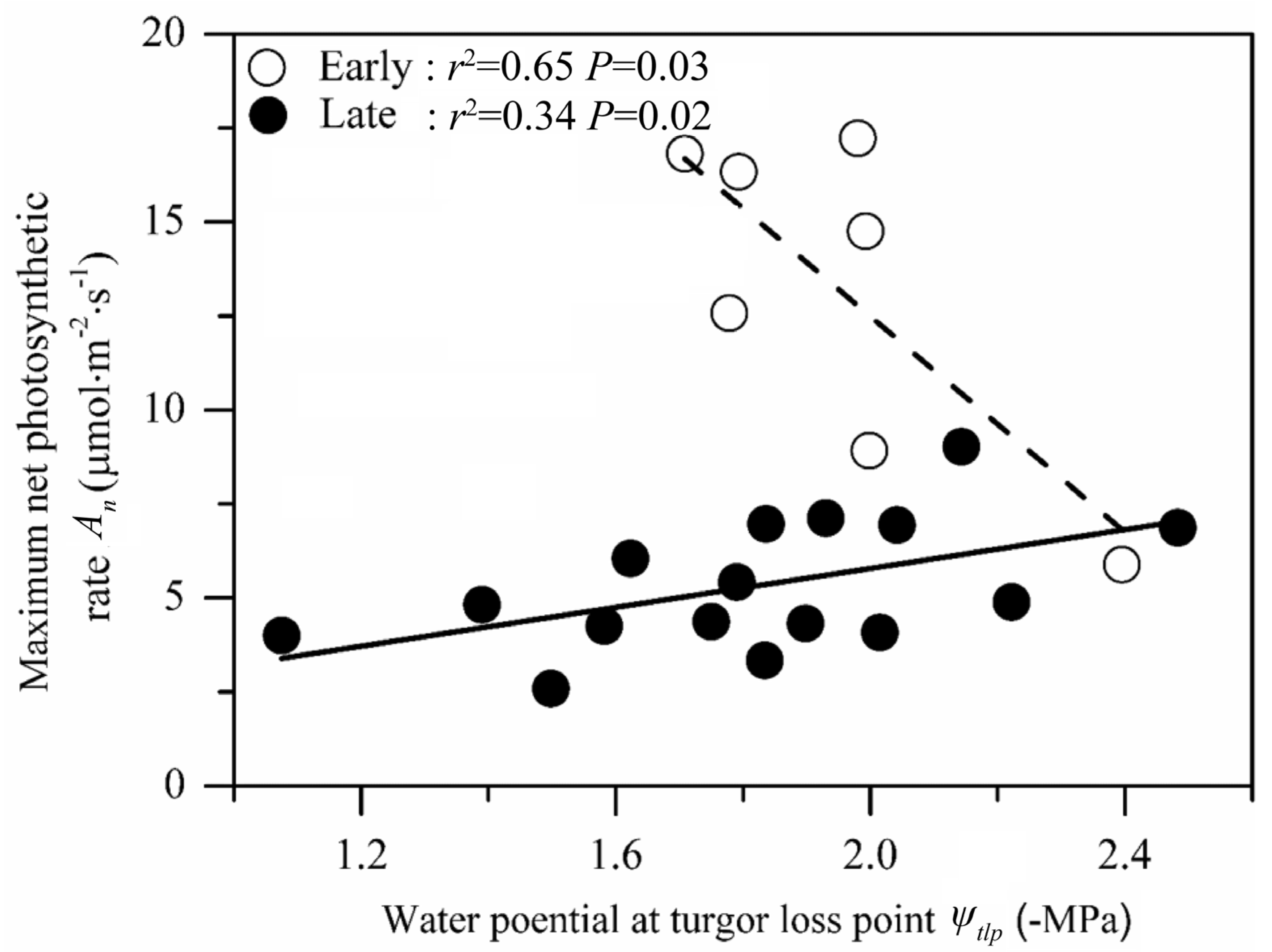
3.2. Relationship between Photosynthetic Rate and Drought Tolerance at Different Successional Stages
4. Discussion
4.1. Resource Acquisition and Drought Tolerance Traits at Different Successional Stages
4.2. Relationship between Photosynthetic Rate and Drought Tolerance at Different Successional Stages
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clements, R.; Sodhi, N.S.; Schilthuizen, M.; Ng, P.K.L. Limestone karsts of southeast Asia: Imperiled arks of biodiversity. Bioscience 2006, 56, 733–742. [Google Scholar] [CrossRef] [Green Version]
- Shi, K.; Yang, Q.; Li, Y. Are karst rocky desertification areas affected by increasing human activity in southern China? An empirical analysis from nighttime light data. Int. J. Environ. Res. Public Heal. 2019, 16, 4175. [Google Scholar] [CrossRef] [Green Version]
- Sun, Z.G.; Wu, J.; Liu, F.; Shao, T.Y.; Liu, X.B.; Chen, Y.Z.; Long, X.H.; Rengel, Z. Quantitatively assessing the effects of climate change and human activities on ecosystem degradation and restoration in southwest China. Rangel. J. 2019, 41, 335. [Google Scholar] [CrossRef]
- Xu, E.; Zhang, H.; Li, M. Mining spatial information to investigate the evolution of karst rocky desertification and its human driving forces in Changshun, China. Sci. Total. Environ. 2013, 458–460, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Poorter, L. Leaf traits show different relationships with shade tolerance in moist versus dry tropical forests. New Phytol. 2009, 181, 890–900. [Google Scholar] [CrossRef]
- Zhu, S.-D.; Song, J.-J.; Li, R.-H.; Ye, Q. Plant hydraulics and photosynthesis of 34 woody species from different successional stages of subtropical forests. Plant. Cell Environ. 2013, 36, 879–891. [Google Scholar] [CrossRef] [PubMed]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef]
- Zhu, S.-D.; Li, R.-H.; Song, J.; He, P.-C.; Liu, H.; Berninger, F.; Ye, Q. Different leaf cost–benefit strategies of ferns distributed in contrasting light habitats of sub-tropical forests. Ann. Bot. 2016, 117, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Liu, M.; Lu, J.; Yang, H. Effects of shading on the growth and leaf photosynthetic characteristics of three forages in an apple orchard on the Loess Plateau of eastern Gansu, China. Peer J. 2019, 7, e7594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartlett, M.K.; Scoffoni, C.; Sack, L. The determinants of leaf turgor loss point and prediction of drought tolerance of species and biomes: A global meta-analysis. Ecol. Lett. 2012, 15, 393–405. [Google Scholar] [CrossRef]
- Bartlett, M.K.; Zhang, Y.; Kreidler, N.; Sun, S.; Ardy, R.C.; Cao, K.-F.; Sack, L. Global analysis of plasticity in turgor loss point, a key drought tolerance trait. Ecol. Lett. 2014, 17, 1580–1590. [Google Scholar] [CrossRef] [PubMed]
- Sack, L.; Cowan, P.; Jaikumar, N.; Holbrook, N. The ‘hydrology’ of leaves: Coordination of structure and function in temperate woody species. Plant. Cell Environ. 2003, 26, 1343–1356. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.; Ye, Q.; Liu, H.; Brodribb, T.J. Wood density predicts mortality threshold for diverse trees. New Phytol. 2020, 229, 3053–3057. [Google Scholar] [CrossRef]
- Lenz, T.I.; Wright, I.; Westoby, M. Interrelations among pressure-volume curve traits across species and water availability gradients. Physiol. Plant. 2006, 127, 423–433. [Google Scholar] [CrossRef]
- Merchant, A.; Callister, A.; Arndt, S.; Tausz, M.; Adams, M. Contrasting Physiological Responses of Six Eucalyptus Species to Water Deficit. Ann. Bot. 2007, 100, 1507–1515. [Google Scholar] [CrossRef] [PubMed]
- Bucci, S.J.; Scholz, F.G.; Campanello, P.I.; Montti, L.; Jimenez-Castillo, M.; Rockwell, F.A.; Manna, L.L.; Guerra, P.; Bernal, P.L.; Troncoso, O. Hydraulic differences along the water transport system of South American Nothofagus species: Do leaves protect the stem functionality? Tree physiol. 2012, 32, 880–893. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, P.J.; O’Grady, A.P. Adaptation of leaf water relations to climatic and habitat water availability. Forests 2015, 6, 2281–2295. [Google Scholar] [CrossRef]
- Oliveira, R.S.; Eller, C.B.; Barros, F.D.V.; Hirota, M.; Brum, M.; Bittencourt, P. Linking plant hydraulics and the fast–slow continuum to understand resilience to drought in tropical ecosystems. New Phytol. 2021, 230, 904–923. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, N.L.; van Mantgem, P.J.; Bunn, A.G.; Bruner, H.; Harmon, M.E.; O’Connell, K.B.; Urban, D.L.; Franklin, J.F. Causes and implications of the correlation between forest productivity and tree mortality rates. Ecol. Monogr. 2011, 81, 527–555. [Google Scholar] [CrossRef] [Green Version]
- Wright, S.J.; Kitajima, K.; Kraft, N.J.; Reich, P.B.; Wright, I.J.; Bunker, D.E.; Condit, R.; Dalling, J.W.; Davies, S.J.; Diaz, S.; et al. Functional traits and the growth–mortality trade-off in tropical trees. Ecology 2010, 91, 3664–3674. [Google Scholar] [CrossRef]
- Markesteijn, L.; Poorter, L.; Paz, H.; Sack, L.; Bongers, F. Ecological differentiation in xylem cavitation resistance is associated with stem and leaf structural traits. Plant. Cell Environ. 2011, 34, 137–148. [Google Scholar] [CrossRef]
- Poorter, L.; Bongers, F. Leaf traits are good predictors of plant performance across 53 rain forest species. Ecology 2006, 87, 1733–1743. [Google Scholar] [CrossRef]
- De Guzman, M.E.; Santiago, L.S.; Schnitzer, S.A.; Álvarez-Cansino, L. Trade-offs between water transport capacity and drought resistance in neotropical canopy liana and tree species. Tree Physiol. 2016, 37, 1404–1414. [Google Scholar] [CrossRef]
- Hacke, U.G.; Sperry, J.S.; Wheeler, J.K.; Castro, L. Scaling of angiosperm xylem structure with safety and efficiency. Tree Physiol. 2006, 26, 689–701. [Google Scholar] [CrossRef]
- Sande, M.T.; Poorter, L.; Schnitzer, T.A.; Engelbrecht, B.M.; Markesteijn, L. The hydraulic efficiency–safety trade-off differs between lianas and trees. Ecology 2019, 100, e02666. [Google Scholar] [CrossRef] [Green Version]
- Fu, Z.; Chen, H.; Zhang, W.; Xu, Q.; Wang, S.; Wang, K. Subsurface flow in a soil-mantled subtropical dolomite karst slope: A field rainfall simulation study. Geomorphology 2015, 250, 1–14. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, H.; Su, Y.; Kong, X.; Zhang, W.; Shi, Y.; Liang, H.; Shen, G. Spatial variability and patterns of surface soil moisture in a field plot of karst area in southwest China. Plant. Soil Environ. 2011, 57, 409–417. [Google Scholar] [CrossRef] [Green Version]
- Fu, P.-L.; Jiang, Y.-J.; Wang, A.-Y.; Brodribb, T.; Zhang, J.-L.; Zhu, S.-D.; Cao, K.-F. Stem hydraulic traits and leaf water-stress tolerance are co-ordinated with the leaf phenology of angiosperm trees in an Asian tropical dry karst forest. Ann. Bot. 2012, 110, 189–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, S.-D.; Chen, Y.-J.; Fu, P.-L.; Cao, K.-F. Different hydraulic traits of woody plants from tropical forests with contrasting soil water availability. Tree Physiol. 2017, 37, 1469–1477. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhu, S.; Jansen, S.; Cao, K. Topography strongly affects drought stress and xylem embolism resistance in woody plants from a karst forest in Southwest China. Funct. Ecol. 2020, 35, 566–577. [Google Scholar] [CrossRef]
- Liu, C.C.; Wei, Y.F.; Liu, Y.G.; Guo, K. Biomass of canopy and shrub layers of karst forests in Puding, Guizhou, China. Chin. J. Plant. Ecol. 2009, 33, 698–705. [Google Scholar]
- Liu, Y.G.; Liu, C.C.; Wei, Y.F.; Liu, Y.G.; Guo, K. Species composition and community structure at different vegetation successional stages in Puding, Guizhou Province, China. Chin. J. Plant. Ecol. 2011, 35, 1009–1018. [Google Scholar]
- Liu, L.; Ni, J.; Zhong, Q.; Hu, G.; Zhang, Z. High mortality and low net change in live woody biomass of karst evergreen and deciduous broad-leaved mixed forest in southwestern China. Forests 2018, 9, 263. [Google Scholar] [CrossRef] [Green Version]
- Sperry, J.S.; Donnelly, J.R.; Tyree, M.T. A method for measuring hydraulic conductivity and embolism in xylem. Plant Cell Environ. 1988, 11, 35–40. [Google Scholar] [CrossRef]
- Schulte, P.; Hinckley, T.M. A comparison of pressure-volume curve data analysis techniques. J. Exp. Bot. 1985, 36, 1590–1602. [Google Scholar] [CrossRef]
- RStudio-Team. RStudio: Integrated Development for RStudio, Inc.; RStudio Inc.: Boston, MA, USA, 2016. [Google Scholar]
- McCulloh, K.A.; Meinzer, F.C.; Sperry, J.S.; Lachenbruch, B.; Voelker, S.L.; Woodruff, D.R.; Domec, J.-C. Comparative hydraulic architecture of tropical tree species representing a range of successional stages and wood density. Oecologia 2011, 167, 27–37. [Google Scholar] [CrossRef]
- Feild, T.S.; Brodribb, T. Stem water transport and freeze-thaw xylem embolism in conifers and angiosperms in a Tasmanian treeline heath. Oecologia 2001, 127, 314–320. [Google Scholar] [CrossRef]
- Santiago, L.S.; Kitajima, K.; Wright, S.J.; Mulkey, S.S. Coordinated changes in photosynthesis, water relations and leaf nutritional traits of canopy trees along a precipitation gradient in lowland tropical forest. Oecologia 2004, 139, 495–502. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.-L.; Cao, K.-F. Stem hydraulics mediates leaf water status, carbon gain, nutrient use efficiencies and plant growth rates across dipterocarp species. Funct. Ecol. 2009, 23, 658–667. [Google Scholar] [CrossRef]
- Coley, P.D.; Bryant, J.P.; Chapin, F.S. Resource availability and plant antiherbivore defense. Science 1985, 230, 895–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wheeler, J.K.; Sperry, J.S.; Hacke, U.G.; Hoang, N. Inter-vessel pitting and cavitation in woody Rosaceae and other vesselled plants: A basis for a safety versus efficiency trade-off in xylem transport. Plant. Cell Environ. 2005, 28, 800–812. [Google Scholar] [CrossRef]
- Dong, X.; Zhang, X. Some observations of the adaptations of sandy shrubs to the arid environment in the Mu Us Sandland: Leaf water relations and anatomic features. J. Arid. Environ. 2001, 48, 41–48. [Google Scholar] [CrossRef]
- Vendramini, F.; Díaz, S.; Gurvich, D.E.; Wilson, P.J.; Thompson, K.; Hodgson, J.G. Leaf traits as indicators of resource-use strategy in floras with succulent species. New Phytol. 2002, 154, 147–157. [Google Scholar] [CrossRef]
- Baltzer, J.L.; Davies, S.J.; Bunyavejchewin, S.; Noor, N.S.M. The role of desiccation tolerance in determining tree species distributions along the Malay–Thai Peninsula. Funct. Ecol. 2008, 22, 221–231. [Google Scholar] [CrossRef]
- Reich, P.B.; Cornelissen, H. The world-wide ‘fast-slow’ plant economics spectrum: A traits manifesto. J. Ecol. 2014, 102, 275–301. [Google Scholar] [CrossRef]
- Litvak, E.; McCarthy, H.R.; Pataki, D.E. Transpiration sensitivity of urban trees in a semi-arid climate is constrained by xylem vulnerability to cavitation. Tree Physiol. 2012, 32, 373–388. [Google Scholar] [CrossRef] [Green Version]
- Sperry, J.S.; Nichols, K.L.; Sullivan, J.E.M.; Eastlack, S.E. Xylem embolism in ring-porous, diffuse-porous, and coniferous trees of northern Utah and interior Alaska. Ecology 1994, 75, 1736–1752. [Google Scholar] [CrossRef]
- Pineda-García, F.; Paz, H.; Meinzer, F.C.; Ángeles, G. Exploiting water versus tolerating drought: Water-use strategies of trees in a secondary successional tropical dry forest. Tree Physiol. 2016, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González, M.R.; Posada, J.M.; Carmona, C.P.; Garzón, F.; Salinas, V.; Idárraga-Piedrahita, P.C.; Avella, A.; López-Camacho, R.; Norden, N. Diverging functional strategies but high sensitivity to an extreme drought in tropical dry forests. Ecol. Lett. 2021, 24, 451–463. [Google Scholar] [CrossRef]
- Yao, G.; Nie, Z.; Turner, N.C.; Li, F.; Gao, T.; Fang, X.; Scoffoni, C. Combined high leaf hydraulic safety and efficiency provides drought tolerance in Caragana species adapted to low mean annual precipitation. New Phytol. 2020, 229, 230–244. [Google Scholar] [CrossRef]
- Liu, H.; Ye, Q.; Gleason, S.M.; He, P.; Yin, D. Weak tradeoff between xylem hydraulic efficiency and safety: Climatic sea-sonality matters. New Phytol. 2021, 229, 1440–1452. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Family | Code | Sampling Site | Successional Stage |
|---|---|---|---|---|
| Campylotropis macrocarpa | Fabaceae | Cm | Open site | Early |
| Coriaria nepalensis | Coriariaceae | Cn | Open site | Early |
| Tirpitzia sinensis | Linaceae | Tis | Open site | Early |
| Rhamnus leptophylla | Rhamnaceae | Rl | Forest margins | Early |
| Pyracantha fortuneana | Rosaceae | Pf | Open site | Early |
| Rhamnella martini | Rhamnaceae | Rm | Open site | Early |
| Toona sinensis | Meliaceae | Tos | Open site | Early |
| Pittosporum brevicalyx | Pittosporaceae | Pb | Understory | Late |
| Ilex coralline | Aquifoliaceae | Ic | Understory | Late |
| Stachyurus obovatus | Stachyuraceae | So | Understory | Late |
| Myrsine semiserrata | Primulaceae | Ms | Understory | Late |
| Itea yunnanensis | Iteaceae | Iy | Understory | Late |
| Rhamnus heterophylla | Rhamnaceae | Rh | Understory | Late |
| Daphne odora | Thymelaeaceae | Do | Understory | Late |
| Machilus cavaleriei | Lauraceae | Mc | Understory | Late |
| Kalopanax septemlobus | Araliaceae | Ks | Forest canopy | Late |
| Lithocarpus confinis | Fagaceae | Lic | Forest canopy | Late |
| Lindera pulcherrima | Lauraceae | Lp | Forest canopy | Late |
| Celtis sinensis | Cannabaceae | Cs | Forest canopy | Late |
| Carpinus turczaninowii | Betulaceae | Ct | Forest canopy | Late |
| Platycarya longipea | Juglandaceae | Pl | Forest canopy | Late |
| Quercus acutissima | Fagaceae | Qa | Forest canopy | Late |
| Lindera communis | Lauraceae | Lc | Forest canopy | Late |
| Zanthoxylum esquirolii | Rutaceae | Ze | Forest canopy | Late |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, G.; Chen, D.; Zhou, Z. Contrasting Hydraulic Efficiency and Photosynthesis Strategy in Differential Successional Stages of a Subtropical Forest in a Karst Region. Plants 2021, 10, 2604. https://doi.org/10.3390/plants10122604
Wu G, Chen D, Zhou Z. Contrasting Hydraulic Efficiency and Photosynthesis Strategy in Differential Successional Stages of a Subtropical Forest in a Karst Region. Plants. 2021; 10(12):2604. https://doi.org/10.3390/plants10122604
Chicago/Turabian StyleWu, Guilin, Dexiang Chen, and Zhang Zhou. 2021. "Contrasting Hydraulic Efficiency and Photosynthesis Strategy in Differential Successional Stages of a Subtropical Forest in a Karst Region" Plants 10, no. 12: 2604. https://doi.org/10.3390/plants10122604
APA StyleWu, G., Chen, D., & Zhou, Z. (2021). Contrasting Hydraulic Efficiency and Photosynthesis Strategy in Differential Successional Stages of a Subtropical Forest in a Karst Region. Plants, 10(12), 2604. https://doi.org/10.3390/plants10122604