
Hydrogen Nanobubble Water Delays Petal Senescence and Prolongs the Vase Life of Cut Carnation (Dianthus caryophyllus L.) Flowers



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
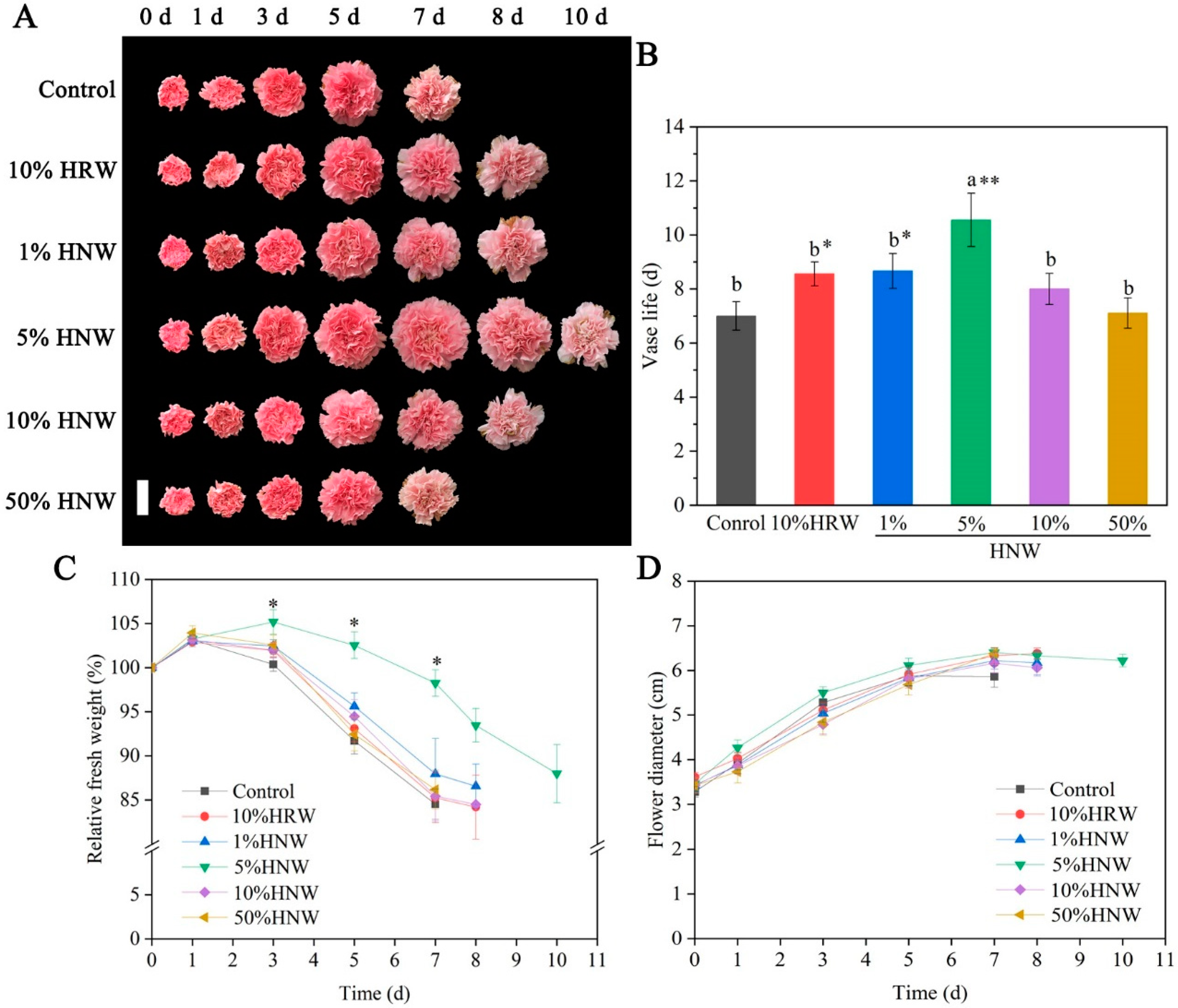
2.1. Effects of HNW on the Vase Life of Cut Carnation Flowers
2.2. Effects of HNW on Relative Water Content and Electrolyte Leakage of Cut Carnation Flowers
2.3. Effects of HNW on Reducing the Oxidative Damage and Cell Death in Carnation Petals
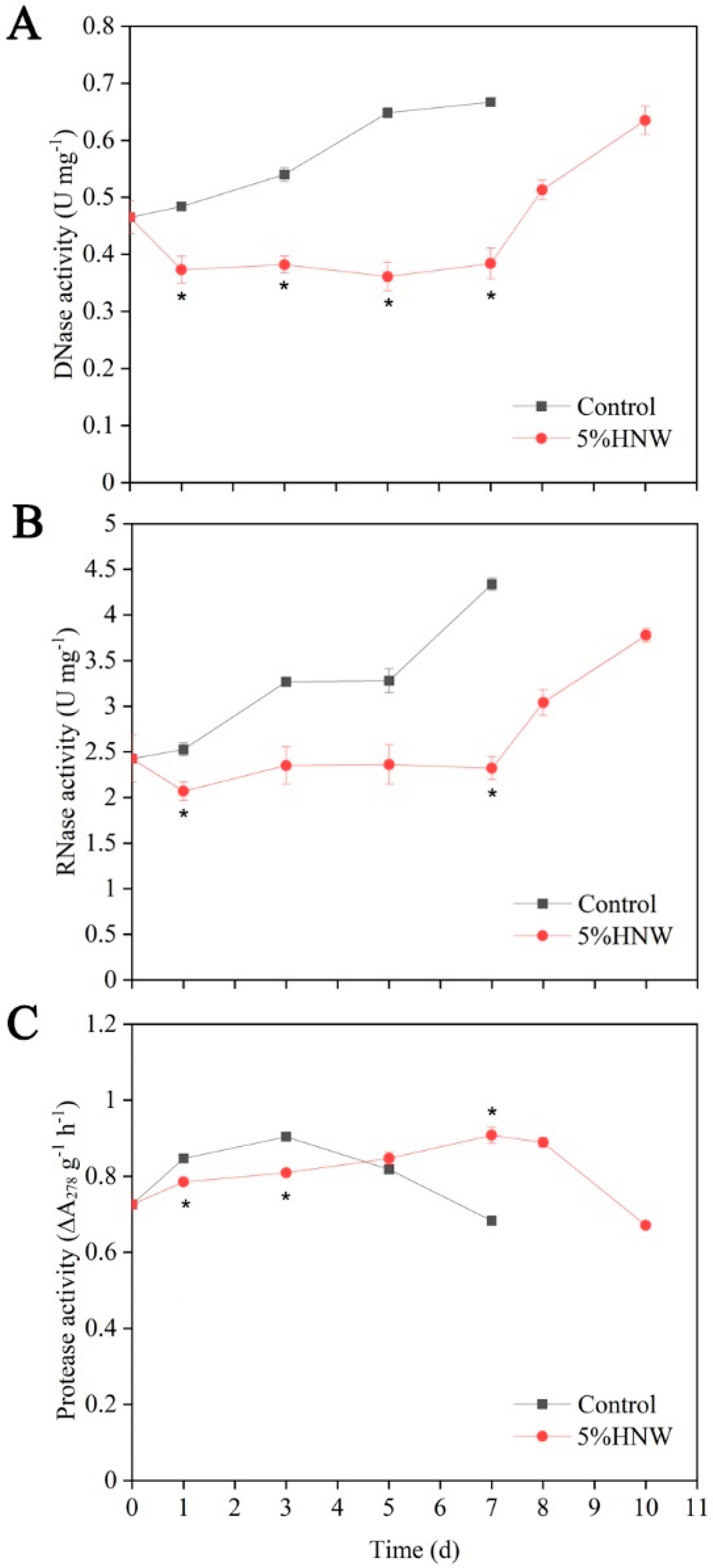
2.4. Effects of HNW on the Activities of DNase, RNase, and Protease
3. Discussion
4. Materials and Methods
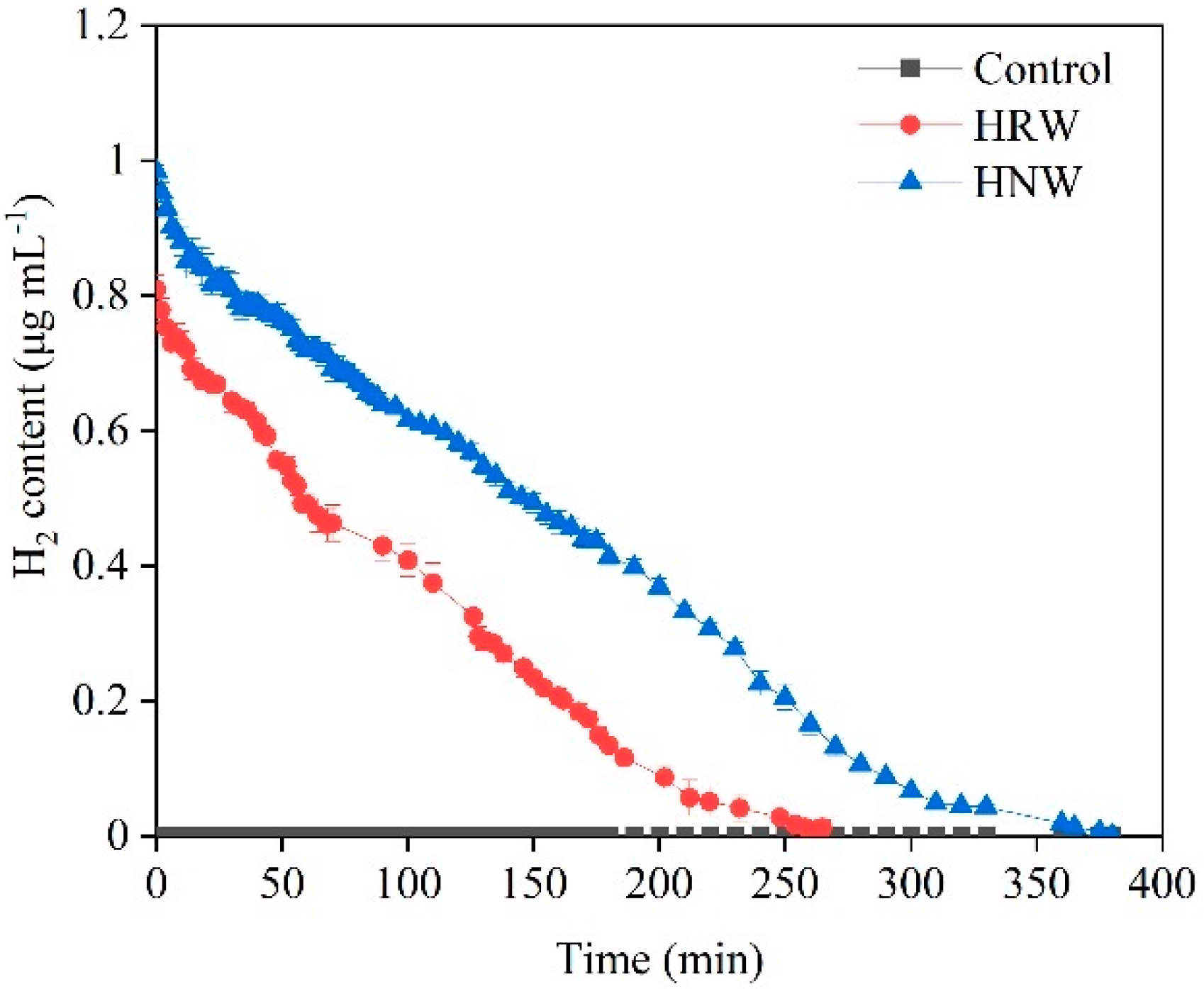
4.1. Preparation of Hydrogen Nanobubble Water and Hydrogen-Rich Water
4.2. Plant Material and Treatments
4.3. Measurement of Vase Life, Fresh Weight, and Flower Diameter
4.4. Determination of Water Content and Electrolyte Leakage of Petals
4.5. Histochemical Staining
4.6. Assays of Enzymatic Activity
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| DAB | 3,3-diaminobenzidine |
| RFW | relative fresh weight |
| H2 | hydrogen gas |
| HNW | hydrogen nanobubble water |
| HRW | hydrogen-rich water |
| H2O2 | hydrogen peroxide |
| NBT | nitro blue tetrazolium |
| O2− | superoxide anions |
| PVP | polyvinylpyridoxone |
| RFW | relative fresh weight |
| ROS | reactive oxygen species |
| RWC | relative water content |
| SE | standard error |
References
- van Doorn, W.G.; Woltering, E.J. Physiology and molecular biology of petal senescence. J. Exp. Bot. 2008, 59, 453–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naing, A.H.; Lee, K.; Arun, M.; Lim, K.B.; Kim, C.K. Characterization of the role of sodium nitroprusside (SNP) involved in long vase life of different carnation cultivars. BMC Plant Biol. 2017, 17, 149. [Google Scholar] [CrossRef] [PubMed]
- Rabiza-Świder, J.; Skutnik, E.; Jędrzejuk, A.; Rochala-Wojciechowska, J. Nanosilver and sucrose delay the senescence of cut snapdragon flowers. Postharvest Biol. Technol. 2020, 165, 111165. [Google Scholar] [CrossRef]
- Langston, B.J.; Bai, S.; Jones, M.L. Increases in DNA fragmentation and induction of a senescence-specific nuclease are delayed during corolla senescence in ethylene-insensitive (etr1-1) transgenic petunias. J. Exp. Bot. 2005, 56, 15–33. [Google Scholar] [CrossRef] [Green Version]
- Panavas, T.; LeVangie, R.; Mistler, J.; Reid, P.D.; Rubinstein, B. Activities of nucleases in senescing daylily petals. Plant Physiol. Biochem. 2000, 38, 837–843. [Google Scholar] [CrossRef]
- Eason, J.R.; Ryan, D.J.; Pinkney, T.T.; O’Donoghue, E.M. Programmed cell death during flower senescence: Isolation and characterization of cysteine proteinases from Sandersonia aurantiaca. Funct. Plant Biol. 2002, 29, 1055–1064. [Google Scholar] [CrossRef] [PubMed]
- Pak, C.; van Doorn, W.G. Delay of Iris flower senescence by protease inhibitors. New Phytol. 2005, 165, 473–480. [Google Scholar] [CrossRef]
- Ohsawa, I.; Ishikawa, M.; Takahashi, K.; Watanabe, M.; Nishimaki, K.; Yamagata, K.; Katsura, K.; Katayama, Y.; Asoh, S.; Ohta, S. Hydrogen acts as a therapeutic antioxidant by selectively reducing cytotoxic oxygen radicals. Nat. Med. 2007, 13, 688–694. [Google Scholar] [CrossRef]
- Ohta, S. Recent progress toward hydrogen medicine: Potential of molecular hydrogen for preventive and therapeutic applications. Curr. Pharm. Des. 2011, 17, 2241. [Google Scholar] [CrossRef] [Green Version]
- Renwick, G.M.; Giumarro, C.; Siegel, S.M. Hydrogen metabolism in higher plants. Plant Physiol. 1964, 39, 303–306. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Zhang, M.; Sun, X. Molecular hydrogen is involved in phytohormone signaling and stress responses in plants. PLoS ONE 2013, 8, e71038. [Google Scholar] [CrossRef]
- Xie, Y.; Mao, Y.; Lai, D.; Zhang, W.; Shen, W. H2 enhances Arabidopsis salt tolerance by manipulating ZAT10/12-mediated antioxidant defence and controlling sodium exclusion. PLoS ONE 2012, 7, e49800. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Mao, Y.; Zhang, W.; Lai, D.; Wang, Q.; Shen, W. Reactive oxygen species-dependent nitric oxide production contributes to hydrogen-promoted stomatal closure in Arabidopsis. Plant Physiol. 2014, 165, 759–773. [Google Scholar] [CrossRef] [Green Version]
- Su, J.; Yang, X.; Shao, Y.; Chen, Z.; Shen, W. Molecular hydrogen–induced salinity tolerance requires melatonin signalling in Arabidopsis thaliana. Plant Cell Environ. 2021, 44, 476–490. [Google Scholar] [CrossRef]
- Cui, W.; Yao, P.; Pan, J.; Dai, C.; Cao, H.; Chen, Z.; Zhang, S.; Xu, S.; Shen, W. Transcriptome analysis reveals insight into molecular hydrogen-induced cadmium tolerance in alfalfa: The prominent role of sulfur and (homo)glutathione metabolism. BMC Plant Biol. 2020, 20, 58. [Google Scholar] [CrossRef]
- Cao, Z.; Duan, X.; Yao, P.; Cui, W.; Cheng, D.; Zhang, J.; Jin, Q.; Chen, J.; Dai, T.; Shen, W. Hydrogen gas is involved in auxin-induced lateral root formation by modulating nitric oxide synthesis. Int. J. Mol. Sci. 2017, 18, 2084. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Su, N.; Jia, L.; Tian, J.; Li, H.; Huang, L.; Shen, Z.; Cui, J. Transcriptome analysis of radish sprouts hypocotyls reveals the regulatory role of hydrogen-rich water in anthocyanin biosynthesis under UV-A. BMC Plant Biol. 2018, 18, 227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, H.; Li, P.; Song, Y.; Gu, R. Hydrogen-rich water delays postharvest ripening and senescence of kiwifruit. Food Chem. 2014, 156, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Zhao, S.; Li, P.; Shen, W. Hydrogen gas prolongs the shelf life of kiwifruit by decreasing ethylene biosynthesis. Postharvest Biol. Technol. 2018, 135, 123–130. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, G.; Cheng, P.; Yan, X.; Li, Y.; Cheng, D.; Wang, R.; Chen, J.; Shen, W. Nitrite accumulation during storage of tomato fruit as prevented by hydrogen gas. Int. J. Food Prop. 2019, 22, 1425–1438. [Google Scholar] [CrossRef] [Green Version]
- Ren, P.; Jin, X.; Liao, W.; Wang, M.; Niu, L.; Li, X.; Xu, X.; Zhu, Y. Effect of hydrogen-rich water on vase life and quality in cut lily and rose flowers. Hortic. Environ. Biotechnol. 2017, 58, 576–584. [Google Scholar] [CrossRef]
- Huo, J.; Huang, D.; Zhang, J.; Fang, H.; Wang, B.; Wang, C.; Ma, Z.; Liao, W. Comparative proteomic analysis during the involvement of nitric oxide in hydrogen gas-improved postharvest freshness in cut lilies. Int. J. Mol. Sci. 2018, 19, 3955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, J.; Nie, Y.; Zhao, G.; Cheng, D.; Wang, R.; Chen, J.; Zhang, S.; Shen, W. Endogenous hydrogen gas delays petal senescence and extends the vase life of lisianthus cut flowers. Postharvest Biol. Technol. 2019, 147, 148–155. [Google Scholar] [CrossRef]
- Cai, M.; Du, H. Effects of hydrogen-rich water pretreatment on vase life of carnation (Dianthus caryophyllus) cut flowers. J. Shanghai Jiaotong Univ. 2015, 33, 41–45. [Google Scholar]
- Li, L.; Liu, Y.; Wang, S.; Zou, J.; Ding, W.; Shen, W. Magnesium hydride-mediated sustainable hydrogen supply prolongs the vase life of cut carnation flowers via hydrogen sulfide. Front. Plant Sci. 2020, 11, 595376. [Google Scholar] [CrossRef]
- Li, Y.; Li, L.; Wang, S.; Liu, Y.; Zou, J.; Ding, W.; Du, H.; Shen, W. Magnesium hydride acts as a convenient hydrogen supply to prolong the vase life of cut roses by modulating nitric oxide synthesis. Postharvest Biol. Technol. 2021, 177, 111526. [Google Scholar] [CrossRef]
- Li, L.; Lou, W.; Kong, L.; Shen, W. Hydrogen commonly applicable from medicine to agriculture: From molecular mechanisms to the field. Curr. Pharm. Des. 2021, 27, 747–759. [Google Scholar] [CrossRef]
- Temesgen, T.; Bui, T.T.; Han, M.; Kim, T.; Park, H. Micro and nanobubble technologies as a new horizon for water-treatment techniques: A review. Adv. Colloid Interface Sci. 2017, 246, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Oshita, S.; Thuyet, D.Q.; Saito, M.; Yolimoto, T. Antioxidant activity of hydrogen nanobubbles in water with different reactive oxygen species both in vivo and in vitro. Langmuir 2018, 34, 11878–11885. [Google Scholar] [CrossRef]
- Fan, W.; Zhang, Y.; Liu, S.; Li, X.; Li, J. Alleviation of copper toxicity in Daphnia magna by hydrogen nanobubble water. J. Hazard. Mater. 2020, 389, 122155. [Google Scholar] [CrossRef]
- Arora, A.; Singh, V.P. Polyols regulate the flower senescence by delaying programmed cell death in Gladiolus. J. Plant Biochem. Biotechnol. 2006, 15, 139–142. [Google Scholar] [CrossRef]
- Kobayashi, H.; Masaoka, Y.; Sato, S. Effects of excess magnesium on the growth and mineral content of rice and Echinochloa. Crop. Physiol. Ecol. 2005, 8, 38–43. [Google Scholar] [CrossRef] [Green Version]
- Bao, S.F.; Zhao, L.; Li, Z.; Cong, T. The influence of different dietary magnesium levels on the metabolism of calcium, phosphorus and magnesium in growing rats. Trace Elem. Electrolytes 2000, 17, 92–96. [Google Scholar]
- Kim, D.; Han, J. Remediation of copper contaminated soils using water containing hydrogen nanobubbles. Appl. Sci. 2020, 10, 2185. [Google Scholar] [CrossRef] [Green Version]
- Ebina, K.; Shi, K.; Hirao, M.; Hashimoto, J.; Kawato, Y.; Kaneshiro, S.; Morimoto, T.; Koizumi, K.; Yoshikawa, H. Oxygen and air nanobubble water solution promote the growth of plants, fishes, and mice. PLoS ONE 2013, 8, e65339. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, A.K.A.; Shi, X.; Hua, L.; Manzueta, L.; Qing, W.; Marhaba, T.; Zhang, W. Influences of air, oxygen, nitrogen, and carbon dioxide nanobubbles on seed germination and plant growth. J. Agric. Food Chem. 2018, 66, 5117–5124. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Wakisaka, M. Effect of air nanobubble water on the growth and metabolism of Haematococcus lacustris and Botryococcus braunii. J. Nutr. Sci. Vitaminol. 2019, 65, S212–S216. [Google Scholar] [CrossRef]
- Guo, Z.; Hu, B.; Han, H.; Lei, Z.; Shimizu, K.; Zhang, L.; Zhang, Z. Metagenomic insights into the effects of nanobubble water on the composition of gut microbiota in mice. Food Funct. 2020, 11, 7175–7182. [Google Scholar] [CrossRef]
- Su, J.; Zhang, Y.; Nie, Y.; Cheng, D.; Wang, R.; Hu, H.; Chen, J.; Zhang, J.; Du, Y.; Shen, W. Hydrogen-induced osmotic tolerance is associated with nitric oxide-mediated proline accumulation and reestablishment of redox balance in alfalfa seedlings. Environ. Exp. Bot. 2018, 147, 249–260. [Google Scholar] [CrossRef]
- Matei, N.; Camara, R.; Zhang, J.H. Emerging mechanisms and novel applications of hydrogen gas therapy. Med. Gas. Res. 2018, 8, 98–102. [Google Scholar]
- Chen, Y.; Wang, M.; Hu, L.; Liao, W.; Dawuda, M.M.; Li, C. Carbon monoxide is involved in hydrogen gas-induced adventitious root development in cucumber under simulated drought stress. Front. Plant Sci. 2017, 8, 128. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Duan, X.; Xu, S.; Wang, R.; Ouyang, Z.; Shen, W. Linking hydrogen-mediated boron toxicity tolerance with improvement of root elongation, water status and reactive oxygen species balance: A case study for rice. Ann. Bot. 2016, 118, 1279–1291. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.S.; Tahir, I. How and why of flower senescence: Understanding from models to ornamentals. Indian J. Plant Physiol. 2016, 21, 446–456. [Google Scholar] [CrossRef]
- Su, N.; Wu, Q.; Liu, Y.; Cai, J.; Shen, W.; Xia, K.; Cui, J. Hydrogen-rich water reestablishes ROS homeostasis but exerts differential effects on anthocyanin synthesis in two varieties of radish sprouts under UV-A irradiation. J. Agric. Food Chem. 2014, 62, 6454–6462. [Google Scholar] [CrossRef]
- Bowling, S.A.; Clarke, J.D.; Liu, Y.; Klessig, D.F.; Dong, X. The cpr5 mutant of Arabidopsis expresses both NPRl-dependent and NPR1-independent resistance. Pant Cell 1997, 9, 1573–1584. [Google Scholar]
- Yang, L.; Guo, A.; Wang, P. Research on the nucleic acid contents and nuclease activated for leaves of wheat albescent line during the albescent period. Acta Agric. Boreali Occident. Sin. 2001, 10, 32–35. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Zhang, Z.; Rui, Q.; Xu, L. Relationship between endopeptidase and H2O2 during the aging of wheat leaf. Acta Bot. Sin. 2001, 42, 127–131. [Google Scholar]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.; Yin, Q.; Zhang, T.; Cheng, P.; Xu, S.; Shen, W. Hydrogen Nanobubble Water Delays Petal Senescence and Prolongs the Vase Life of Cut Carnation (Dianthus caryophyllus L.) Flowers. Plants 2021, 10, 1662. https://doi.org/10.3390/plants10081662
Li L, Yin Q, Zhang T, Cheng P, Xu S, Shen W. Hydrogen Nanobubble Water Delays Petal Senescence and Prolongs the Vase Life of Cut Carnation (Dianthus caryophyllus L.) Flowers. Plants. 2021; 10(8):1662. https://doi.org/10.3390/plants10081662
Chicago/Turabian StyleLi, Longna, Qianlan Yin, Tong Zhang, Pengfei Cheng, Sheng Xu, and Wenbiao Shen. 2021. "Hydrogen Nanobubble Water Delays Petal Senescence and Prolongs the Vase Life of Cut Carnation (Dianthus caryophyllus L.) Flowers" Plants 10, no. 8: 1662. https://doi.org/10.3390/plants10081662
APA StyleLi, L., Yin, Q., Zhang, T., Cheng, P., Xu, S., & Shen, W. (2021). Hydrogen Nanobubble Water Delays Petal Senescence and Prolongs the Vase Life of Cut Carnation (Dianthus caryophyllus L.) Flowers. Plants, 10(8), 1662. https://doi.org/10.3390/plants10081662