
Unraveling the Influence of Land-Use Change on δ13C, δ15N, and Soil Nutritional Status in Coniferous, Broadleaved, and Mixed Forests in Southern China: A Field Investigation

 ,
, 
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
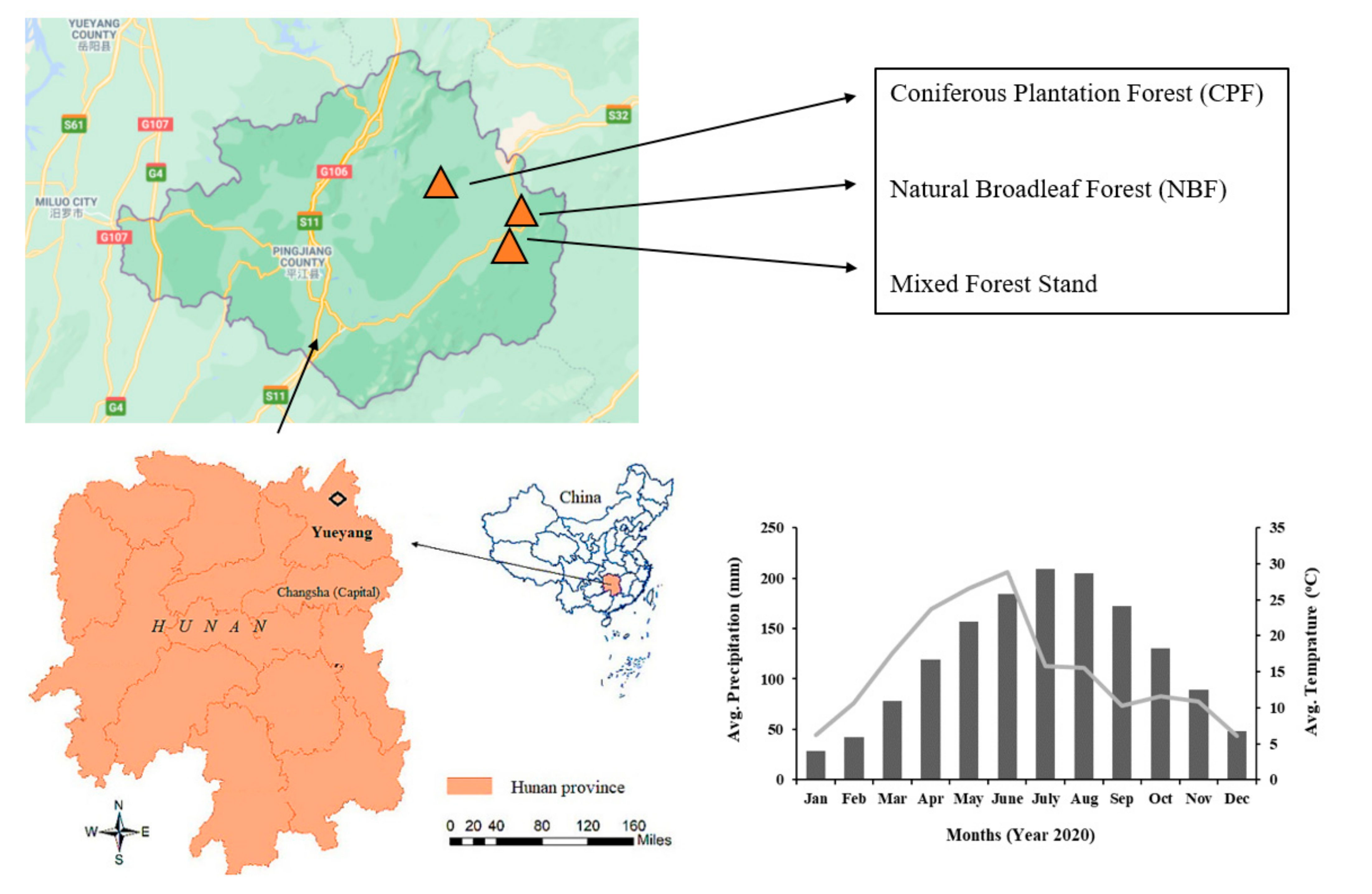
2.1. Study Area
2.2. Forest Types and Species
- The mid-subtropical zone of China is covered with natural forests dominated by broadleaf evergreen species, and Castanopsis eyeri (Fagaceae) is one of them [34]. C. eyeri is a vital part of forest ecosystems and provides services such as water conservation, biodiversity protection, biomass maintenance, and local climate regulation [35]. The C. eyeri (NBF) stand in the research station is the largest and most complete community of Castonpsis, and therefore has crucial protection, scientific research, and landscape values. The forest is neat, and the forest canopy undulating. At the Lutou forest farm, it is mainly distributed on mountain slopes. The average slope gradient is 20–25°. The shrub layer includes species such as Rhododendron simsii, Ilex formosana, Daphniphyllum oldhami, Toxicodendron vernicifluum, Cyclobalanopsis glauca, Eurya tetragonoclada, Symplocos sumuntia, Rhododendron simiarum, Lindera aggregate, Eurya muricat, Dendropanax dentiger, and Myrica rubra.
- Chinese fir (Cunninghamia lanceolata) is a typical evergreen coniferous timber tree species. The pure natural forest of C. lanceolata is rare to find, and mainly presents as an artificial forest plantation. The CPF stand at Lutou forest station are present on both flat soil and mountain slopes. The average slope gradient is 18–23°. Besides timber production, climate regulation, and soil and water conservation, it also provides biomass energy [21,36]. The CPF stand was established 30 years ago under the Chinese afforestation program for forest area enhancement in 1980–1990. It was established as a plantation forest after clearing the site. The shrub layers’ primary species include Litsea cubeba, Eurya mauricata, Rhus chinensis, Sapium discolor, Rhododendron mariesii, Diplospora dubia, Ilex chinensis, and Rubus lambertianus.
- The P. massoniana + C. eyeri mixed forest is an important forest type at Lutou forest station. In Lutou, it is distributed in the ridges, hillside, and mountain slopes. The average slope gradient was 20–25°. P. massoniana is an evergreen coniferous species that has been widely planted in the red soils of southern China since 1980, mainly for soil conservation purposes [37,38]. At the Lutou forest ecosystem observation and research station, the mixed forest stand is formed by planting P. massoniana in the natural regenerated C. eyeri. The shrub layer primarily comprises Loropetalum chinensis, Rhododendron longipetalon, Ilex latifolia, Camellia oleifera, Phobe hupehensis, Ilex ficifolia, Eugenia glabra, Rubus cranbergii, Rata thunbergia, etc.
2.3. Soil and Foliar Sampling
2.4. Isotopic 13C, 15N Abundance, and C, N Content Analysis
2.5. Determination of Soil Available Nutrients
2.6. Data Analysis
3. Results
3.1. Soil and Foliar TC, TN Content, and C:N Ratio
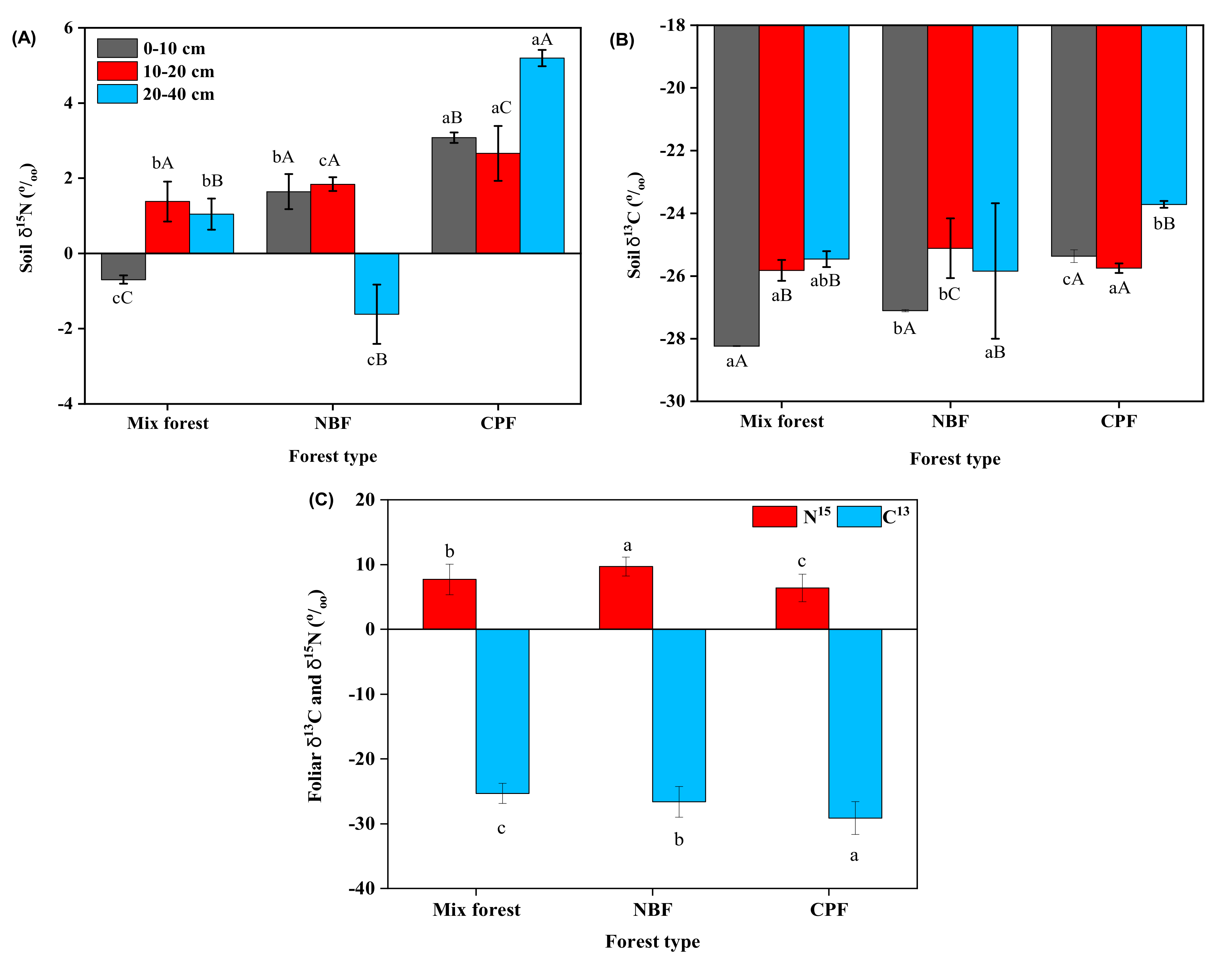
3.2. C13 and 15N Natural Abundance
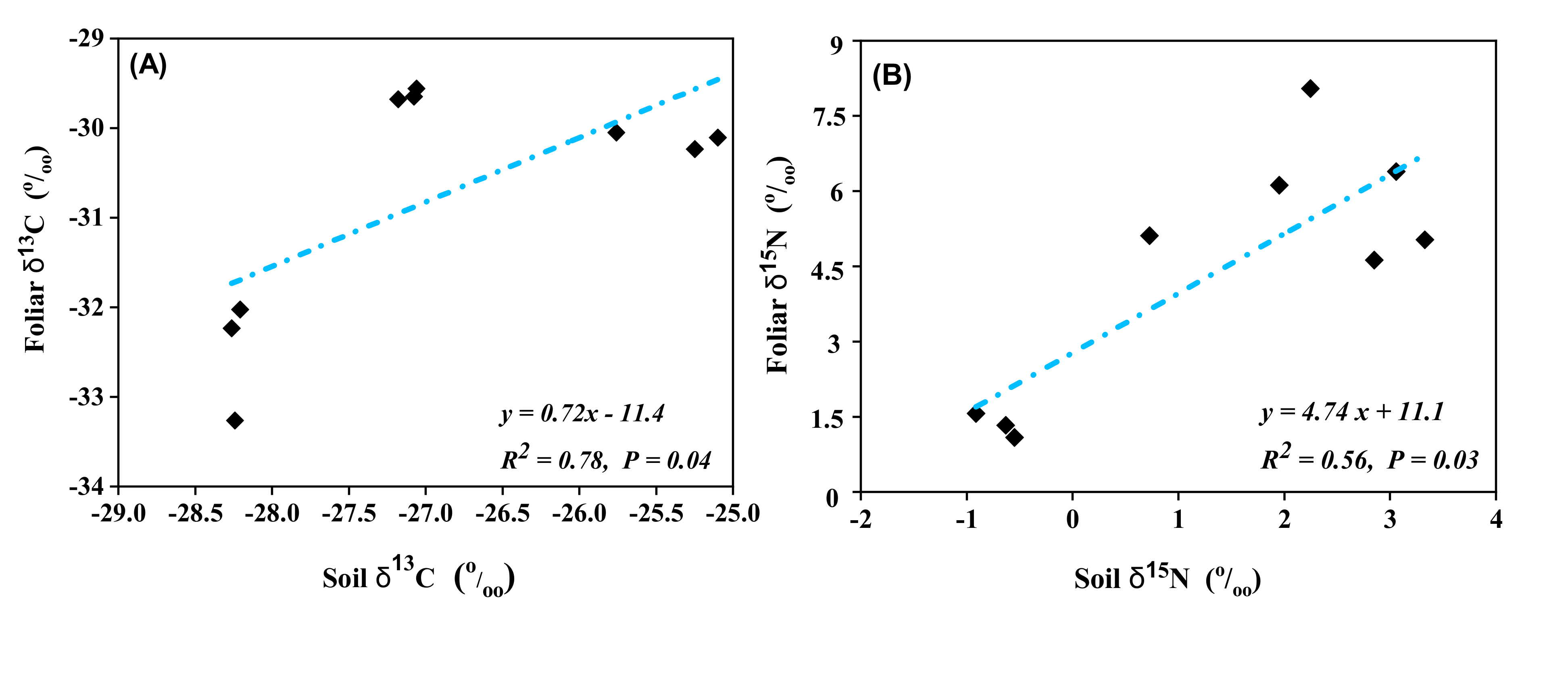
3.3. Relationship between Soil, Foliar Isotopic Abundance, and C:N Ratio
3.4. Correlation between Isotopic Abundance and Potentially Influential Factors
3.5. Soil Nutrient Availability
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Variable | Forest Type | Soil Depth | Forest Type × Soil Depth |
|---|---|---|---|
| TN | <0.05 | <0.001 | 0.02 |
| TC | 0.04 | <0.001 | 0.04 |
| δ15N | <0.001 | 0.048 | <0.001 |
| δ13C | 0.078 | 0.028 | 0.329 |
| C:N | <0.001 | <0.001 | 0.04 |
| SOC | 0.01 | 0.01 | 0.03 |
| AN | 0.04 | <0.001 | 0.02 |
| AP | 0.02 | 0.03 | <0.001 |
| AK | 0.05 | 0.05 | 0.02 |
References
- Ayres, E.; Dromph, K.M.; Cook, R.; Ostle, N.; Bardgett, R.D. The influence of belowground herbivory and defoliation of a legume on nitrogen transfer to neighbouring plants. Funct. Ecol. 2007, 21, 256–263. [Google Scholar] [CrossRef] [Green Version]
- Farooq, T.H.; Yan, W.; Chen, X.; Shakoor, A.; Rashid, M.H.U.; Gilani, M.M.; He, Z.; Wu, P. Dynamics of canopy development of Cunninghamia lanceolata mid-age plantation in relation to foliar nitrogen and soil quality influenced by stand density. Glob. Ecol. Conserv. 2020, 24, e01209. [Google Scholar] [CrossRef]
- Finzi, A.C.; Van Breemen, N.; Canham, C.D. Canopy tree-soil interactions within temperate forests: Species effects on soil carbon and nitrogen. Ecol. Appl. 1998, 8, 440–446. [Google Scholar]
- Macinnis-Ng, C.; Schwendenmann, L. Litterfall, carbon and nitrogen cycling in a southern hemisphere conifer forest dominated by kauri (Agathis australis) during drought. Plant Ecol. 2015, 216, 247–262. [Google Scholar] [CrossRef]
- Chen, C.; Jia, Y.; Chen, Y.; Mehmood, I.; Fang, Y.; Wang, G. Nitrogen isotopic composition of plants and soil in an arid mountainous terrain: South slope versus north slope. Biogeosciences 2018, 15, 369. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G. A large and persistent carbon sink in the world’s forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunn, M.; Spielvogel, S.; Sauer, T.; Oelmann, Y. Temperature and precipitation effects on δ13C depth profiles in SOM under temperate beech forests. Geoderma 2014, 235, 146–153. [Google Scholar] [CrossRef]
- Wang, C.; Houlton, B.Z.; Liu, D.; Hou, J.; Cheng, W.; Bai, E. Stable isotopic constraints on global soil organic carbon turnover. Biogeosciences 2018, 15, 987–995. [Google Scholar] [CrossRef] [Green Version]
- Ngaba, M.J.Y.; Hu, Y.-L.; Bol, R.; Ma, X.-Q.; Jin, S.-F.; Mgelwa, A.S. Effects of land use change from natural forest to plantation on C, N and natural abundance of 13C and 15N along a climate gradient in eastern China. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Natelhoffer, K.J.; Fry, B. Controls on natural nitrogen-15 and carbon-13 abundances in forest soil organic matter. Soil Sci. Soc. Am. J. 1988, 52, 1633–1640. [Google Scholar] [CrossRef]
- Ehleringer, J.R.; Buchmann, N.; Flanagan, L.B. Carbon isotope ratios in belowground carbon cycle processes. Ecol. Appl. 2000, 10, 412–422. [Google Scholar] [CrossRef]
- Guillaume, T.; Damris, M.; Kuzyakov, Y. Losses of soil carbon by converting tropical forest to plantations: Erosion and decomposition estimated by δ13C. Glob. Chang. Biol. 2015, 21, 3548–3560. [Google Scholar] [CrossRef] [PubMed]
- Pardo, L.H.; Hemond, H.F.; Montoya, J.P.; Fahey, T.J.; Siccama, T.G. Response of the natural abundance of 15N in forest soils and foliage to high nitrate loss following clear-cutting. Can. J. For. Res. 2002, 32, 1126–1136. [Google Scholar] [CrossRef]
- Wang, A.; Fang, Y.T.; Chen, D.X.; Koba, K.; Makabe, A.; Luo, T.-S.; Yoh, M. Variations in nitrogen-15 natural abundance of plant and soil systems in four remote tropical rainforests, southern China. Oecologia 2014, 174, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Mgelwa, A.S.; Hu, Y.L.; Liu, J.-F.; Qiu, Q.; Liu, Z.; Ngaba, M.J.Y. Differential patterns of nitrogen and δ15N in soil and foliar along two urbanized rivers in a subtropical coastal city of southern China. Environ. Pollut. 2019, 244, 907–914. [Google Scholar] [CrossRef]
- Robinson, D. δ15N as an integrator of the nitrogen cycle. Trends Ecol. Evol. 2001, 16, 153–162. [Google Scholar] [CrossRef]
- Lim, S.S.; Kwak, J.H.; Lee, K.S.; Chang, S.X.; Yoon, K.-S.; Kim, H.-Y.; Choi, W.-J. Soil and plant nitrogen pools in paddy and upland ecosystems have contrasting δ15N. Biol. Fertil. Soils 2015, 51, 231–239. [Google Scholar] [CrossRef]
- Denk, T.R.A.; Mohn, J.; Decock, C.; Lewicka-Szczebak, D.; Harris, E.; Butterbach-Bahl, K.; Kiese, R.; Wolf, B. The nitrogen cycle: A review of isotope effects and isotope modeling approaches. Soil Biol. Biochem. 2017, 105, 121–137. [Google Scholar] [CrossRef] [Green Version]
- Farooq, T.H.; Tigabu, M.; Ma, X.; Zou, X.; Liu, A.; Odén, P.C.; Wu, P. Nutrient uptake, allocation and biochemical changes in two Chinese fir cuttings under heterogeneous phosphorus supply. iForest 2018, 11, 411–417. [Google Scholar] [CrossRef] [Green Version]
- Farooq, T.H.; Wu, W.; Tigabu, M.; Ma, X.; Zou, X.H.; Rashid, M.H.U.; Gilani, M.M.; Wu, P.F. Growth, biomass production and root development of Chinese fir in relation to initial planting density. Forests 2019, 10, 236. [Google Scholar] [CrossRef] [Green Version]
- Farooq, T.H.; Yan, W.; Rashid, M.H.U.; Tigabu, M.; Gilani, M.M.; Zou, X.H.; Wu, P.F. Chinese fir (Cunninghamia Lanceolata) a green gold of China with continues decline in its productivity over the successive rotations: A review. Appl. Ecol. Environ. Res. 2019, 17, 11055–11067. [Google Scholar] [CrossRef]
- Shakoor, A.; Shakoor, S.; Rehman, A.; Ashraf, F.; Abdullah, M.; Shahzad, S.M.; Farooq, T.H.; Ashraf, M.; Manzoor, M.A.; Altaf, M.M.; et al. Effect of animal manure, crop type, climate zone, and soil attributes on greenhouse gas emissions from agricultural soils—A global meta-analysis. J. Clean. Prod. 2020, 278, 124019. [Google Scholar] [CrossRef]
- Shakoor, A.; Shahzad, S.M.; Chatterjee, N.; Arif, M.S.; Farooq, T.H.; Altaf, M.M.; Tufail, M.A.; Dar, A.A.; Mehmood, T. Nitrous oxide emission from agricultural soils: Application of animal manure or biochar? A global meta-analysis. J. Environ. Manag. 2021, 285, 112170. [Google Scholar] [CrossRef] [PubMed]
- Shakoor, A.; Shahbaz, M.; Farooq, T.H.; Sahar, N.E.; Shahzad, S.M.; Altaf, M.M.; Ashraf, M. A global meta-analysis of greenhouse gases emission and crop yield under no-tillage as compared to conventional tillage. Sci. Total Environ. 2021, 750, 142299. [Google Scholar] [CrossRef]
- Fang, Y.; Gundersen, P.; Mo, J.; Zhu, W. Nitrogen leaching in response to increased nitrogen inputs in subtropical monsoon forests in southern China. For. Ecol. Manag. 2009, 257, 332–342. [Google Scholar] [CrossRef]
- Anaya, C.A.; Huber-Sannwald, E. Long-term soil organic carbon and nitrogen dynamics after conversion of tropical forest to traditional sugarcane agriculture in East Mexico. Soil Tillage Res. 2015, 147, 20–29. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, J.; Chang, S.X.; Jiang, P.; Zhou, G.; Shen, Z.; Wu, J.; Lin, L.; Wang, Z.; Shen, M. Converting native shrub forests to Chinese chestnut plantations and subsequent intensive management affected soil C and N pools. For. Ecol. Manag. 2014, 312, 161–169. [Google Scholar] [CrossRef]
- Guo, X.; Meng, M.; Zhang, J.; Chen, H.Y.H. Vegetation change impacts on soil organic carbon chemical composition in subtropical forests. Sci. Rep. 2016, 6, 29607. [Google Scholar] [CrossRef]
- Maes, S.L.; Blondeel, H.; Perring, M.P.; Depauw, L.; Brūmelis, G.; Brunet, J.; Decocq, G.; den Ouden, J.; Härdtle, W.; Hédl, R. Litter quality, land-use history, and nitrogen deposition effects on topsoil conditions across European temperate deciduous forests. For. Ecol. Manag. 2019, 433, 405–418. [Google Scholar] [CrossRef] [Green Version]
- Cornwell, W.K.; Cornelissen, J.H.C.; Amatangelo, K.; Dorrepaal, E.; Eviner, V.T.; Godoy, O.; Hobbie, S.E.; Hoorens, B.; Kurokawa, H.; Pérez-Harguindeguy, N. Plant species traits are the predominant control on litter decomposition rates within biomes worldwide. Ecol. Lett. 2008, 11, 1065–1071. [Google Scholar] [CrossRef] [PubMed]
- Shelton, R.E.; Jacobsen, K.L.; McCulley, R.L. Cover crops and fertilization alter nitrogen loss in organic and conventional conservation agriculture systems. Front. Plant Sci. 2018, 8, 2260. [Google Scholar] [CrossRef] [Green Version]
- Thiffault, E.; Hannam, K.D.; Paré, D.; Titus, B.D.; Hazlett, P.W.; Maynard, D.G.; Brais, S. Effects of forest biomass harvesting on soil productivity in boreal and temperate forests—A review. Environ. Rev. 2011, 19, 278–309. [Google Scholar] [CrossRef]
- Ringeval, B.; Augusto, L.; Monod, H.; Van Apeldoorn, D.; Bouwman, L.; Yang, X.; Achat, D.L.; Chini, L.P.; Van Oost, K.; Guenet, B. Phosphorus in agricultural soils: Drivers of its distribution at the global scale. Glob. Chang. Biol. 2017, 23, 3418–3432. [Google Scholar] [CrossRef]
- Hu, Z.H.; Qian, H.Y.; Yu, M.J. The niche of dominant species populations in Castanopsis eyrei forest in Gutian Mountain National Nature Reserve. Acta Ecol. Sin. 2009, 29, 3670–3677. [Google Scholar]
- Ren, Y.; Peng, D.; Pan, J.; Hong, Z.; Ye, G. Relationship between species biodiversity and biomass accumulation in a Castanopsis eyrei Forest in Wuyishan Mountain under different temporal and spatial scales. Sci. Silvae Sin. 2010, 46, 33–38. [Google Scholar]
- Farooq, T.H.; Shakoor, A.; Wu, X.; Li, Y.; Rashid, M.H.U.; Zhang, X.; Gilani, M.M.; Kumar, U.; Chen, X.; Yan, W. Perspectives of plantation forests in the sustainable forest development of China. iForest 2021, 14, 166. [Google Scholar] [CrossRef]
- Cao, L.; Liang, Y.; Wang, Y.; Lu, H. Runoff and soil loss from Pinus massoniana forest in southern China after simulated rainfall. Catena 2015, 129, 1–8. [Google Scholar] [CrossRef]
- Gilani, M.M.; Tigabu, M.; Liu, B.; Farooq, T.H.; Rashid, M.H.U.; Ramzan, M.; Ma, X. Seed germination and seedling emergence of four tree species of southern China in response to acid rain. J. For. Res. 2020, 32, 471–481. [Google Scholar] [CrossRef]
- Ryan, J.; Estefan, G.; Rashid, A. Soil and Plant Analysis Laboratory Manual; ICARDA: Beirut, Lebanon, 2007. [Google Scholar]
- Bray, R.H.; Kurtz, L.T. Determination of total, organic, and available forms of phosphorus in soils. Soil Sci. 1945, 59, 39–46. [Google Scholar] [CrossRef]
- Schollenberger, C.; Simon, R. Determination of exchange capacity and exchangeable bases in soil ammonium acetate method. Soil Sci. 1945, 59, 13–24. [Google Scholar] [CrossRef]
- Farooq, T.H.; Ma, X.; Rashid, M.H.U.; Wu, W.; Xu, J.; Tarin, M.W.K.; He, Z.; Wu, P. Impact of stand density on soil quality in Chinese Fir (Cunninghamia Lanceolata) monoculture. Appl. Ecol. Environ. Res. 2019, 17, 3553–3566. [Google Scholar] [CrossRef]
- Farooq, T.H.; Kumar, U.; Mo, J.; Shakoor, A.; Wang, J.; Rashid, M.H.U.; Tufail, M.A.; Chen, X.; Yan, W. Intercropping of Peanut–Tea Enhances Soil Enzymatic Activity and Soil Nutrient Status at Different Soil Profiles in Subtropical Southern China. Plants 2021, 10, 881. [Google Scholar] [CrossRef] [PubMed]
- Obalum, S.E.; Chibuike, G.U.; Peth, S.; Ouyang, Y. Soil organic matter as sole indicator of soil degradation. Environ. Monit. Assess. 2017, 189, 176. [Google Scholar] [CrossRef] [PubMed]
- Lal, R. Managing organic matter content for restoring health and ecosystem services of soils of India. J. Indian Soc. Soil Sci. 2020, 68, 1–5. [Google Scholar] [CrossRef]
- Bhowmik, A.; Kukal, S.S.; Saha, D.; Sharma, H.; Kalia, A.; Sharma, S. Potential indicators of soil health degradation in different land use-based ecosystems in the Shiwaliks of Northwestern India. Sustainability 2019, 11, 3908. [Google Scholar] [CrossRef] [Green Version]
- Kooch, Y.; Rostayee, F.; Hosseini, S.M. Effects of tree species on topsoil properties and nitrogen cycling in natural forest and tree plantations of northern Iran. Catena 2016, 144, 65–73. [Google Scholar] [CrossRef]
- Neirynck, J.; Mirtcheva, S.; Sioen, G.; Lust, N. Impact of Tilia platyphyllos Scop., Fraxinus excelsior L., Acer pseudoplatanus L., Quercus robur L. and Fagus sylvatica L. on earthworm biomass and physico-chemical properties of a loamy topsoil. For. Ecol. Manag 2000, 133, 275–286. [Google Scholar] [CrossRef]
- Lovett, G.M.; Weathers, K.C.; Arthur, M.A. Control of Nitrogen Loss from Forested Watersheds by Soil Carbon: Nitrogen Ratio andTree Species Composition. Ecosystems 2002, 5, 712–718. [Google Scholar] [CrossRef]
- Ariz, I.; Cruz, C.; Neves, T.; Irigoyen, J.J.; García, C.; Nogués, S.; Aparicio-Tejo, P.M.; Aranjuelo, I. Leaf δ15N as a physiological indicator of the responsiveness of N2-fixing alfalfa plants to elevated [CO2], temperature and low water availability. Front. Plant Sci. 2015, 6, 574. [Google Scholar] [CrossRef]
- Lawson, S.; Pike, C. Stable isotope ratios and reforestation potential in Acacia koa populations on Hawai’i. Ann. For. Res. 2017. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.S.; Guo, J.; Chen, G.; Xie, J.; Gao, R.; Li, Z.; Jin, Z. Carbon and nitrogen pools in Chinese fir and evergreen broadleaved forests and changes associated with felling and burning in mid-subtropical China. For. Ecol. Manag. 2005, 216, 216–226. [Google Scholar] [CrossRef]
- Rahman, M.M.; Zhang, X.; Ahmed, I.; Iqbal, Z.; Zeraatpisheh, M.; Kanzaki, M.; Xu, M. Remote sensing-based mapping of senescent leaf C: N ratio in the sundarbans reserved forest using machine learning techniques. Remote Sens. 2020, 12, 1375. [Google Scholar] [CrossRef]
- Ellert, B.H.; Janzen, H.H. Long-term biogeochemical cycling in agroecosystems inferred from 13C, 14C and 15N. J. Geochem. Explor. 2006, 88, 198–201. [Google Scholar] [CrossRef]
- Shi, H.; Wang, X.; Xu, M.; Zhang, H.; Luo, Y. Characteristics of soil C: N ratio and δ 13C in wheat-maize cropping system of the North China Plain and influences of the Yellow River. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Aguilos, M.; Stahl, C.; Burban, B.; Hérault, B.; Courtois, E.; Coste, S.; Wagner, F.; Ziegler, C.; Takagi, K.; Bonal, D. Interannual and seasonal variations in ecosystem transpiration and water use efficiency in a tropical rainforest. Forests 2019, 10, 14. [Google Scholar] [CrossRef] [Green Version]
- Phillips, C.L.; Nickerson, N.; Risk, D.; Kayler, Z.E.; Andersen, C.; Mix, A.; Bond, B.J. Soil moisture effects on the carbon isotope composition of soil respiration. Rapid Commun. Mass Spectrom. 2010, 24, 1271–1280. [Google Scholar] [CrossRef] [PubMed]
- Palta, J.A.; Gregory, P.J. Drought affects the fluxes of carbon to roots and soil in 13C pulse-labelled plants of wheat. Soil Biol. Biochem. 1997, 29, 1395–1403. [Google Scholar] [CrossRef]
- Burke, I.C.; Yonker, C.M.; Parton, W.J.; Cole, C.V.; Flach, K.; Schimel, D.S. Texture, climate, and cultivation effects on soil organic matter content in US grassland soils. Soil Sci. Soc. Am. J. 1989, 53, 800–805. [Google Scholar] [CrossRef]
- Selvaraj, S.; Duraisamy, V.; Huang, Z.; Guo, F.; Ma, X. Influence of long-term successive rotations and stand age of Chinese fir (Cunninghamia lanceolata) plantations on soil properties. Geoderma 2017, 306, 127–134. [Google Scholar] [CrossRef]
- Groppo, J.D.; Lins, S.R.M.; de Camargo, P.B.; Assad, E.D.; Pinto, H.S.; Martins, S.C.; Salgado, P.R.; Evangelista, B.; Vasconcellos, E.; Sano, E.E. Changes in soil carbon, nitrogen, and phosphorus due to land-use changes in Brazil. Biogeosciences 2015, 12, 4765–4780. [Google Scholar] [CrossRef] [Green Version]
- Breulmann, M.; Boettger, T.; Buscot, F.; Gruendling, R.; Schulz, E. Carbon storage potential in size–density fractions from semi-natural grassland ecosystems with different productivities over varying soil depths. Sci. Total Environ. 2016, 545, 30–39. [Google Scholar] [CrossRef]
- McGrath, M.J.; Luyssaert, S.; Meyfroidt, P.; Kaplan, J.O.; Burgi, M.; Chen, Y.; Erb, K.; Gimmi, U.; McInerney, D.; Naudts, K. Reconstructing European forest management from 1600 to 2010. Biogeosciences 2015, 12, 4291–4316. [Google Scholar] [CrossRef] [Green Version]
- Wu, P.; Wang, G.; Farooq, T.H.; Li, Q.; Zou, X.; Ma, X. Low phosphorus and competition affect Chinese fir cutting growth and root organic acid content: Does neighboring root activity aggravate P nutrient deficiency? J. Soils Sediments 2017, 17, 2775–2785. [Google Scholar] [CrossRef]
- Rashid, M.H.U.; Tigabu, M.; Chen, H.; Farooq, T.H.; Ma, X.; Wu, P. Calcium-mediated adaptive responses to low phosphorus stress in Chinese fir. Trees 2020, 34, 825–834. [Google Scholar] [CrossRef]
- Saqib, H.M.U.; Ahmad, I.; Rashid, M.H.U.; Farooq, T.H.; Asif, M.; Kashif, M.; Iqbal, A.; Nawaz, M.F. Effect of Compost Application on the Growth of Acacia nilotica. Cercet. Agron. Mold. 2019, 177, 66–73. [Google Scholar] [CrossRef] [Green Version]
- Rashid, M.H.U.; Farooq, T.H.; Iqbal, W.; Asif, M.; Islam, W.; Lin, D.C.; Ahmad, I.; Wu, P.F. Role of indole acetic acid on growth and biomass production of athel tree (tamarix aphylla) by using different cutting lengths. Appl. Ecol. Environ. Res. 2020, 18, 3805–3816. [Google Scholar] [CrossRef]
- Rehman, A.; Arif, M.S.; Tufail, M.A.; Shahzad, S.M.; Farooq, T.H.; Ahmed, W.; Mehmood, T.; Farooq, M.R.; Javed, Z.; Shakoor, A. Biochar potential to relegate metal toxicity effects is more soil driven than plant system: A global meta-analysis. J. Clean. Prod. 2021, 316, 128276. [Google Scholar] [CrossRef]
- Yousaf, M.S.; Farooq, T.H.; Ahmad, I.; Gilani, M.M.; Rashid, M.H.U.; Gautam, N.P.; Islam, W.; Asif, M.; Wu, P. Effect of Drought Stress on the Growth and Morphological Traits of Eucalyptus camaldulensis and Eucalyptus citriodora. PSM Biol. Res. 2018, 3, 85–91. [Google Scholar]
- Wang, J.; Farooq, T.H.; Aslam, A.; Shakoor, A.; Chen, X.; Yan, W. Non-targeted metabolomics reveal the impact of phenanthrene stress on root exudates of ten urban greening tree species. Environ. Res. 2021, 196, 110370. [Google Scholar] [CrossRef]
- Gilani, M.M.; Irfan, A.; Farooq, T.H.; Wu, P.; Yousaf, M.S.; Khan, M.W.; Talha, Y.; Ma, X. Effects of pre-sowing treatments on seed germination and morphological growth of Acacia nilotica and Faidherbia albida. Sci. For. 2019, 122, 374–382. [Google Scholar] [CrossRef]
- Rashid, M.H.U.; Asif, M.; Farooq, T.H.; Gautam, N.P.; Nawaz, M.F.; Ahmad, I.; Gilani, M.M.; Wu, P. Cuttings growth response of Dalbergia sissoo (shisham) to soil compaction stress. Appl. Ecol. Environ. Res. 2019, 17, 1049–1059. [Google Scholar] [CrossRef]
- Lejeune, Q.; Davin, E.L.; Gudmundsson, L.; Winckler, J.; Seneviratne, S.I. Historical deforestation locally increased the intensity of hot days in northern mid-latitudes. Nat. Clim. Chang. 2018, 8, 386–390. [Google Scholar] [CrossRef]



| Mixed Forest | NBF a | CPF | |
|---|---|---|---|
| Species | C. eyeri + P. massoniana b | C. eyeri | C. lanceolata |
| Elevation | 777 m (a.s.l) | 800 m (a.s.l) | 427 m (a.s.l) |
| Soil pH (0–40 cm) | 4.60 ± 0.06 b | 4.40 ± 0.15 c | 4.81 ± 0.02 a |
| Soil BD (0–40 cm) c | 1.28 ± 0.03 a | 1.12 ± 0.08 c | 1.25 ± 0.03 b |
| SMC (%) (0–40 cm) | 16.3 a | 11.63 b | 10.48 b |
| Forest age | 30–60 years | 60 years | 30 years |
| Elements | Depth | Mix Forest | NBF | CPF | |
|---|---|---|---|---|---|
| Soil | TN a (g·kg−1) | 0–10 | 3.33 ± 0.03 aA | 1.56 ± 0.01 cA | 2.10 ± 0.02 bA |
| 10–20 | 1.12 ± 0.03 bB | 0.77 ± 0.01 cB | 1.83 ± 0.01 aB | ||
| 20–40 | 0.79 ± 0.04 bC | 0.56 ± 0.01 cC | 1.19 ± 0.01 aC | ||
| TC (g·kg−1) | 0–10 | 46.88 ± 0.44 aA | 16.63 ± 0.24 cA | 26.02 ± 0.28 bA | |
| 10–20 | 10.19 ± 0.07 bB | 4.17 ± 0.09 cB | 19.40 ± 0.06 aB | ||
| 20–40 | 5.33 ± 0.06 bC | 1.69 ± 0.03 cC | 8.91 ± 0.09 aC | ||
| C:N | 0–10 | 14.06 ± 0.10 aA | 10.65 ± 0.23 bA | 12.39 ± 0.12 aA | |
| 10–20 | 9.07 ± 0.25 bB | 5.40 ± 0.06 cB | 10.62 ± 0.12 cB | ||
| 20–40 | 6.73 ± 0.32 aC | 3.01 ± 0.07 bC | 7.48 ± 0.15 aC | ||
| Foliar | TN (g·kg−1) | - | 27.28 ± 1.96 ab | 28.92 ± 1.43 a | 26.40 ± 0.68 b |
| TC (g·kg−1) | - | 482.23 ± 3.77 b | 495.72 ± 0.27a | 475.30 ± 0.42 c | |
| C:N | - | 15.33 ± 1.56 a | 17.21 ± 0.83 a | 18.02 ± 0.48 a |
| Foliar δ13C | Soil δ13C | Foliar δ15N | Soil δ15N | ||||||
|---|---|---|---|---|---|---|---|---|---|
| r | p | r | p | r | p | r | p | ||
| Foliar TN a | 0.220 | 0.263 | −0.526 ** | 0.001 | Foliar TN | 0.923 ** | 0.001 | −0.414 * | 0.032 |
| Soil TN | 0.118 | 0.551 | −0.470 * | 0.015 | Soil TN | −0.152 | 0.448 | −0.058 | 0.774 |
| Foliar TC | 0.047 | 0.81 | −0.190 | 0.346 | Foliar TC | 0.395 * | 0.041 | −0.526 ** | 0.004 |
| Soil TC | 0.167 | 0.403 | −0.504 ** | 0.012 | Soil TC | −0.124 | 0.536 | −0.126 | 0.531 |
| Foliar C:N | −0.724 ** | 0.001 | 0.551 ** | 0.018 | Foliar C:N | −0.780 ** | 0.001 | 0.404 * | 0.036 |
| Soil C:N | 0.088 | 0.665 | −0.406 * | 0.038 | Soil C:N | −0.194 * | 0.033 | 0.141 * | 0.048 |
| Soil pH | −0.198 | 0.324 | 0.107 | 0.591 | Soil pH | −0.285 | 0.149 | 0.194 | 0.332 |
| BD | 0.567 ** | 0.002 | −0.043 | 0.862 | BD | −0.098 | 0.626 | 0.076 | 0.707 |
| SMC | −0.431 | 0.130 | 0.359 ** | 0.015 | SMC | −0.321 ** | 0.001 | −0.478 ** | 0.001 |
| Forest type | −0.261 | 0.186 | 0.267 | 0.179 | Forest type | −0.463 * | 0.015 | 0.616 ** | 0.001 |
| Soil depth | 0.005 | 1.02 | 0.470 * | 0.013 | Soil depth | 0.005 | 1.01 | 0.040 * | 0.842 |
| Elements | Depth | Mix Forest | NBF | CPF |
|---|---|---|---|---|
| Available N (mg·kg−1) | 0–10 | 177 ± 2.06 aA | 167 ± 2.04 bA | 155 ± 2.2 cA |
| 10–20 | 142 ± 2.49 bB | 155 ± 2.45 aB | 137 ± 1.58 cB | |
| 20–40 | 109 ± 1.0 aC | 105 ± 3.54 bC | 98 ± 1.15 cC | |
| Available P (mg·kg−1) | 0–10 | 62 ± 2.42 bB | 55 ± 0.75 cA | 71 ± 0.69 aA |
| 10–20 | 82 ± 2.46 aA | 54 ± 0.47 bA | 36 ± 1.22 cB | |
| 20–40 | 60 ± 2.59 aB | 38 ± 0.29 bB | 29 ± 0.15 cC | |
| Available K (mg·kg−1) | 0–10 | 147 ± 0.37 aA | 104 ± 1.94 bB | 91 ± 0.81 cA |
| 10–20 | 41 ± 1.02 bB | 119 ± 1.71 aA | 22 ± 0.85 cB | |
| 20–40 | 22 ± 0.07 bC | 41 ± 0.92 aC | 12 ± 0.21 cC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farooq, T.H.; Chen, X.; Shakoor, A.; Li, Y.; Wang, J.; Rashid, M.H.U.; Kumar, U.; Yan, W. Unraveling the Influence of Land-Use Change on δ13C, δ15N, and Soil Nutritional Status in Coniferous, Broadleaved, and Mixed Forests in Southern China: A Field Investigation. Plants 2021, 10, 1499. https://doi.org/10.3390/plants10081499
Farooq TH, Chen X, Shakoor A, Li Y, Wang J, Rashid MHU, Kumar U, Yan W. Unraveling the Influence of Land-Use Change on δ13C, δ15N, and Soil Nutritional Status in Coniferous, Broadleaved, and Mixed Forests in Southern China: A Field Investigation. Plants. 2021; 10(8):1499. https://doi.org/10.3390/plants10081499
Chicago/Turabian StyleFarooq, Taimoor Hassan, Xiaoyong Chen, Awais Shakoor, Yong Li, Jun Wang, Muhammad Haroon U. Rashid, Uttam Kumar, and Wende Yan. 2021. "Unraveling the Influence of Land-Use Change on δ13C, δ15N, and Soil Nutritional Status in Coniferous, Broadleaved, and Mixed Forests in Southern China: A Field Investigation" Plants 10, no. 8: 1499. https://doi.org/10.3390/plants10081499
APA StyleFarooq, T. H., Chen, X., Shakoor, A., Li, Y., Wang, J., Rashid, M. H. U., Kumar, U., & Yan, W. (2021). Unraveling the Influence of Land-Use Change on δ13C, δ15N, and Soil Nutritional Status in Coniferous, Broadleaved, and Mixed Forests in Southern China: A Field Investigation. Plants, 10(8), 1499. https://doi.org/10.3390/plants10081499