
Transcriptional Changes of Cell Wall Organization Genes and Soluble Carbohydrate Alteration during Leaf Blade Development of Rice Seedlings


Abstract
:1. Introduction
2. Results
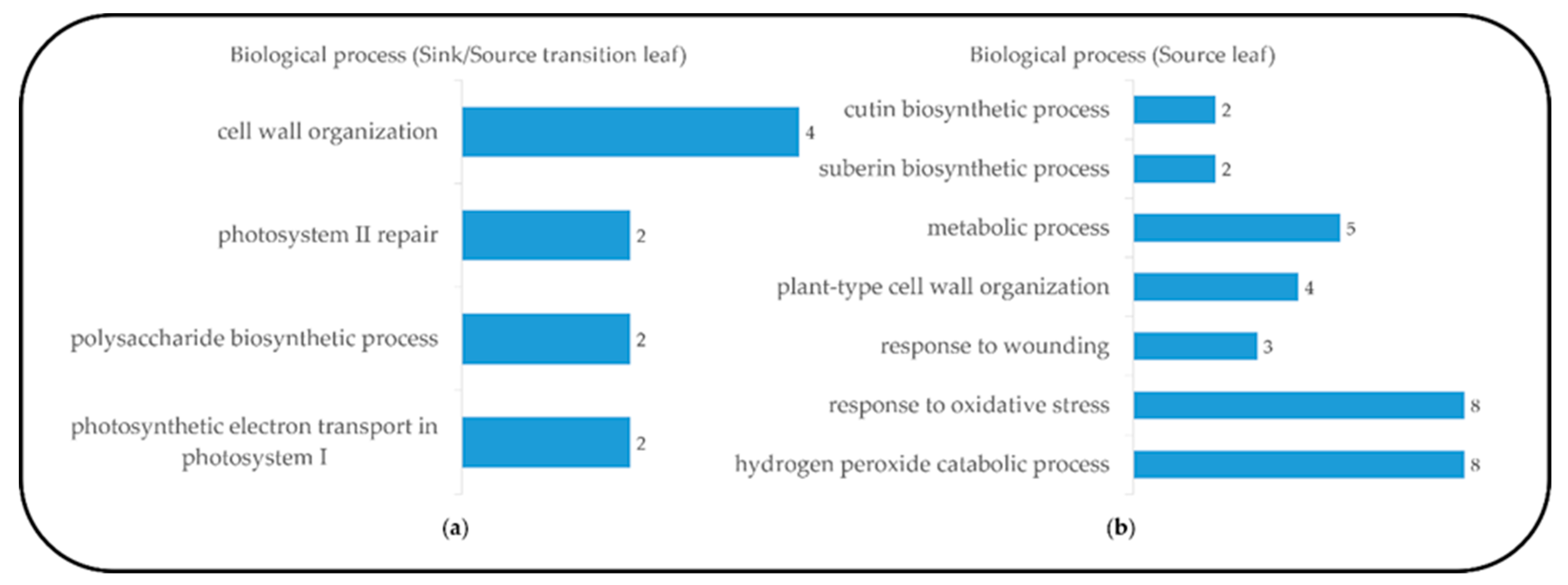
2.1. Leaf Blade Transcriptome and DEGs Analysis
2.2. Validation of RNA-seq Data by qRT-PCR Analysis
2.3. Primary and Secondary Cell Wall Organization and Soluble Carbohydrates Compositions during Leaf Development
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Sampling
4.2. RNA Extraction, cDNA Synthesis and qRT-PCR
4.3. RNA-Seq Library Construction, Sequencing and DEGs Analysis
4.4. Soluble Carbohydrates Analysis
4.5. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reiter, W.D. Biosynthesis and properties of the plant cell wall. Curr. Opin. Plant Biol. 2002, 5, 536–542. [Google Scholar] [CrossRef]
- Scheller, Z.A.; Ulvskov, P. Hemicelluloses. Annu. Rev. Plant Biol. 2010, 61, 263–289. [Google Scholar] [CrossRef]
- McCann, M.C.; Knox, J.P. Plant cell wall biology: Polysaccharides in architectural and developmental contexts. Ann. Plant Rew. Online 2018, 41, 343–366. [Google Scholar]
- Zhong, R.; Cui, D.; Ye, Z.H. Secondary cell wall biosynthesis. New Phytol. 2019, 221, 1703–1723. [Google Scholar] [CrossRef] [Green Version]
- Brégrad, A.; Allard, G. Sink to source transition in developing leaf blades of tall fescue. New Phytol. 1999, 141, 45–50. [Google Scholar] [CrossRef]
- Schnyder, H.; Seo, S.; Rademacher, I.F.; Kühbauch, W. Spatial distribution of growth rates and of epidermal cell lengths in the elongation zone during leaf development in Lolium perenne L. Planta 1990, 181, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Turgeon, R. The sink±source transition in leaves. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1989, 40, 119–138. [Google Scholar] [CrossRef]
- Krapp, A.; Quick, W.P.; Stitt, M. Ribulose-1,5-bisphosphate carboxylase-oxygenase, other Calvin-cycle enzymes and chlorophyll decrease when glucose is supplied to mature spinach leaves via the transpiration stream. Planta. 1991, 186, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Araya, T.; Noguchi, K.; Terashima, I. Effects of Carbohydrate Accumulation on Photosynthesis Differ between Sink and Source Leaves of Phaseolus vulgaris L. Plant Cell Physiol. 2006, 47, 644–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.L.; Jiang, H.; Chen, H.S.; Tsai, C.J.; Harding, S.A. Metabolic Profiling of the Sink-to-Source Transition in Developing Leaves of Quaking Aspen. Plant Physiol. 2004, 136, 3364–3375. [Google Scholar]
- Yun, H.; Lim, S.; Kim, Y.X.; Lee, Y.; Lee, S.; Lee, D.; Park, K.; Sung, J. Diurnal changes in C-N metabolism and response of rice seedlings to UV-B radiation. J. Plant Physiol. 2018, 228, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Hara, Y.; Yokoyama, R.; Osakabe, K.; Toki, S.; Nishitani, K. Function of xyloglucan endotransglucosylase/hydrolases in rice. Ann. Bot. 2014, 114, 1309–1318. [Google Scholar] [CrossRef] [Green Version]
- Hazen, S.; Scott-Craig, J.; Walton, J. Cellulose synthase like genes of rice. Plant Physiol. 2002, 128, 336–340. [Google Scholar] [CrossRef] [PubMed]
- Oraby, H.; Venkatesh, B.; Dale, B.; Ahmad, R.; Ransom, C.; Oehmke, J.; Sticklen, M. Enhanced conversion of plant biomass into glucose using transgenic rice-produced endoglucanase for cellulosic ethanol. Transgenic Res. 2007, 16, 739–749. [Google Scholar] [CrossRef] [PubMed]
- Carpita, N.C. The structure and biogenesis of the cell walls of grasses. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1996, 47, 445–476. [Google Scholar] [CrossRef]
- Kim, S.J.; Chandrasekar, B.; Rea, A.C.; Danhof, L.; Zemelis-Durfee, S.; Thrower, N.; Shepard, Z.S.; Pauly, M.; Brandizzi, F.; Keegstra, K. The synthesis of xyloglucan, an abundant plant cell wall polysaccharide, requires CSLC function. Proc. Natl. Acad. Sci. USA 2020, 117, 20316–20324. [Google Scholar] [CrossRef]
- Hsieh, Y.S.; Harris, P.J. Xyloglucans of monocotyledons have diverse structures. Mol. Plant 2009, 2, 943–965. [Google Scholar] [CrossRef]
- Lane, D.R.; Wiedemeier, A.; Peng, L.; Hofte, H.; Vernhettes, S.; Desprez, T.; Hocart, C.H.; Birch, R.J.; Baskin, T.I.; Burn, J.E.; et al. Temperature-sensitive alleles of RSW2 link the KORRIGAN endo-1,4-beta-glucanase to cellulose synthesis and cytokinesis in Arabidopsis. Plant Physiol. 2001, 126, 278–288. [Google Scholar] [CrossRef] [Green Version]
- Yokoyama, R. A genomic perspective on the evolutionary diversity of the plant cell wall. Plants 2020, 9, 1195. [Google Scholar] [CrossRef]
- Dong, L.; Qin, L.; Dai, X.; Ding, Z.; Bi, R.; Liu, P.; Chen, Y.; Brutnell, T.P.; Wang, X.; Li, P. Transcriptomic analysis of leaf sheath maturation in maize. Int. J. Mol. Sci. 2019, 20, 2472. [Google Scholar] [CrossRef] [Green Version]
- Nelson, T. The grass leaf developmental gradient as a platform for a systems understanding of the anatomical specialization of C4 leaves. J. Exp. Bot. 2011, 62, 3039–3048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hrmova, M.; Farkaš, V.; Lahnstein, J.; Fincher, G.B. A barley xyloglucan xyloglucosyl transferase covalently links xyloglucan, cellusosic substrates, and (1,3;1,4)-β-D-glucans. J. Biol. Chem. 2007, 283, 27344. [Google Scholar] [CrossRef]
- Franková, L.; Fry, S.C. Biochemistry and physiological roles of enzymes that ‘cut and paste’ plant cell-wall polysaccharides. J. Exp. Bot. 2013, 64, 3519–3550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van-Sandt, V.S.T.; Sullov, D.; Verbelen, J.P.; Vissenberg, K. Xyloglucan endotransglucosylase activity loosens a plant cell wall. Ann. Bot. 2007, 100, 1467–1473. [Google Scholar] [CrossRef] [Green Version]
- Vioque, J.; Kolattukudy, P.E. Resolution and purification of an aldehyde-generating and an alcohol-generating fatty acyl-CoA reductase from pea leaves (Pisum sativum L.). Arch. Biochem. Biophys. 1997, 340, 64–72. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, M.; Sun, Y.; Wang, Y.; Li, T.; Chai, G.; Jiang, W.; Shan, L.; Li, C.; Xiao, E.; et al. FAR5, a fatty acyl-coenzyme A reductase, is involved in primary alcohol biosynthesis of the leaf blade cuticular wax in wheat (Triticum aestivum L.). J. Exp. Bot. 2015, 66, 1165–1178. [Google Scholar] [CrossRef] [Green Version]
- Beisson, F.; Li-Beisson, Y.; Pollard, M. Solving the puzzles of cutin and suberin polymer biosynthesis. Curr. Opi. Plant Biol. 2012, 15, 329–337. [Google Scholar] [CrossRef]
- Li, Y.H.; Beisson, F.; Koo, A.J.K.; Molina, I.; Pollard, M.; Ohlrogge, J. Identification of acyltransferases required for cutin biosynthesis and production of cutin with suberin-like monomers. Proc. Natl. Acad. Sci. USA 2007, 104, 18339–18344. [Google Scholar] [CrossRef] [Green Version]
- Bontpart, T.; Cheynier, V.; Ageorges, A.; Terrier, N. BAHD or SCPL acyltransferase? What a dilemma for aclation in the world of plant phenolic compounds. New Phytol. 2015, 208, 695–707. [Google Scholar] [CrossRef]
- Bartley, L.E.; Peck, M.L.; Kim, S.R.; Ebert, B.; Manisseri, C.; Chiniquy, D.M.; Sykes, R.; Gao, L.; Rautengarten, C.; Vega-Sanchez, M.E.; et al. Overexpression of a BAHD acyltransferase, OsAt10, alters rice cell wall hydroxycinnamic acid content and saccharification. Plant Physiol. 2013, 161, 1615–1633. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.X.; Komatsuda, T.; Ma, J.F.; Nawrath, C.; Pourkheirandish, M.; Tagiri, A.; Hu, Y.G.; Sameri, M.; Li, X.R.; Zhao, X.; et al. An ATP-binding cassette subfamily G full transporter is essential for retention of leaf water in both wild barley and rice. Proc. Natl. Acad. Sci. USA 2011, 108, 12354–12359. [Google Scholar] [CrossRef] [Green Version]
- Anderson, L.S.; Dale, J.E. The sources of carbon for developing leaves of barley. J. Exp. Bot. 1983, 34, 405–414. [Google Scholar] [CrossRef]
- Wang, L.; Czedik-Eysenberg, A.; Mertz, R.A.; Si, Y.; Tohge, T.; Nunes-Nesi, A.; Arrivault, S.; Dedow, L.K.; Bryant, D.W.; Zhou, W.; et al. Comparative analyses of C4 and C3 photosynthesis in developing leaves of maize and rice. Nat. Biotech. 2014, 32, 1158–1168. [Google Scholar] [CrossRef]
- Verbancic, J.; Lunn, J.E.; Stitt, M.; Persson, S. Carbon supply and the regulation of cell wall synthesis. Mol. Plant 2018, 11, 75–94. [Google Scholar] [CrossRef] [PubMed]
- Shim, D.H.; Park, S.G.; Kim, K.M.; Bae, W.S.; Lee, G.W.; Ha, B.S.; Ro, H.S.; Kim, M.K.; Ryoo, R.; Rhee, S.K.; et al. Whole genome de novo sequencing and genome annotation of the world popular cultivated edible mushroom, Lentinula edodes. J. Biotechnol. 2016, 223, 24–25. [Google Scholar] [CrossRef] [PubMed]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef] [Green Version]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.B.; Park, S.Y.; Park, C.H.; Park, W.T.; Kim, S.J.; Ha, S.H.; Arasu, M.V.; Al-Dhabi, N.A.; Kim, J.K.; Park, S.U. Metabolomics of differently colored Gladiolus cultivars. Appl. Biol. Chem. 2016, 59, 597–607. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GO Term | Gene ID | Gene | Short Description | Log2 FC | p-Value | Relative Upregulation |
|---|---|---|---|---|---|---|
| Cell wall organization | Os05g0123100 | GT 43 | Xylan synthesis | 4.1 | 0.021 | 5.0 ± 0.6 *** |
| Os10g0577500 | XTH | Xyloglucan cleavage | 6.0 | 0.035 | 5.3 ± 0.5 ** | |
| Os06g0247900 | Endoglucanase 16 | Xyloglucan cleavage | 4.7 | 0.008 | 1.6 ± 1.1 * | |
| Os09g0428000 | CslC | Xyloglucan synthesis | 3.9 | 0.039 | 1.7 ± 1.4 | |
| Polysaccharide biosynthetic process | Os03g0678800 | GT 8 | Xylan biosynthesis | 9.1 | 0.002 | 4.1 ± 1.3 *** |
| Os04g0530900 | GT 8 | Xylan biosynthesis | 4.3 | 0.037 | 2.8 ± 0.9 ** |
| GO Term | Gene ID | Gene Name | Description | Log2 FC | p-Value | Relative Upregulation |
|---|---|---|---|---|---|---|
| Suberin and cutin biosynthesis | Os09g0567500 | FAR | Fatty acyl reduction | 16.7 | 0.022 | 1.4 ± 0.8 * |
| Os03g0281900 | ABC transporter | Wax/Cutin transport | 19.7 | 0.048 | 4.1 ± 0.3 * | |
| Os11g0679700 | GPAT 3 | Suberin/Cutin fatty alcohol transfer | 6.9 | 0.022 | 1.9 ± 0.5 * | |
| Os08g0562500 | BAHD acyltransferase | BAHD acyl transfer | 36.8 | 0.019 | 1.2 ± 0.9 * | |
| H2O2 catabolism | Os02g0236600 | POD | H2O2 peroxidation | 47.2 | 0.049 | 3.1 ± 0.0 *** |
| Os05g0499300 | POD | H2O2 peroxidation | 24.6 | 0.037 | 1.5 ± 0.2 | |
| Os07g0677100 | POD | H2O2 peroxidation | 24.1 | 0.037 | 2.4 ± 0.0 * |
| Stage | Glucose | Fructose | Galactose | Sucrose | Raffinose |
|---|---|---|---|---|---|
| Sink | 42.8 ± 1.7 | 13.3 ± 0.4 | 0.3 ± 0.1 | 180.0 ± 23.3 | 0.9 ± 0.1 |
| Sink-to-source | 15.6 ± 1.7 | 5.1 ± 0.7 | 0.1 ± 0.0 | 211.3 ± 57.8 | 0.9 ± 0.4 |
| Source | 26.0 ± 1.0 | 7.9 ± 0.4 | 0.4 ± 0.1 | 335.8 ± 66.0 | 2.8 ± 0.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Joo, J.-Y.; Kim, M.-S.; Sung, J. Transcriptional Changes of Cell Wall Organization Genes and Soluble Carbohydrate Alteration during Leaf Blade Development of Rice Seedlings. Plants 2021, 10, 823. https://doi.org/10.3390/plants10050823
Joo J-Y, Kim M-S, Sung J. Transcriptional Changes of Cell Wall Organization Genes and Soluble Carbohydrate Alteration during Leaf Blade Development of Rice Seedlings. Plants. 2021; 10(5):823. https://doi.org/10.3390/plants10050823
Chicago/Turabian StyleJoo, Jae-Yeon, Me-Sun Kim, and Jwakyung Sung. 2021. "Transcriptional Changes of Cell Wall Organization Genes and Soluble Carbohydrate Alteration during Leaf Blade Development of Rice Seedlings" Plants 10, no. 5: 823. https://doi.org/10.3390/plants10050823
APA StyleJoo, J. -Y., Kim, M. -S., & Sung, J. (2021). Transcriptional Changes of Cell Wall Organization Genes and Soluble Carbohydrate Alteration during Leaf Blade Development of Rice Seedlings. Plants, 10(5), 823. https://doi.org/10.3390/plants10050823