
Interactive Impacts of Beneficial Microbes and Si-Zn Nanocomposite on Growth and Productivity of Soybean Subjected to Water Deficit under Salt-Affected Soil Conditions

 ,
,  ,
,  , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Results
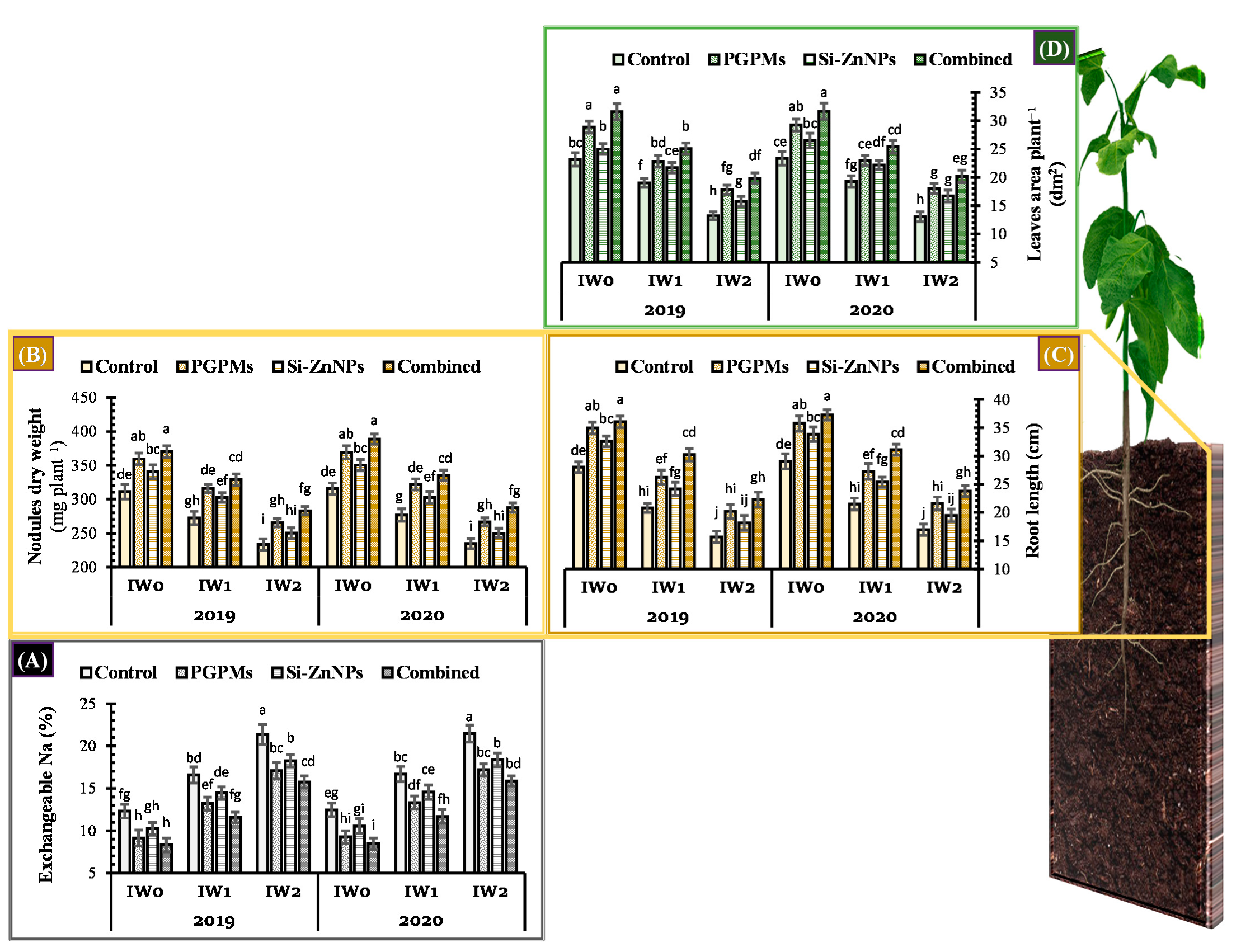
2.1. Exchangeable Sodium Percentage (ESP)
2.2. Vegetative Growth Characteristics
2.2.1. Root Length and Nodules Dry Weight
2.2.2. Total Leaves Area
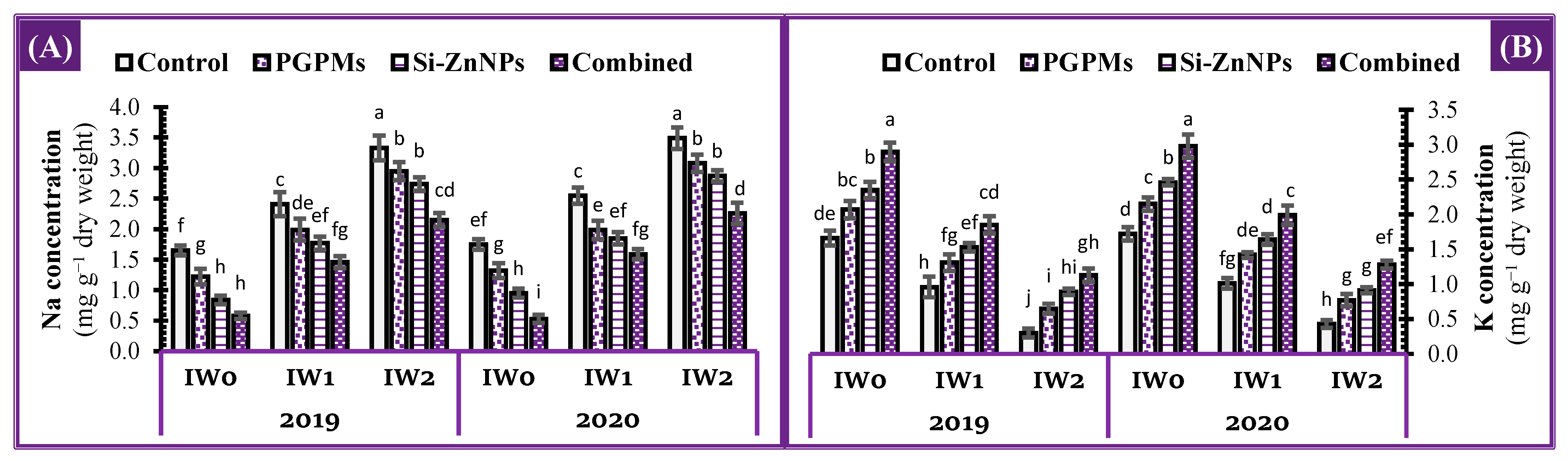
2.3. Concentration of Sodium and Potassium in Leaves
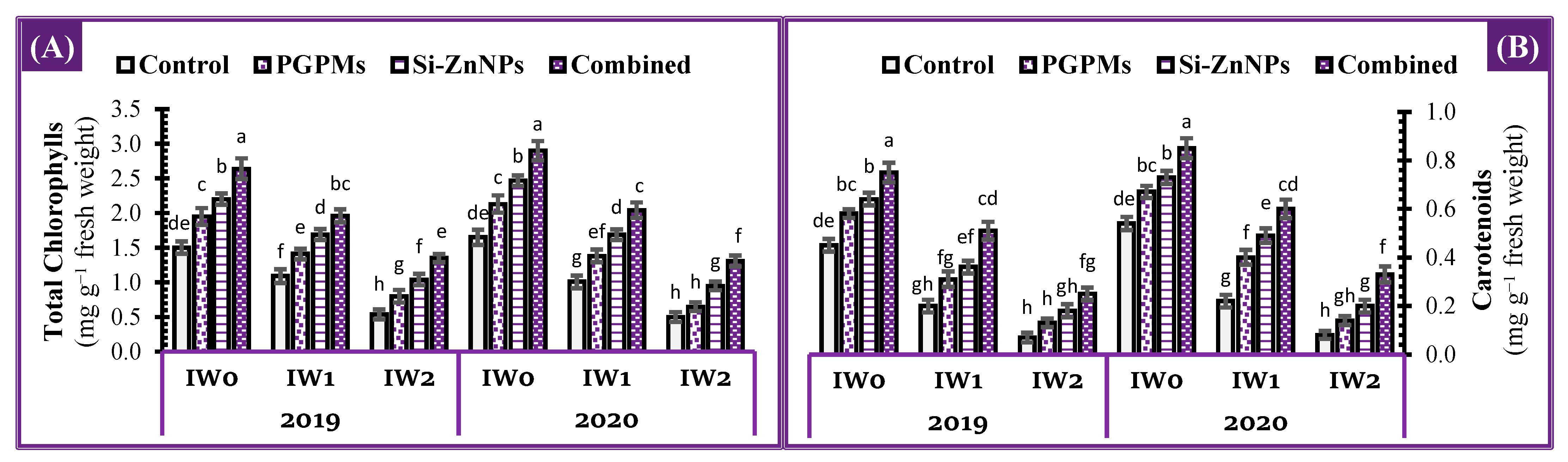
2.4. Photosynthetic Pigments Concentration
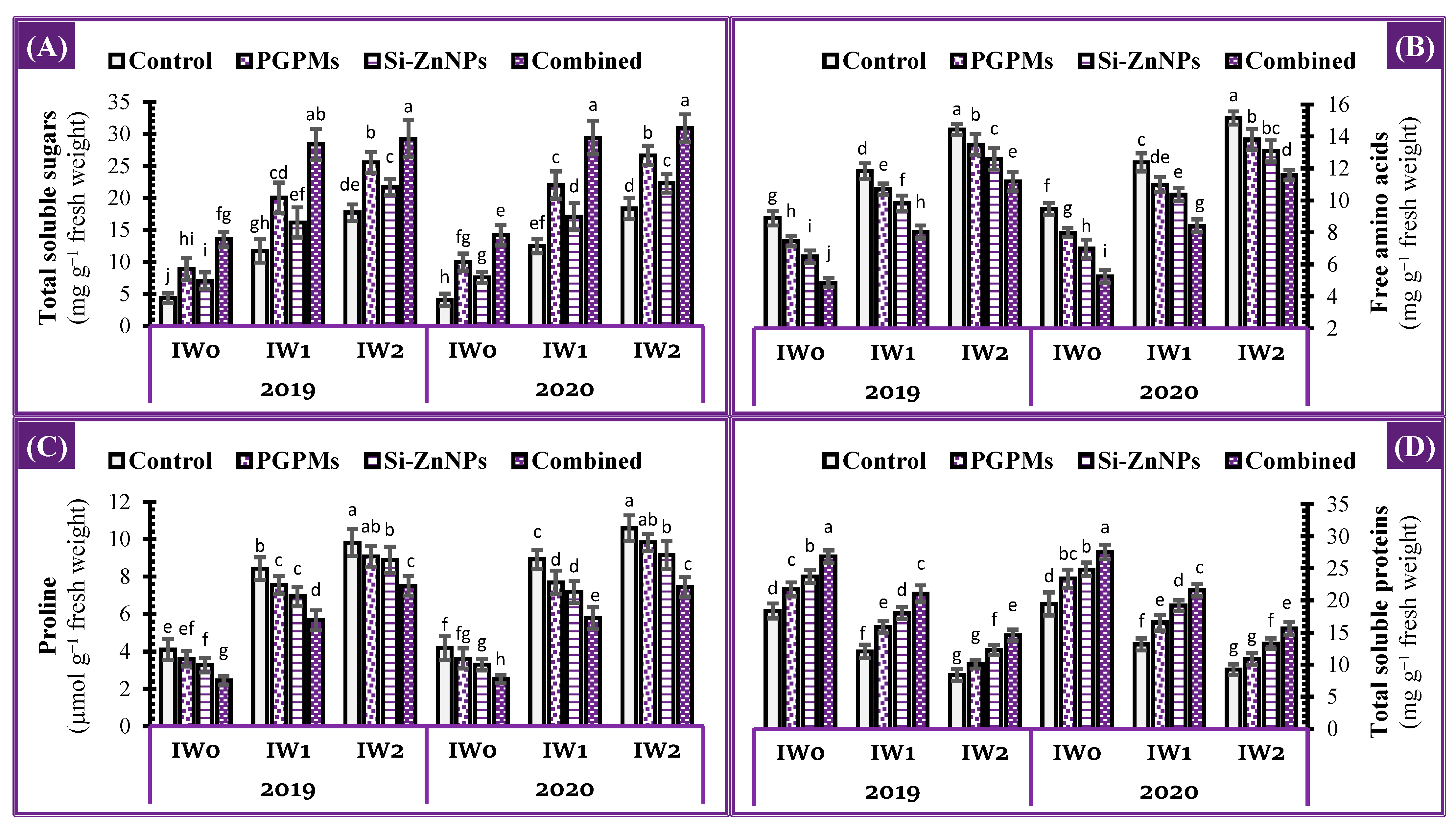
2.5. Osmoprotectants
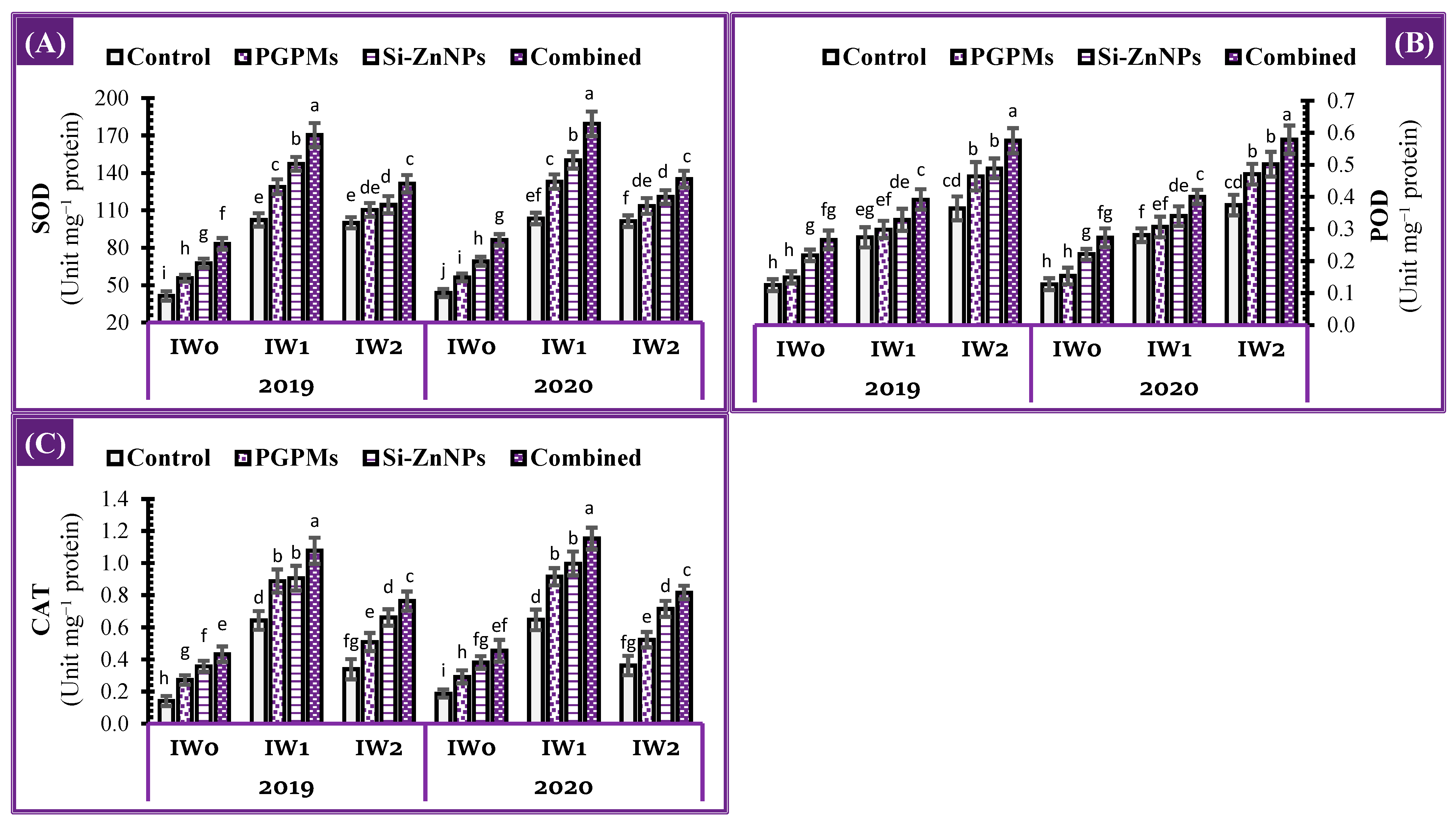
2.6. Activity of Antioxidant Enzymes
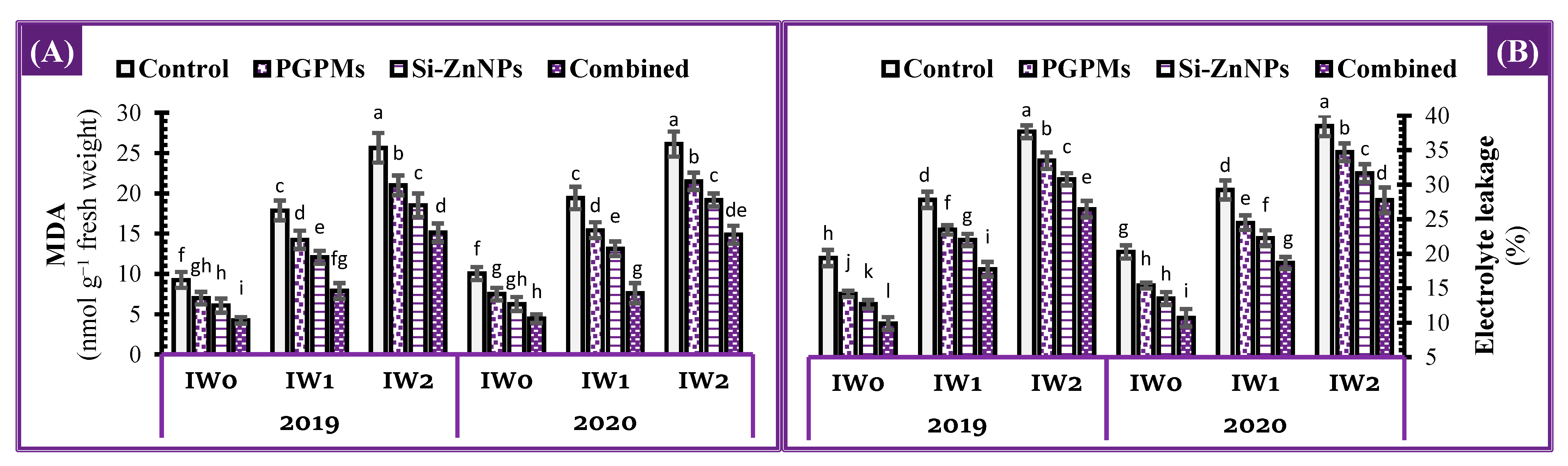
2.7. Oxidative Damage Status
2.7.1. Lipid Peroxidation
2.7.2. Electrolytes Leakage Percentage
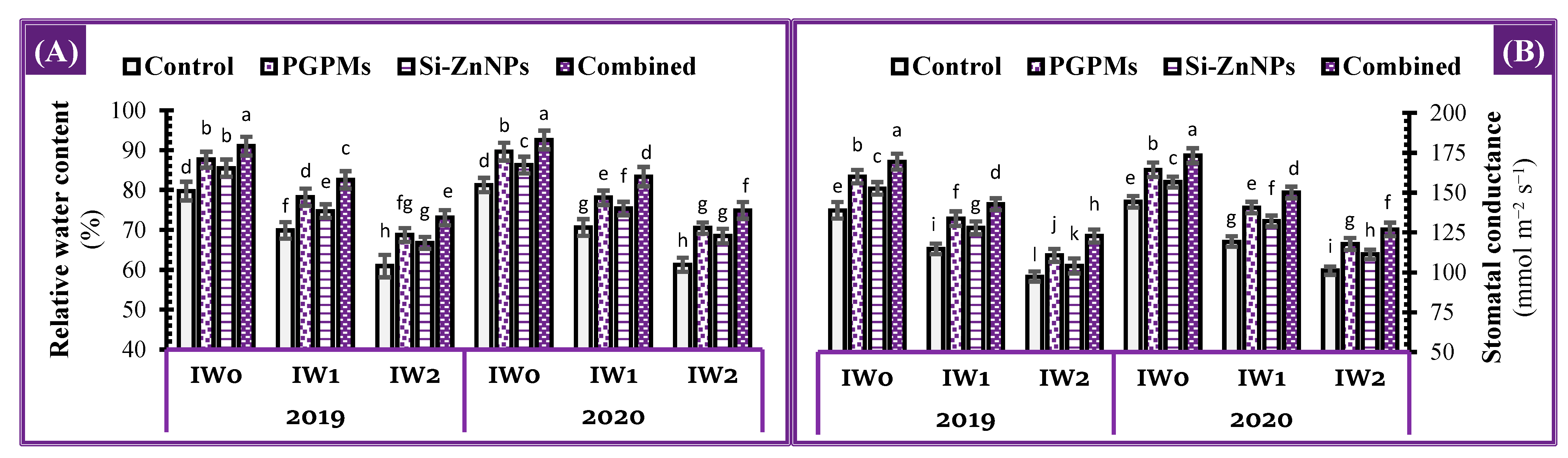
2.8. Plant Water Relations
2.9. Yield Related Traits and Seed Quality
3. Discussion
3.1. Yield Physiology under Well Irrigation
3.1.1. Rhizosphere-Shoot Connections
3.1.2. Physio-Biochemical Changes and Yield Relationship
3.2. Yield Physiology under Deficit Irrigation
4. Materials and Methods
4.1. Experimental Layout
4.2. Soil Physicochemical Analysis
4.3. Treatments Preparation and Application
4.3.1. Seed Inoculation with Microbial Consortium
4.3.2. Beneficial and Micro-Elements Nanoparticles
4.4. Traits Measurements
4.4.1. Exchangeable Sodium % (ESP)
4.4.2. Root Length, Nodules Dry Weight and Leaves Area
4.4.3. Potassium and Sodium Concentration
4.4.4. Total Chlorophylls and Carotenoids Concentration
4.4.5. Osmo-protectants and Protein Concentrations
Total Soluble Sugars (TSS)
Proline
Free Amino Acid (FAA)
Total Soluble Protein (TSP)
4.4.6. Activity of Antioxidant Enzymes (CAT, POD and SOD)
4.4.7. Oxidative Damage Status
Malondialdehyde (MDA)
Electrolyte Leakage Percentage (EL%)
4.4.8. Plant Water Relations
Leaf Relative Water Content (LRWC)
Stomatal Conductance
4.4.9. Seed Yield Related Traits and Nutritional Quality
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Munns, R.; Gilliham, M. Salinity tolerance of crops—What is the cost? New Phytol. 2015, 208, 668–673. [Google Scholar] [CrossRef] [Green Version]
- Negrão, S.; Schmöckel, S.M.; Tester, M. Evaluating physiological responses of plants to salinity stress. Ann. Bot 2017, 119, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parihar, P.; Singh, S.; Singh, R.; Singh, V.P.; Prasad, S.M. Effect of salinity stress on plants and its tolerance strategies: A review. Environ. Sci. Pollut. Res. 2014, 22, 4056–4075. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant. Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Hafez, E.M.; Kheir, A.M.S.; Badawy, S.A.; Rashwan, E.; Farig, M.; Osman, H.S. Differences in physiological and biochemical attributes of wheat in response to single and combined salicylic acid and biochar subjected to limited water irrigation in saline sodic soil. Plants 2020, 9, 1346. [Google Scholar] [CrossRef] [PubMed]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxic. Environ. Safety 2005, 60, 324–349. [Google Scholar] [CrossRef]
- Salim, B.B.M.; Hikal, M.S.; Osman, H.S. Ameliorating the deleterious effects of saline water on the antioxidants defense system and yield of eggplant using foliar application of zinc sulphate. Ann. Agric. Sci. 2019, 64, 244–251. [Google Scholar] [CrossRef]
- Osman, H.S.; Salim, B.B. Improving yield and quality of kohlrabi stems growing under NaCl salinity using foliar application of urea and seaweed extract. J. Hort. Sci. Ornament. Plant. 2016, 8, 149–160. [Google Scholar]
- Osman, H.S.; Salim, B.B.M. Enhancing antioxidants defense system of snap bean under NaCl salinity using foliar application of salicylic acid, spermidine and glycine betaine. Am. Eurasian J. Agric. Environ. Sci. 2016, 16, 1200–1210. [Google Scholar]
- Wang, B.; Zhang, J.; Pei, D.; Yu, L. Combined effects of water stress and salinity on growth, physiological, and biochemical traits in two walnut genotypes. Physiol. Plant. 2021, 172, 176–187. [Google Scholar] [CrossRef]
- Osman, H.S. Enhancing antioxidant–yield relationship of pea plant under drought at different growth stages by exogenously applied glycine betaine and proline. Ann. Agric. Sci. 2015, 60, 389–402. [Google Scholar] [CrossRef] [Green Version]
- Hafez, E.; Omara, A.E.D.; Ahmed, A. The coupling effects of plant growth promoting rhizobacteria and salicylic acid on physiological modifications, yield traits, and productivity of wheat under water deficient conditions. Agronomy 2019, 9, 524. [Google Scholar] [CrossRef] [Green Version]
- Munns, R. Comparative physiology of salt and water stress. Plant. Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Sahin, U.; Ekinci, M.; Ors, S.; Turan, M.; Yildiz, S.; Yildirim, E. Effects of individual and combined effects of salinity and drought on physiological, nutritional and biochemical properties of cabbage (Brassica oleracea var. capitata). Sci. Hortic. 2018, 240, 196–204. [Google Scholar] [CrossRef]
- Araújo, S.S.; Beebe, S.; Crespi, M.; Delbreil, B.; González, E.M.; Gruber, V.; Lejeune-Henaut, I.; Link, W.; Monteros, M.J.; Prats, E.; et al. Abiotic stress responses in legumes: Strategies used to cope with environmental challenges. Crit. Rev. Plant. Sci. 2015, 34, 237–280. [Google Scholar] [CrossRef]
- Pagano, M.C.; Miransari, M. The importance of soybean production worldwide. In Abiotic and Biotic Stresses in Soybean Production; Miransari, M., Ed.; Academic Press: San Diego, CA, USA, 2016; pp. 1–26. [Google Scholar]
- Han, H.S.; Lee, K.D. Physiological responses of soybean-inoculation of Bradyrhizobium japonicum with PGPR in saline soil conditions. Res. J. Agric. Biol. Sci. 2005, 1, 216–221. [Google Scholar]
- Maas, E.V.; Grattan, S.R. Crop yields as affected by salinity. In Agricultural Drainage; Skaggs, R.W., van Schilfgaarde, J., Eds.; Wiley: Hoboken, NJ, USA, 1999; pp. 55–108. [Google Scholar]
- Ku, Y.-S.; Au-Yeung, W.-K.; Yung, Y.-L.; Li, M.-W.; Wen, C.-Q.; Liu, X.; Lam, H.-M. Drought stress and tolerance in soybean. In A Comprehensive Survey of International Soybean Research—Genetics, Physiology, Agronomy and Nitrogen Relationships; Board, J.E., Ed.; IntechOpen: London, UK, 2013. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Jensen, C.R.; Andersen, M.N. Drought stress effect on carbohydrate concentration in soybean leaves and pods during early reproductive development: Its implication in altering pod set. Field Crops Res. 2004, 86, 1–13. [Google Scholar] [CrossRef]
- Hafez, E.; Farig, M. Efficacy of salicylic acid as a cofactor for ameliorating effects of water stress and enhancing wheat yield and water use efficiency in saline soil. Int. J. Plant. Prod. 2019, 13, 163–176. [Google Scholar] [CrossRef]
- Hafez, E.M.; Alsohim, A.S.; Farig, M.; Omara, A.E.-D.; Rashwan, E.; Kamara, M.M. Synergistic effect of biochar and plant growth promoting rhizobacteria on alleviation of water deficit in rice plants under salt-affected soil. Agronomy 2019, 9, 847. [Google Scholar] [CrossRef] [Green Version]
- Hafez, E.M.; Omara, A.E.D.; Alhumaydhi, F.A.; El-Esawi, M.A. Minimizing hazard impacts of soil salinity and water stress on wheat plants by soil application of vermicompost and biochar. Physiol. Plant. 2020, 1–16. [Google Scholar] [CrossRef]
- Hafez, E.M.; Osman, H.S.; Gowayed, S.M.; Okasha, S.A.; Omara, A.E.-D.; Sami, R.; Abd El-Monem, A.M.; Abd El-Razek, U.A. Minimizing the adversely impacts of water deficit and soil salinity on maize growth and productivity in response to the application of plant growth-promoting rhizobacteria and silica nanoparticles. Agronomy 2021, 11, 676. [Google Scholar] [CrossRef]
- Hafez, E.M.; Osman, H.S.; El-Razek, U.A.A.; Elbagory, M.; Omara, A.E.-D.; Eid, M.A.; Gowayed, S.M. Foliar-applied potassium silicate coupled with plant growth-promoting rhizobacteria improves growth, physiology, nutrient uptake and productivity of faba bean (Vicia faba L.) irrigated with saline water in salt-affected soil. Plants 2021, 10, 894. [Google Scholar] [CrossRef] [PubMed]
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The rhizosphere microbiome: Significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol. Rev. 2013, 37, 634–663. [Google Scholar] [CrossRef] [PubMed]
- Olanrewaju, O.S.; Glick, B.R.; Babalola, O.O. Mechanisms of action of plant growth promoting bacteria. World J. Microbiol. Biotechnol. 2017, 33, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Dias, M.C.; Freitas, H. Drought and salinity stress responses and microbe-induced tolerance in plants. Front. Plant. Sci. 2020, 11. [Google Scholar] [CrossRef]
- Kumar, A.; Verma, J.P. Does plant—Microbe interaction confer stress tolerance in plants: A review? Microbiol. Res. 2018, 207, 41–52. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Wirth, S.J.; Alqarawi, A.A.; Abd Allah, E.F.; Hashem, A. Phytohormones and beneficial microbes: Essential components for plants to balance stress and fitness. Front. Microbiol. 2017, 8, 2104. [Google Scholar] [CrossRef]
- Solyman, S.N.; Abdel-Monem, M.; Abou-Taleb, K.; Osman, H.S.; El-Sharkawy, R.M. Production of plant growth regulators by some fungi isolated under salt stress. South Asian J. Res. Microbiol. 2019, 3, 1–10. [Google Scholar] [CrossRef]
- Masciarelli, O.; Llanes, A.; Luna, V. A new PGPR co-inoculated with Bradyrhizobium japonicum enhances soybean nodulation. Microbiol. Res. 2014, 169, 609–615. [Google Scholar] [CrossRef] [PubMed]
- Egamberdieva, D.; Wirth, S.; Jabborova, D.; Räsänen, L.A.; Liao, H. Coordination between Bradyrhizobium and Pseudomonas alleviates salt stress in soybean through altering root system architecture. J. Plant. Interact. 2017, 12, 100–107. [Google Scholar] [CrossRef] [Green Version]
- Hermosa, R.; Viterbo, A.; Chet, I.; Monte, E. Plant-beneficial effects of Trichoderma and of its genes. Microbiology 2012, 158, 17–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mona, S.A.; Hashem, A.; Abd_Allah, E.F.; Alqarawi, A.A.; Soliman, D.W.K.; Wirth, S.; Egamberdieva, D. Increased resistance of drought by Trichoderma harzianum fungal treatment correlates with increased secondary metabolites and proline content. J. Integrat. Agric. 2017, 16, 1751–1757. [Google Scholar] [CrossRef]
- Marra, R.; Lombardi, N.; d’Errico, G.; Troisi, J.; Scala, G.; Vinale, F.; Woo, S.L.; Bonanomi, G.; Lorito, M. Application of Trichoderma strains and metabolites enhances soybean productivity and nutrient content. J. Agric. Food Chemist. 2019, 67, 1814–1822. [Google Scholar] [CrossRef]
- Broadley, M.; Brown, P.; Cakmak, I.; Rengel, Z.; Zhao, F. Function of nutrients: Micronutrients. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Academic Press: San Diego, CA, USA, 2012; pp. 191–248. [Google Scholar]
- Kheir, A.M.S.; Abouelsoud, H.M.; Hafez, E.M.; Ali, O.A.M. Integrated effect of nano-Zn, nano-Si, and drainage using crop straw–filled ditches on saline sodic soil properties and rice productivity. Arab. J. Geosci. 2019, 12, 471. [Google Scholar] [CrossRef]
- Osman, H.S.; Abd El-Gawad, H.G. Impact of stimulators of amylase activity (GA3, CaCl2) and protein synthesis (ZnSO4) on yield, quality and reducing seed abortion of pea plant. Res. J. Agric. Biol. Sci. 2013, 9, 381–390. [Google Scholar]
- Batista, P.F.; Müller, C.; Merchant, A.; Fuentes, D.; Silva-Filho, R.d.O.; da Silva, F.B.; Costa, A.C. Biochemical and physiological impacts of zinc sulphate, potassium phosphite and hydrogen sulphide in mitigating stress conditions in soybean. Physiol. Plant. 2020, 168, 456–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youssif, N.E.E.; Osman, H.S.M.; Salama, Y.A.M.; Zaghlool, S.A.M. Effect of rice straw and applications of potassium silicate, potassium humate and seaweed extract on growth and some macronutrients of sweet pepper plants under irrigation deficit. Arab Univ. J. Agric. Sci. 2018, 26, 755–773. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Refay, Y.; Al-Suhaibani, N.; Al-Ashkar, I.; El-Hendawy, S.; Hafez, E.M. Integrative effects of rice-straw biochar and silicon on oil and seed quality, yield and physiological traits of Helianthus annuus L. grown under water deficit stress. Agronomy 2019, 9, 637. [Google Scholar] [CrossRef] [Green Version]
- Yeo, A.R.; Flowers, S.A.; Rao, G.; Welfare, K.; Senanayake, N.; Flowers, T.J. Silicon reduces sodium uptake in rice (Oryza sativa L.) in saline conditions and this is accounted for by a reduction in the transpirational bypass flow. Plant. Cell Environ. 1999, 22, 559–565. [Google Scholar] [CrossRef]
- Liu, R.; Lal, R. Potentials of engineered nanoparticles as fertilizers for increasing agronomic productions. Sci. Total Environ. 2015, 514, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Boiero, L.; Perrig, D.; Masciarelli, O.; Penna, C.; Cassán, F.; Luna, V. Phytohormone production by three strains of Bradyrhizobium japonicum and possible physiological and technological implications. Appl. Microbiol. Biotech. 2007, 74, 874–880. [Google Scholar] [CrossRef]
- Oliveira, C.M.; Almeida, N.O.; da Rocha, M.R.; Rezende, M.H.; Carneiro, R.G.d.S.; Ulhoa, C.J. Anatomical changes induced by isolates of Trichoderma spp. in soybean plants. PLoS ONE 2020, 15, e0242480. [Google Scholar] [CrossRef] [PubMed]
- Pal, V.; Singh, G.; Dhaliwal, S.S. Symbiotic parameters, growth, productivity and profitability of chickpea as influenced by zinc sulphate and urea application. J. Soil Sci. Plant. Nutr. 2020, 20, 738–750. [Google Scholar] [CrossRef]
- Tripathi, P.; Na, C.-I.; Kim, Y. Effect of silicon fertilizer treatment on nodule formation and yield in soybean (Glycine max L.). Eur. J. Agron. 2021, 122, 126172. [Google Scholar] [CrossRef]
- Osman, H.S.; Salim, B.B.M. Influence of exogenous application of some phytoprotectants on growth, yield and pod quality of snap bean under NaCl salinity. Ann. Agric. Sci. 2016, 61, 1–13. [Google Scholar] [CrossRef] [Green Version]
- McKeague, J.A. Manual on Soil Sampling and Methods of Analysis, 2nd ed.; Canadian Society of Soil Science: Ottawa, ON, Canada, 1978; 212p. [Google Scholar]
- Mclean, E.O. Soil pH and Lime Requirement. In Methods of Soil Analysis; Page, A.L., Ed.; ASA-SSSA; Soil Science Society of America: Madison, WI, USA, 1983; pp. 199–224. [Google Scholar]
- El-Nahrawy, S.; Elbagory, M.; Omara, A.E.-D. Biocompatibility effect of Bradyrhizobium japonicum and Trichoderma strains on growth, nodulation and physiological traits of soybean (Glycine max l.) under water deficit conditions. J. Adv. Microbiol. 2020, 20, 52–66. [Google Scholar] [CrossRef]
- Atlas, R.M. Handbook of Microbiological Media, 4th ed.; CRC press: Boca Raton, FL, USA, 2010; 2040p. [Google Scholar]
- Seilsepour, M.; Rashidi, M.; Khabbaz, B.G. Prediction of soil exchangeable sodium percentage based on soil sodium adsorption ratio. Am. Eurasian J. Agric. Environ. Sci. 2009, 5, 1–4. [Google Scholar]
- Chapman, H.D.; Pratt, P.F. Methods of Analysis for Soils, Plants and Waters. Soil Sci. 1962, 93, 68. [Google Scholar] [CrossRef] [Green Version]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. In Methods in Enzymology; Academic Press: San Diego, CA, USA, 1987; Volume 148, pp. 350–382. [Google Scholar]
- Hendrix, D.L. Rapid extraction and analysis of nonstructural carbohydrates in plant tissues. Crop. Sci. 1993, 33, 1306–1311. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant. Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Misra, P.S.; Mertz, E.T.; Glover, D.V. Studies on corn proteins. VIII. Free amino acid content of opaque-2 double mutants. Cereal Chem. 1975, 52, 844–848. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analyt. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. In Methods in Enzymology; Academic Press: San Diego, CA, USA, 1984; pp. 121–126. [Google Scholar]
- Vetter, J.L.; Steinberg, M.P.; Nelson, A.I. Enzyme assay, quantitative determination of peroxidase in sweet corn. J. Agric. Food Chemist. 1958, 6, 39–41. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Analyt. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Li, Q.-T.; Yeo, M.H.; Tan, B.K. Lipid peroxidation in small and large phospholipid unilamellar vesicles induced by water-soluble free radical sources. Biochem. Biophysic. Res. Communic. 2000, 273, 72–76. [Google Scholar] [CrossRef] [PubMed]
- Bajji, M.; Kinet, J.-M.; Lutts, S. The use of the electrolyte leakage method for assessing cell membrane stability as a water stress tolerance test in durum wheat. Plant. Growth Regul. 2002, 36, 61–70. [Google Scholar] [CrossRef]
- Weatherley, P.E. Studies in the water relations of the cotton plant. I. The field measurement of water deficits in leaves. New Phytol. 1950, 49, 81–97. [Google Scholar] [CrossRef]
- A.O.A.C. Official Methods of Analysis of AOAC International, 18th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 2005. [Google Scholar]
- Sadasivam, S.; Manickam, A. Biochemical Methods, 3rd ed.; New Age International Publishers: New Delhi, India, 2010. [Google Scholar]
- Duncan, D.B. Multiple range and multiple F tests. Biometrics 1955, 11, 1–42. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Season | Irrigation | Treatments | Pods Plant−1 (No.) | 100-Seed (g) | Seed Yield (t ha−1) | Protein Seed−1 (%) | Carb. Seed−1 (%) |
|---|---|---|---|---|---|---|---|
| 2019 | IW0 | Control | 82.4 ± 5.2 de | 16.1 ± 0.8 c | 1.7 ± 0.08 e | 28.1 ± 1.0 cd | 20.2 ± 1.2 c |
| PGPMs | 99.7 ± 4.7 c | 17.5 ± 1.1 b | 2.0 ± 0.14 c | 33.1 ± 1.1 b | 23.4 ± 1.2 b | ||
| Si-ZnNPs | 105.6 ± 4.0 b | 18.0 ± 1.0 b | 2.1 ± 0.11 b | 35.1 ± 1.4 b | 26.0 ± 1.0 a | ||
| PGPMs + Si-ZnNPs | 115.3 ± 4.1 a | 19.4 ± 1.3 a | 2.3 ± 0.12 a | 38.3 ± 1.6 a | 27.9 ± 1.0 a | ||
| IW1 | Control | 64.6 ± 3.8 h | 13.7 ± 0.8 eg | 1.3 ± 0.07 h | 18.4 ± 1.4 g | 15.9 ± 0.9 ef | |
| PGPMs | 79.6 ± 3.2 e | 14.5 ± 0.5 de | 1.5 ± 0.09 f | 23.8 ± 1.2 e | 18.7 ± 0.8 cd | ||
| Si-ZnNPs | 86.4 ± 3.0 d | 15.2 ± 1.0 cd | 1.6 ± 0.11 e | 26.3 ± 1.3 d | 19.9 ± 1.1 c | ||
| PGPMs + Si-ZnNPs | 96.1 ± 4.6 c | 16.3 ± 1.3 c | 1.8 ± 0.09 d | 30.1 ± 1.5 c | 22.2 ± 1.1 b | ||
| IW2 | Control | 57.7 ± 2.5 i | 11.9 ± 0.7 h | 0.9 ± 0.07 k | 11.3 ± 1.1 i | 11.4 ± 1.0 h | |
| PGPMs | 70.4 ± 4.3 g | 12.5 ± 0.9 gh | 1.1 ± 0.07 j | 14.4 ± 1.3 h | 13.1 ± 0.9 gh | ||
| Si-ZnNPs | 73.2 ± 3.3 fg | 13.2 ± 1.1 fg | 1.2 ± 0.06 i | 16.7 ± 0.9 gh | 14.4 ± 0.6 fg | ||
| PGPMs + Si-ZnNPs | 76.9 ± 4.4 ef | 13.8 ± 1.1 ef | 1.4 ± 0.08 g | 20.9 ± 1.91 f | 17.2 ± 1.5 de | ||
| 2020 | IW0 | Control | 84.0 ± 4.1 e | 16.3 ± 0.9 cd | 1.8 ± 0.09 d | 29.5 ± 0.9 de | 21.3 ± 1.9 cd |
| PGPMs | 102.3 ± 2.4 c | 17.3 ± 0.9 bc | 2.1 ± 0.07 c | 33.9 ± 1.9 c | 24.7 ± 1.1 b | ||
| Si-ZnNPs | 107.8 ± 3.5 b | 18.2 ± 1.1 b | 2.2 ± 0.05 b | 36.5 ± 1.1 b | 27.2 ± 0.9 a | ||
| PGPMs + Si-ZnNPs | 118.3 ± 3.9 a | 20.5 ± 0.9 a | 2.5 ± 0.11 a | 39.7 ± 1.2 a | 28.8 ± 1.3 a | ||
| IW1 | Control | 68.9 ± 2.7 f | 14.9 ± 0.9 ef | 1.3 ± 0.09 f | 19.6 ± 1.4 h | 16.8 ± 1.1 fg | |
| PGPMs | 83.3 ± 2.3 e | 15.5 ± 0.9 df | 1.6 ± 0.09 e | 25.2 ± 1.4 f | 19.4 ± 1.2 de | ||
| Si-ZnNPs | 90.4 ± 3.3 d | 16.1 ± 0.9 ce | 1.8 ± 0.08 d | 27.7 ± 1.2 e | 20.6 ± 1.0 cd | ||
| PGPMs + Si-ZnNPs | 102.2 ± 4.6 c | 16.9 ± 0.8 c | 2.0 ± 0.11 c | 31.1 ± 1.7 d | 22.6 ± 1.1 c | ||
| IW2 | Control | 60.5 ± 4.5 g | 13.1 ± 1.1 g | 1.0 ± 0.08 g | 12.5 ± 0.9 j | 12.5 ± 1.1 i | |
| PGPMs | 71.0 ± 3.1 f | 14.3 ± 1.0 fg | 1.3 ± 0.08 f | 15.6 ± 1.1 i | 14.3 ± 0.7 hi | ||
| Si-ZnNPs | 73.5 ± 3.7 f | 14.6 ± 1.0 f | 1.3 ± 0.07 f | 18.4 ± 1.2 h | 15.7 ± 0.9 gh | ||
| PGPMs + Si-ZnNPs | 83.3 ± 5.4 e | 15.4 ± 1.0 df | 1.6 ± 0.06 e | 22.2 ± 2.1 g | 18.4 ± 1.6 ef | ||
| F test | Deficit irrigation | *** | ** | *** | *** | *** | |
| Soil and foliar treatments | *** | *** | *** | *** | *** | ||
| Irrigation X Treatments | * | ns | ** | ns | ns | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Osman, H.S.; Gowayed, S.M.; Elbagory, M.; Omara, A.E.-D.; El-Monem, A.M.A.; Abd El-Razek, U.A.; Hafez, E.M. Interactive Impacts of Beneficial Microbes and Si-Zn Nanocomposite on Growth and Productivity of Soybean Subjected to Water Deficit under Salt-Affected Soil Conditions. Plants 2021, 10, 1396. https://doi.org/10.3390/plants10071396
Osman HS, Gowayed SM, Elbagory M, Omara AE-D, El-Monem AMA, Abd El-Razek UA, Hafez EM. Interactive Impacts of Beneficial Microbes and Si-Zn Nanocomposite on Growth and Productivity of Soybean Subjected to Water Deficit under Salt-Affected Soil Conditions. Plants. 2021; 10(7):1396. https://doi.org/10.3390/plants10071396
Chicago/Turabian StyleOsman, Hany S., Salah M. Gowayed, Mohssen Elbagory, Alaa El-Dein Omara, Ahmed M. Abd El-Monem, Usama A. Abd El-Razek, and Emad M. Hafez. 2021. "Interactive Impacts of Beneficial Microbes and Si-Zn Nanocomposite on Growth and Productivity of Soybean Subjected to Water Deficit under Salt-Affected Soil Conditions" Plants 10, no. 7: 1396. https://doi.org/10.3390/plants10071396
APA StyleOsman, H. S., Gowayed, S. M., Elbagory, M., Omara, A. E. -D., El-Monem, A. M. A., Abd El-Razek, U. A., & Hafez, E. M. (2021). Interactive Impacts of Beneficial Microbes and Si-Zn Nanocomposite on Growth and Productivity of Soybean Subjected to Water Deficit under Salt-Affected Soil Conditions. Plants, 10(7), 1396. https://doi.org/10.3390/plants10071396