
LED and HPS Supplementary Light Differentially Affect Gas Exchange in Tomato Leaves



Abstract
:1. Introduction
2. Results
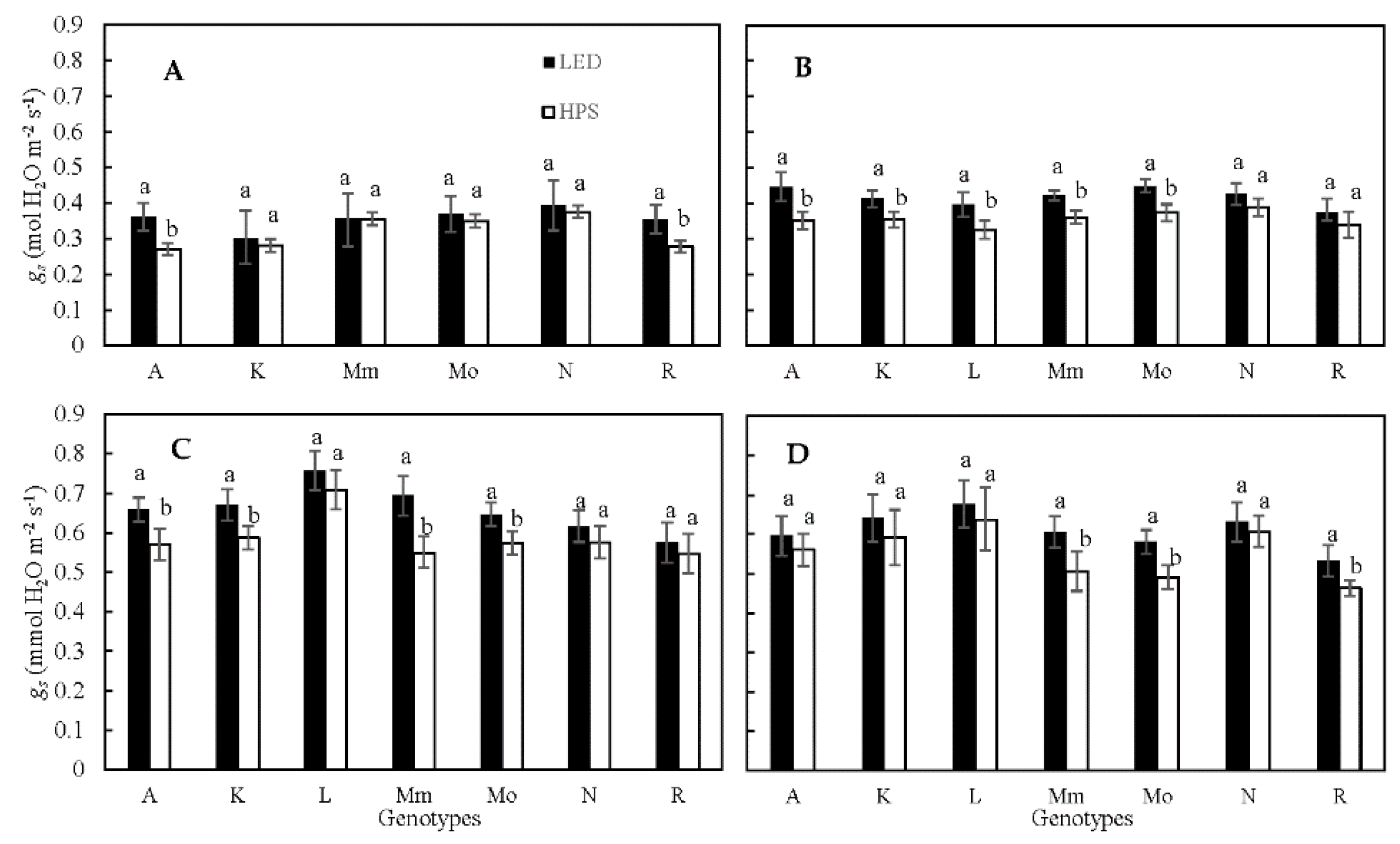
2.1. Stomatal Conductance (gs) and Transpiration Rate (E)
2.2. Stomatal Density
2.3. Leaf Temperature
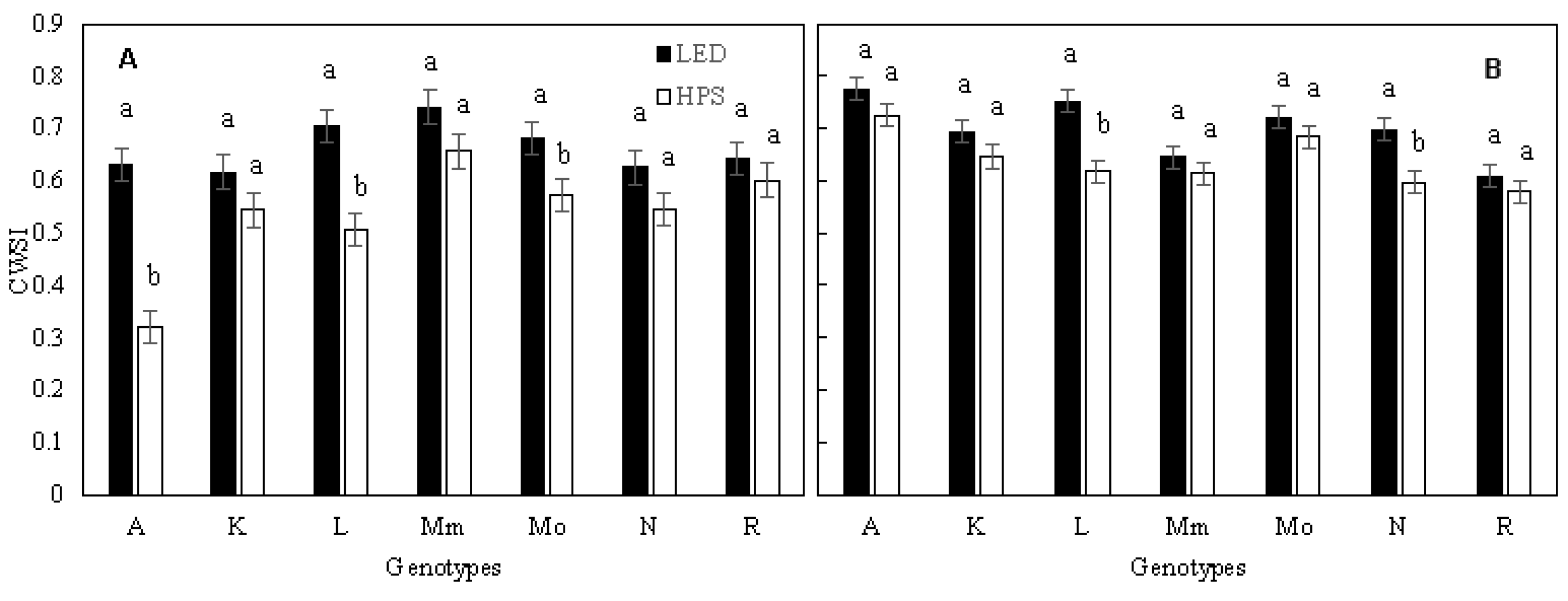
2.4. Plant Water Stress Indices
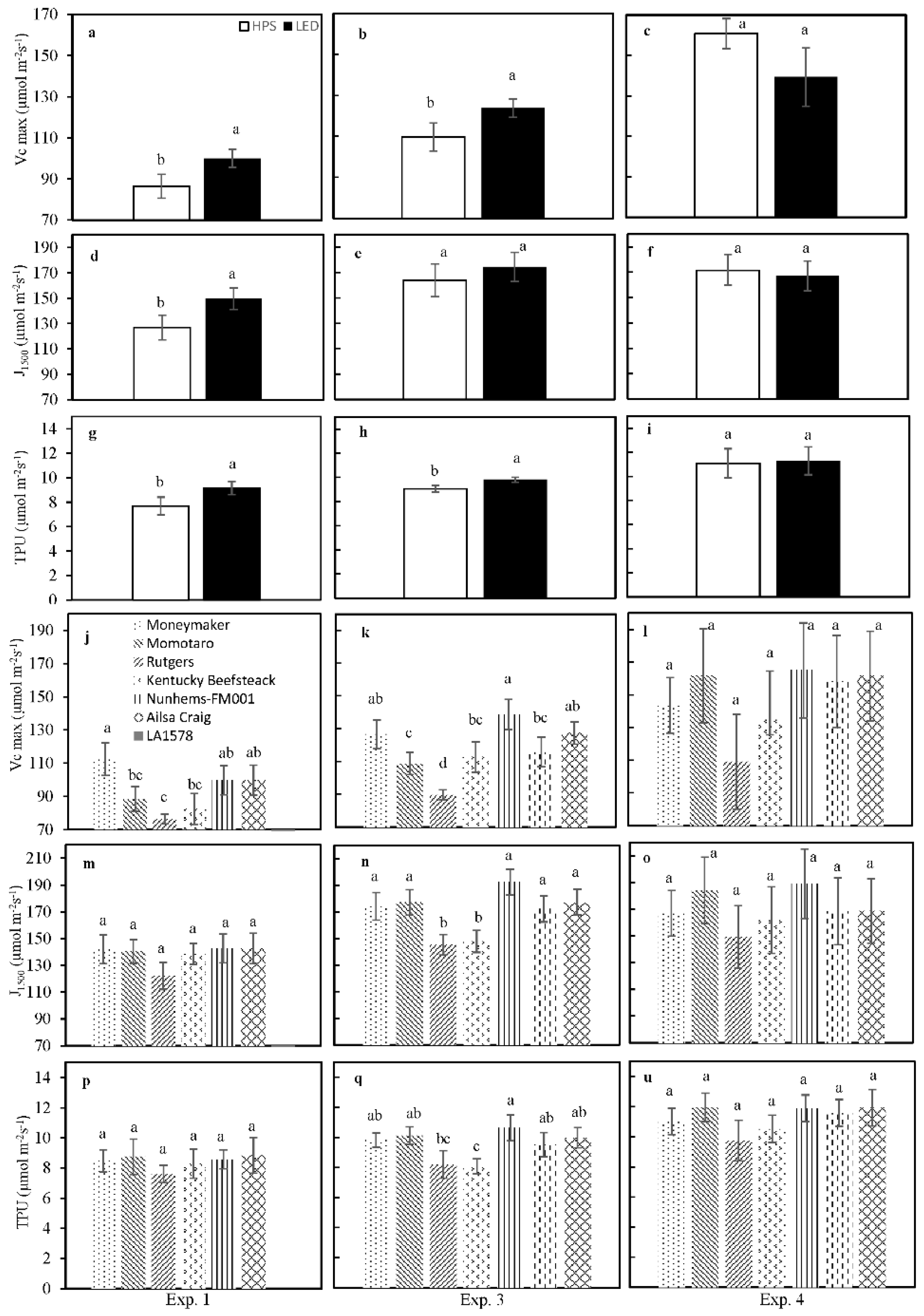
2.5. CO2 Response of Net Photosynthesis Rate
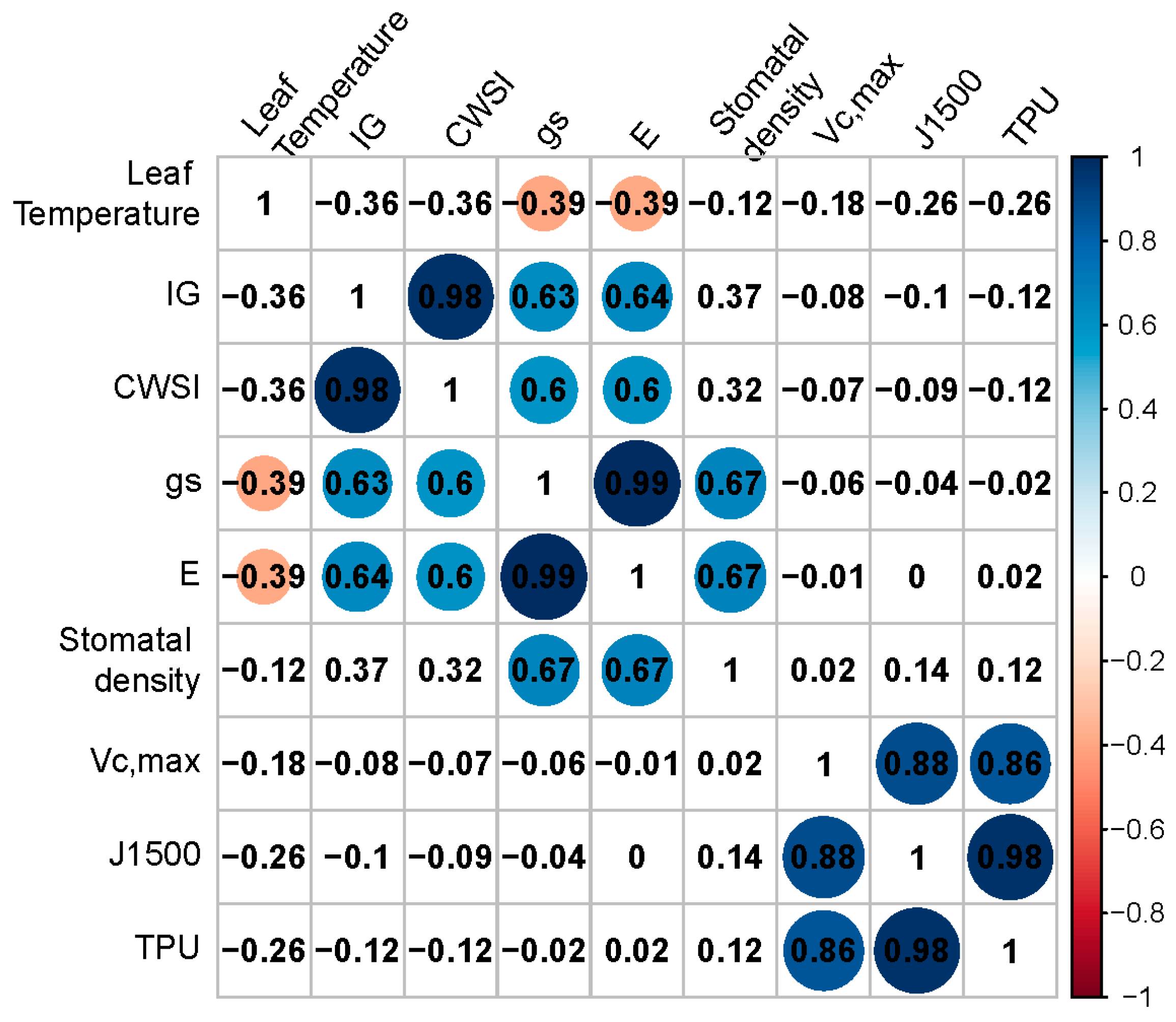
2.6. Correlations between Physiological Parameters
3. Discussion
3.1. High Pressure Sodium (HPS) and LED Supplemental Light Influenced Leaf Temperature
3.2. The Effects of Supplemental Light Spectral Quality on Plant Physiology
3.3. Plant Water Stress Indices for Phenotyping of Transpiration
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Daily Light Integral (DLI) and Light Quality
4.3. Stomatal Conductance (gs) and Transpiration Rate (E)
4.4. Photosynthetic CO2 Response Curves
4.5. Stomatal Density
4.6. Leaf Temperature
4.7. Plant Water Stress Indices
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Gómez, C.; Mitchell, C.A. Supplemental lighting for greenhouse-grown tomatoes: Intracanopy LED towers vs overhead HPS lamps. Acta Hortic. 2014, 1037, 855–862. [Google Scholar] [CrossRef]
- Wei, H.; Zhao, J.; Hu, J.; Jeong, B.R. Effect of supplementary light intensity on quality of grafted tomato seedlings and expression of two photosynthetic genes and proteins. Agronomy 2019, 9, 339. [Google Scholar] [CrossRef] [Green Version]
- Toida, H.; Ohyama, K.; Omura, Y.; Kozai, T. Enhancement of growth and development of tomato seedlings by extending the light period each day. HortScience 2005, 40, 370–373. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.M.; Hwang, S.J. The growth and development of ‘mini chal’ tomato plug seedlings grown under various wavelengths using light emitting diodes. Agronomy 2019, 9, 157. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Lin, M.; Mitchell, C.A. Light spectral and thermal properties govern biomass allocation in tomato through morphological and physiological changes. Environ. Exp. Bot. 2019, 157, 228–240. [Google Scholar] [CrossRef]
- Särkkä, L.E.; Jokinen, K.; Ottosen, C.O.; Kaukoranta, T. Effects of HPS and LED lighting on cucumber leaf photosynthesis, light quality penetration and temperature in the canopy, plant morphology and yield. Agric. Food Sci. 2017, 26, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Morrow, R.C. LED lighting in horticulture. HortScience 2008, 43, 1947–1950. [Google Scholar] [CrossRef] [Green Version]
- Gilewski, M. The state of art in the horticulture lighting. Photonics Lett. Pol. 2020, 12, 100–102. [Google Scholar] [CrossRef]
- Nelson, J.A.; Bugbee, B. Analysis of environmental effects on leaf temperature under sunlight, high pressure sodium and light emitting diodes. PLoS ONE 2015, 10, e0138930. [Google Scholar] [CrossRef]
- Leong, T.; Anderson, J.A.N.M. Changes in composition and function of thylakoid membranes as a result of photosynthetic adaptation of chloroplast from pea plants grown under different light conditions. Biochim. Biophys. Acta. 1983, 723, 391–399. [Google Scholar] [CrossRef]
- Rascher, U.; Nedbal, L. Dynamics of photosynthesis in fluctuating light. Curr. Opin. Plant Biol. 2006, 9, 671–678. [Google Scholar] [CrossRef]
- Gratani, L. Plant phenotypic plasticity in response to environmental factors. Adv. Bot. 2014, 2014, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.Y.; Chang, T.T.; Guo, S.R.; Xu, Z.G.; Li, J. Effect of different light quality of LED on growth and photosynthetic character in cherry tomato seedling. Acta Hortic. 2011, 907, 325–330. [Google Scholar] [CrossRef]
- Wang, J.; Lu, W.; Tong, Y.; Yang, Q. Leaf morphology, photosynthetic performance, chlorophyll fluorescence, stomatal development of lettuce (Lactuca sativa L.) exposed to different ratios of red light to blue light. Front. Plant Sci. 2016, 7, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giuliani, R.; Koteyeva, N.; Voznesenskaya, E.; Evans, M.A.; Cousins, A.B.; Edwards, G.E. Coordination of leaf photosynthesis, transpiration, and structural traits in rice and wild relatives (Genus Oryza). Plant Physiol. 2013, 162, 1632–1651. [Google Scholar] [CrossRef] [Green Version]
- Fischer, R.A.; Rebetzke, G.J. Indirect selection for potential yield in early-generation, spaced plantings of wheat and other small-grain cereals: A review. Crop Pasture Sci. 2018, 69, 439–459. [Google Scholar] [CrossRef]
- Lanoue, J.; Leonardos, E.D.; Khosla, S.; Hao, X.; Grodzinski, B. Effect of elevated CO2 and spectral quality on whole plant gas exchange patterns in tomatoes. PLoS ONE 2018, 13, e0205861. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Xiaoxiao, W.; Min, P.; Xiaoying, L.; Lijun, G.; Zhigang, X. Effect different spectral LED on photosynthesis and distribution of photosynthate of cherry tomato seedlings. In Proceedings of the 14th China International Forum on Solid State Lighting: International Forum on Wide Bandgap Semiconductors China (SSLChina: IFWS), Beijing, China, 1–3 November 2017; pp. 78–84. [Google Scholar]
- Eisinger, W.R.; Bogomolni, R.A.; Taiz, L. Interactions between a blue-green reversible photoreceptor and a separate UV-B receptor in stomatal guard cells. Am. J. Bot. 2003, 90, 1560–1566. [Google Scholar] [CrossRef] [Green Version]
- Frechilla, S.; Talbott, L.D.; Bogomolni, R.A.; Zeiger, E. Reversal of blue light-stimulated stomatal opening by green light. Plant Cell Physiol. 2000, 41, 171–176. [Google Scholar] [CrossRef] [Green Version]
- Talbott, L.D.; Nikolova, G.; Ortiz, A.; Shmayevich, I.; Zeiger, E. Green light reversal of blue-light-stimulated stomatal opening is found in a diversity of plant species. Am. J. Bot. 2002, 89, 366–368. [Google Scholar] [CrossRef] [Green Version]
- Smith, H.L.; Mcausland, L.; Murchie, E.H. Don’t ignore the green light: Exploring diverse roles in plant processes. J. Exp. Bot. 2017, 68, 2099–2110. [Google Scholar] [CrossRef]
- Li, S.; Hamani, A.K.M.; Si, Z.; Liang, Y.; Gao, Y.; Duan, A. Leaf gas exchange of tomato depends on abscisic acid and jasmonic acid in response to neighboring plants under different soil nitrogen regimes. Plants 2020, 9, 1674. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, D.; Baret, F.; Welcker, C.; Bostrom, A.; Ball, J.; Cellini, F.; Lorence, A.; Chawade, A.; Khafif, M.; Noshita, K.; et al. What is cost-efficient phenotyping? Optimizing costs for different scenarios. Plant Sci. 2019, 282, 14–22. [Google Scholar] [CrossRef] [Green Version]
- Vialet-Chabrand, S.; Lawson, T. Dynamic leaf energy balance: Deriving stomatal conductance from thermal imaging in a dynamic environment. J. Exp. Bot. 2019, 70, 2839–2855. [Google Scholar] [CrossRef] [Green Version]
- Vialet-Chabrand, S.; Lawson, T. Thermography methods to assess stomatal behaviour in a dynamic environment. J. Exp. Bot. 2020, 71, 2329–2338. [Google Scholar] [CrossRef] [Green Version]
- Jones, H.G. Use of infrared thermography for monitoring stomatal closure in the field: Application to grapevine. J. Exp. Bot. 2002, 53, 2249–2260. [Google Scholar] [CrossRef]
- Mott, K.A.; Peak, D. Stomatal responses to humidity and temperature in darkness. Plant Cell Environ. 2010, 33, 1084–1090. [Google Scholar] [CrossRef] [Green Version]
- von Caemmerer, S.; Evans, J.R. Temperature responses of mesophyll conductance differ greatly between species. Plant Cell Environ. 2015, 38, 629–637. [Google Scholar] [CrossRef] [PubMed]
- Lahr, E.C.; Schade, G.W.; Crossett, C.C.; Watson, M.R. Photosynthesis and isoprene emission from trees along an urban-rural gradient in Texas. Glob. Chang. Biol. 2015, 21, 4221–4236. [Google Scholar] [CrossRef] [PubMed]
- Bañon, S.; Fernandez, J.A.; Franco, J.A.; Torrecillas, A. Effects of water stress and night temperature preconditioning on water relations and morphological and anatomical changes of Lotus creticus plants. Sci. Hortic. 2004, 101, 333–342. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Sharkey, T.D. Stomatal conductance and photosynthesis. Annu. Rev. Plant Physiol. 1982, 33, 317–345. [Google Scholar] [CrossRef]
- Xu, Z.; Zhou, G. Responses of leaf stomatal density to water status and its relationship with photosynthesis in a grass. J. Exp. Bot. 2008, 59, 3317–3325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sobejano-Paz, V.; Mikkelsen, T.N.; Baum, A.; Mo, X.; Liu, S.; Köppl, C.J.; Johnson, M.S.; Gulyas, L.; García, M. Hyperspectral and thermal sensing of stomatal conductance, transpiration, and photosynthesis for soybean and maize under drought. Remote Sens 2020, 12, 3182. [Google Scholar] [CrossRef]
- Kim, H.H.; Goins, G.D.; Wheeler, R.M.; Sager, J.C. Stomatal conductance of lettuce grown under or exposed to different light qualities. Ann. Bot. 2004, 94, 691–697. [Google Scholar] [CrossRef] [Green Version]
- Muneer, S.; Kim, E.J.; Park, J.S.; Lee, J.H. Influence of green, red and blue light emitting diodes on multiprotein complex proteins and photosynthetic activity under different light intensities in lettuce leaves (Lactuca sativa L.). Int. J. Mol. Sci. 2014, 15, 4657–4670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- XiaoYing, L.; ShiRong, G.; ZhiGang, X.; XueLei, J.; Tezuka, T. Regulation of chloroplast ultrastructure, cross-section anatomy of leaves, and morphology of stomata of cherry tomato by different light irradiations of light-emitting diodes. HortScience 2011, 46, 217–221. [Google Scholar] [CrossRef] [Green Version]
- Hogewoning, S.W.; Trouwborst, G.; Maljaars, H.; Poorter, H.; van Ieperen, W.; Harbinson, J. Blue light dose-responses of leaf photosynthesis, morphology, and chemical composition of Cucumis sativus grown under different combinations of red and blue light. J. Exp. Bot. 2010, 61, 3107–3117. [Google Scholar] [CrossRef]
- Trouwborst, G.; Hogewoning, S.W.; van Kooten, O.; Harbinson, J.; van Ieperen, W. Plasticity of photosynthesis after the ‘red light syndrome’ in cucumber. Environ. Exp. Bot. 2016, 121, 75–82. [Google Scholar] [CrossRef]
- Ji, Y.; Ouzounis, T.; Courbier, S.; Kaiser, E.; Nguyen, P.T.; Schouten, H.J.; Visser, R.G.F.; Pierik, R.; Marcelis, L.F.M.; Heuvelink, E. Far-red radiation increases dry mass partitioning to fruits but reduces Botrytis cinerea resistance in tomato. Environ. Exp. Bot. 2019, 168, 103889. [Google Scholar] [CrossRef]
- Kaiser, E.; Ouzounis, T.; Giday, H.; Schipper, R.; Heuvelink, E.; Marcelis, L.F.M. Adding blue to red supplemental light increases biomass and yield of greenhouse-grown tomatoes, but only to an optimum. Front. Plant Sci. 2019, 9, 2002. [Google Scholar] [CrossRef] [Green Version]
- Yamori, W.; Noguchi, K.; Hikosaka, K.; Terashima, I. Phenotypic plasticity in photosynthetic temperature acclimation among crop species with different cold tolerances. Plant Physiol. 2010, 152, 388–399. [Google Scholar] [CrossRef] [Green Version]
- Yamori, W.; Hikosaka, K.; Way, D.A. Temperature response of photosynthesis in C3, C4, and CAM plants: Temperature acclimation and temperature adaptation. Photosynth. Res. 2014, 119, 101–117. [Google Scholar] [CrossRef] [PubMed]
- Jones, H.G. Use of thermography for quantitative studies of spatial and temporal variation of stomatal conductance over leaf surfaces. Plant Cell Environ. 1999, 22, 1043–1055. [Google Scholar] [CrossRef] [Green Version]
- Idso, S.B. Non-water-stressed baselines: A key to measuring and interpreting plant water stress. Agric. Meteorol. 1982, 27, 59–70. [Google Scholar] [CrossRef]
- Jones, H.G. Application of thermal imaging and infrared sensing in plant physiology and ecophysiology. Adv. Bot. Res. 2004, 41, 107–163. [Google Scholar] [CrossRef]
- Fourty, T.; Baret, F.; Jacquemoud, S.; Schmuck, G.; Verdebout, J. Leaf optical properties with explicit description of its biochemical composition: Direct and inverse problems. Remote Sens. Environ. 1996, 56, 104–117. [Google Scholar] [CrossRef]
- Lanoue, J.; Leonardos, E.D.; Ma, X.; Grodzinski, B. The effect of spectral quality on daily patterns of gas exchange, biomass gain, and water-use-efficiency in tomatoes and Lisianthus: An assessment of whole plant measurements. Front. Plant Sci. 2017, 8, 1076. [Google Scholar] [CrossRef]
- McCree, K.J. The action spectrum, absorptance and quantum yield of photosynthesis in crop plants. Agric. Meteorol. 1972, 9, 191–216. [Google Scholar] [CrossRef]
- Long, S.P.; Bernacchi, C.J. Gas exchange measurements, what can they tell us about the underlying limitations to photosynthesis? Procedures and sources of error. J. Exp. Bot. 2003, 54, 2393–2401. [Google Scholar] [CrossRef] [Green Version]
- Sharkey, T.D. What gas exchange data can tell us about photosynthesis. Plant Cell Environ. 2016, 39, 1161–1163. [Google Scholar] [CrossRef]
- Berghuijs, H.N.C.; Yin, X.; Tri Ho, Q.; van der Putten, P.E.L.; Verboven, P.; Retta, M.A.; Nicolaï, B.M.; Struik, P.C. Modeling the relationship between CO2 assimilation and leaf anatomical properties in tomato leaves. Plant Sci. 2015, 238, 297–311. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.; Weyers, J.D.B.; Berry, W.G. Variation in stomatal characteristics over the lower surface of Commelina communis leaves. Plant Cell Environ. 1989, 12, 653–659. [Google Scholar] [CrossRef]
- de Mendiburu, F. Agricolae; Universidad Nacional Agraria La Molina: Lima, Peru, 2020. [Google Scholar]
- Graves, S.; Piepho, H.-P.; Selzer, L. Multcompview: Visualizations of Paired Comparisons. R Package Version 0.1-5. 2019. Available online: http://CRAN.Rproject.org/package=multcompView (accessed on 21 January 2021).
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage Publications: Thousand Oaks, CA, USA, 2014. [Google Scholar]
- Kassambara, A. Ggpubr: “Ggplot2” Based Publication Ready Plots. R Package Version 0.2. 2018. Available online: https://CRAN.R-project.org/package=ggpubr (accessed on 21 January 2021).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stomatal Conductance (mol H2O m−2 s−1) | ||||
|---|---|---|---|---|
| Experiment | 1 | 2 | 3 | 4 |
| Treatments (SL) | ||||
| HPS | 0.32 b | 0.37 b | 0.59 b | 0.56 b |
| LED | 0.35 a | 0.40 a | 0.66 a | 0.60 a |
| Genotypes (Gn) | ||||
| Moneymaker | 0.36 a | 0.39 ab | 0.62 b | 0.56 cd |
| Momotaro | 0.36 a | 0.41 a | 0.61 b | 0.54 d |
| LA1578 | - | 0.36 b | 0.73 a | 0.66 a |
| Rutgers | 0.31 b | 0.36 b | 0.56 c | 0.50 e |
| Kentucky Beefsteak | 0.29 b | 0.38 ab | 0.63 b | 0.62 b |
| Nunhems-FM001 | 0.35 a | 0.41 a | 0.60 bc | 0.62 b |
| Ailsa Craig | 0.32 b | 0.40 a | 0.61 b | 0.58 c |
| Significance 1 | ||||
| SL | * | * | ** | *** |
| Gn | *** | * | *** | *** |
| SL × Gn | * | *** | NS | ** |
| Stomatal Density (Number mm−2) | ||
|---|---|---|
| Experiment | 1 | 4 |
| Treatment (SL) | ||
| HPS | 185 ± 9 b | 186 ± 7 b |
| LED | 216 ± 11 a | 205 ± 8 a |
| Genotype (Gn) | ||
| Moneymaker | 192 ± 13 bcd | 176 ± 6 c |
| Momotaro | 195 ± 13 bc | 188 ± 13 bc |
| LA1578 | 276 ± 14 a | 347 ± 7 a |
| Rutgers | 168 ± 8 cd | 135 ± 8 d |
| Kentucky Beefsteak | 214 ± 11 bc | 193 ± 7 bc |
| Nunhems-FM001 | 216 ± 11 b | 210 ± 10 b |
| Ailsa Craig | 95 ± 11 d | 136 ± 7 d |
| Significance 1 | ||
| SL | *** | ** |
| Gn | *** | *** |
| SL × Gn | NS | NS |
| Leaf Temperature (°C) | ||||
|---|---|---|---|---|
| Experiment | 1 | 2 | 3 | 4 |
| Treatment (SL) | ||||
| HPS | 23.8 ± 1.4 a | 21.9 ± 1.0 a | 22.0 ± 1.5 a | 23.4 ± 0.9 a |
| LED | 22.0 ± 1.2 b | 21.1 ± 0.7 b | 22.4 ± 1.0 a | 23.1 ± 1.2 a |
| Significance 1 | ||||
| SL | *** | *** | NS | NS |
| Crop Water Stress Index (CWSI) | ||||
|---|---|---|---|---|
| Experiment | 1 | 2 | 3 | 4 |
| Treatments (SL) | ||||
| HPS | 0.53 b | 0.54 b | 0.65 b | 0.68 a |
| LED | 0.58 a | 0.66 a | 0.69 a | 0.69 a |
| Genotypes (Gn) | ||||
| Moneymaker | 0.63 a | 0.70 a | 0.63 de | 0.70 b |
| Momotaro | 0.56 b | 0.63 b | 0.70 b | 0.64 de |
| LA1578 | - | 0.62 b | 0.69 bc | 0.76 a |
| Rutgers | 0.58 ab | 0.61 b | 0.59 e | 0.61 e |
| Kentucky Beefsteak | 0.46 c | 0.59 b | 0.67 bcd | 0.67 bc |
| Nunhems-FM001 | 0.50 c | 0.58 b | 0.65 cd | 0.66 cd |
| Ailsa Craig | 0.59 ab | 0.48 c | 0.75 a | 0.75 a |
| Significance 1 | ||||
| SL | ** | *** | ** | NS |
| Gn | *** | *** | *** | *** |
| SL × Gn | NS | *** | *** | NS |
| Light Quality Parameters | Supplemental Light | |
|---|---|---|
| LED | HPS | |
| % Blue (400–500 nm) | 5 | 5 |
| % Green–Yellow (501–600 nm) | 0 | 50 |
| % Red (601–700 nm) | 95 | 40 |
| % Far–Red (701–800 nm) | 0 | 5 |
| (Green–Yellow):Blue | 0 | 10 |
| Red:Far Red | NA | 965 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palmitessa, O.D.; Prinzenberg, A.E.; Kaiser, E.; Heuvelink, E. LED and HPS Supplementary Light Differentially Affect Gas Exchange in Tomato Leaves. Plants 2021, 10, 810. https://doi.org/10.3390/plants10040810
Palmitessa OD, Prinzenberg AE, Kaiser E, Heuvelink E. LED and HPS Supplementary Light Differentially Affect Gas Exchange in Tomato Leaves. Plants. 2021; 10(4):810. https://doi.org/10.3390/plants10040810
Chicago/Turabian StylePalmitessa, Onofrio Davide, Aina E. Prinzenberg, Elias Kaiser, and Ep Heuvelink. 2021. "LED and HPS Supplementary Light Differentially Affect Gas Exchange in Tomato Leaves" Plants 10, no. 4: 810. https://doi.org/10.3390/plants10040810
APA StylePalmitessa, O. D., Prinzenberg, A. E., Kaiser, E., & Heuvelink, E. (2021). LED and HPS Supplementary Light Differentially Affect Gas Exchange in Tomato Leaves. Plants, 10(4), 810. https://doi.org/10.3390/plants10040810