
Morphological and Molecular Characterization of Selected Chilean Runner Bean (Phaseolus coccineus L.) Genotypes Shows Moderate Agronomic and Genetic Variability

 ,
,  and
and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Qualitative Morphological Characterization
2.2. Quantitative Morphological Characterization
2.3. Phenological Characterization
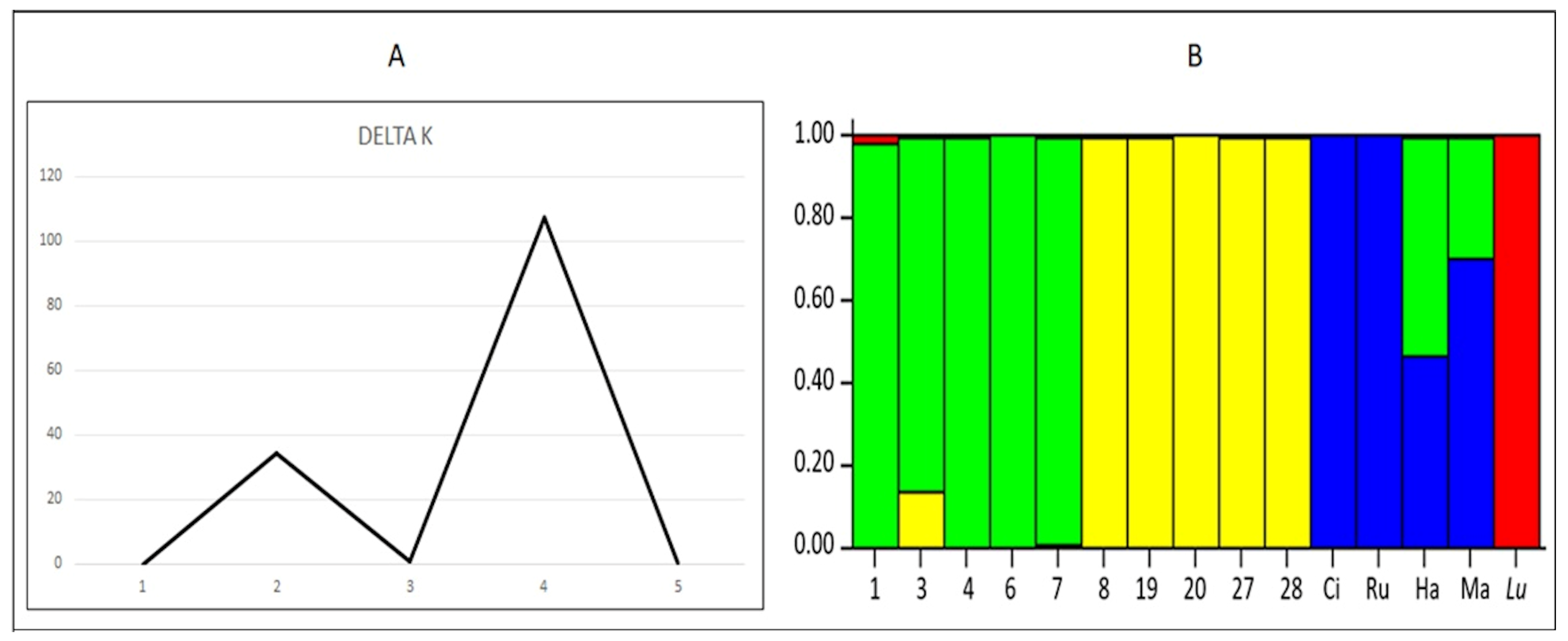
2.4. Molecular Characterization
3. Materials and Methods
3.1. Plant Material
3.2. Experiment Design and Agronomic Management
3.3. Morphological and Molecular Characterization
3.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Spataro, G.; Tiranti, B.; Arcaleni, P.; Bellucci, E.; Attene, G.; Papa, R.; Zeuli, P.S.; Negri, V. Genetic diversity and structure of a worldwide collection of Phaseolus coccineus L. Theor. Appl. Genet. 2011, 122, 1281–1291. [Google Scholar] [CrossRef] [PubMed]
- Guerra-García, A.; Suárez-Atilano, M.; Mastretta-Yanes, A.; Delgado-Salinas, A.; Piñero, D. Domestication Genomics of the Open-Pollinated Scarlet Runner Bean (Phaseolus coccineus L.). Front. Plant Sci. 2017, 8, 1891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwember, A.R.; Carrasco, B.; Gepts, P. Unraveling agronomic and genetic aspects of runner bean (Phaseolus coccineus L.). Field Crop. Res. 2017, 206, 86–94. [Google Scholar] [CrossRef]
- Łabuda, H. Runner bean (Phaseolus coccineus L.)-Biology and use. Acta Sci. Pol-Hortoru. 2010, 9, 117–132. [Google Scholar]
- Rodriguez, M.; Rau, M.; Angioi, S.A.; Bellucci, E.; Bitocchi, E.; Nanni, L.; Knüpffer, H.; Negri, V.; Papa, R.; Attene, G. European Phaseolus coccineus L. landraces: Population Structure and Adaptation, as Revealed by cpSSRs and Phenotypic Analyses. PLoS ONE 2013, 8, e57337. [Google Scholar] [CrossRef] [Green Version]
- Delgado-Salinas, A. Variation, taxonomy, domestication, and germplasm potentialities in Phaseolus coccineus. In Genetic Resources of Phaseolus Beans; Gepts, P., Ed.; Kluwer: Dordrecht, The Netherlands, 1988; pp. 441–463. [Google Scholar]
- Rodiño, A.P.; Lema, M.; Pérez-Barbeito, M.; Santalla, M.; De Ron, A.M. Assessment of runner bean (Phaseolus coccineus L.) germplasm for tolerance to low temperature during early seedling growth. Euphytica 2006, 155, 63–70. [Google Scholar] [CrossRef] [Green Version]
- Santalla, M.; Monteagudo, A.; González, A.; De Ron, A. Agronomical and quality traits of runner bean germplasm and implications for breeding. Euphytica 2004, 135, 205–215. [Google Scholar] [CrossRef]
- López-Báez, L.I.; Taboada-Gaytán, O.R.; Gil-Muñoz, A.; López, P.A.; Ortiz-Torres, E.; Vargas-Vázquez, M.; Díaz-Cervantes, R. Diversidad morfo-agronómica del frijol ayocote en el Altiplano Centro-Oriente de Puebla. Rev. Fitotec. Mex. 2018, 41, 487–497. [Google Scholar]
- Mulanya, M.M.; Kimani, P.M.; Narla, R.D. Genotypic Performance of Short-Day Runner Bean (Phaseolus coccineus L.) Lines Combining High Grain Yield and Disease Resistance. J. Agric. Sci. 2018, 10, p54. [Google Scholar] [CrossRef]
- Kimani, P.M.; Njau, S.; Mulanya, M.; Narla, R.D. Breeding runner bean for short-day adaptation, grain yield, and disease resistance in eastern Africa. Food Energy Secur. 2019, 8, 171. [Google Scholar] [CrossRef] [Green Version]
- Tay, J.; Pedreros, A.; France, A. Runner bean (Phaseolus coccineus L.) production in Chile. Annu. Rep. Bean Improv. Coop. 2008, 51, 270. [Google Scholar]
- Gilmore, B.; Myers, J.R.; Kean, D. Completion of testing of Phaseolus coccineus plant introduction (PIs) for white mold, Sclerotinia sclerotiorum resistance. Annu. Rep. Bean Improv. Coop. 2002, 45, 64–65. [Google Scholar]
- Welsh, M.; Grafton, K. Resistance to Common Bacterial Blight of Bean Introgressed from Phaseolus coccineus. HortScience 2001, 36, 750–751. [Google Scholar] [CrossRef]
- Sinkovič, L.; Pipan, B.; Vasić, M.; Antić, M.; Todorović, V.; Ivanovska, S.; Brezeanu, C.; Šuštar-Vozlič, J.; Meglič, V. Morpho-Agronomic Characterisation of Runner Bean (Phaseolus coccineus L.) from South-Eastern Europe. Sustainability 2019, 11, 6165. [Google Scholar] [CrossRef] [Green Version]
- Acampora, A.; Ciaffi, M.; De Pace, C.; Paolacci, A.R.; Tanzarella, O.A. Pattern of variation for seed size traits and molecular markers in Italian germplasm of Phaseolus coccineus L. Euphytica 2007, 157, 69–82. [Google Scholar] [CrossRef]
- Boczkowska, M.; Bulińska-Radomska, Z.; Nowosielski, J. AFLP analysis of genetic diversity in five accessions of Polish runner bean (Phaseolus coccineus L.). Genet. Resour. Crop. Evol. 2012, 59, 473–478. [Google Scholar] [CrossRef] [Green Version]
- Mercati, F.; Catarcione, G.; Paolacci, A.R.; Abenavoli, M.R.; Sunseri, F.; Ciaffi, M. Genetic diversity and population structure of an Italian landrace of runner bean (Phaseolus coccineus L.): Inferences for its safeguard and on-farm conservation. Genet. 2015, 143, 473–485. [Google Scholar] [CrossRef]
- Ruiz, R.; Mayek-Pérez, N.; Vargas-Vázquez, M.; Hernández-Delgado, S.; Muruaga-Martínez, J. Análisis De La Estructura Poblacional Del Frijol Ayocote (Phaseolus coccineus L.) Mediante Aflp. Polibotánica 2019, 47, 13–24. [Google Scholar] [CrossRef]
- Guerra-Garcia, A.; Rojas-Barrera, I.C.; Ross-Ibarra, J.; Papa, R.; Piñero, D. The genomic signature of wild-to-crop intro-gression during the domestication of scarlet runner bean (Phaseolus coccineus L.). BioRxiv 2021. [Google Scholar] [CrossRef]
- Tay, J. Quilapallar INIA: Variedad de Poroto Pallar de Excelente Calidad Culinaria [en Línea]. Chillán: Informativo INIA Quilamapu. 2013. No. 121. Available online: https://biblioteca.inia.cl/handle/123456789/4507 (accessed on 12 July 2021).
- Tay, J.; France, A.; Pedreros, A. Producción de poroto pallar. Tierra Adentro 2007, 73, 24–25. [Google Scholar]
- Wulf, K.E.; Reid, J.B.; Foo, E. What drives interspecies graft union success? Exploring the role of phylogenetic relatedness and stem anatomy. Physiol. Plant. 2020, 170, 132–147. [Google Scholar] [CrossRef]
- Gurgel, E.; Dos Santos, J.U.M.; Lucas, F.C.A.; Bastos, M.D.N.D.C. Morfologia de plântulas de Leguminosae e o potencial sistemático. Rodriguésia 2012, 63, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Laskar, R.A.; Khan, S.; Deb, C.R.; Tomlekova, N.; Wani, M.R.; Raina, A.; Amin, R. Lentil (Lens culinaris Medik.) Diversity, Cytogenetics and Breeding. In Advances in Plant Breeding Strategies: Legumes; Springer: Berlin/Heidelberg, Germany, 2019; pp. 319–369. [Google Scholar]
- Corzo-Ríos, L.J.; Sánchez-Chino, X.M.; Cardador-Martínez, A.; Martínez-Herrera, J.; Jiménez-Martínez, C. Effect of cooking on nutritional and non-nutritional compounds in two species of Phaseolus (P. vulgaris and P. coccineus) cultivated in Mexico. Int. J. Gastron. Food Sci. 2020, 20, 100206. [Google Scholar] [CrossRef]
- Sicard, D.; Nanni, L.; Porfiri, O.; Bulfon, D.; Papa, R. Genetic diversity of Phaseolus vulgaris L. and P. coccineus L. landraces in central Italy. Plant Breed. 2005, 124, 464–472. [Google Scholar] [CrossRef]
- Kwak, M.; Toro, O.; Debouck, D.G.; Gepts, P. Multiple origins of the determinate growth habit in domesticated common bean (Phaseolus vulgaris). Ann. Bot. 2012, 110, 1573–1580. [Google Scholar] [CrossRef] [Green Version]
- Repinski, S.L.; Kwak, M.; Gepts, P. The common bean growth habit gene PvTFL1y is a functional homolog of Arabidopsis TFL1. Theor. Appl. Genet. 2012, 124, 1539–1547. [Google Scholar] [CrossRef]
- Etemadi, F.; Hashemi, M.; Zandvakili, O.; Mangan, F.X. Phenology, Yield and Growth Pattern of Faba Bean Varieties. Int. J. Plant Prod. 2018, 12, 243–250. [Google Scholar] [CrossRef]
- Alvares, R.C.; Stonehouse, R.; Souza, T.L.P.O.; Melo, P.G.S.; Miklas, P.N.; Bett, K.E.; Melo, L.C.; Rodrigues, L.A.; Souza, L.L.; Pereira, H.S. Generation and validation of genetic markers for the selection of carioca dry bean genotypes with the slow-darkening seed coat trait. Euphytica 2019, 215, 1–11. [Google Scholar] [CrossRef]
- McClean, P.E.; Lee, R.K.; Otto, C.; Gepts, P.; Bassett, M.J. Molecular and phenotypic mapping of genes controlling seed coat pattern and color in common bean (Phaseolus vulgaris L.). J. Hered. 2002, 93, 148–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.; Wu, J.; Wang, L.; Blair, M.W.; Wang, S. Novel Alleles for Black and Gray Seed Color Genes in Common Bean. Crop. Sci. 2017, 57, 1603–1610. [Google Scholar] [CrossRef] [Green Version]
- McClean, P.E.; Bett, K.E.; Stonehouse, R.; Lee, R.; Pflieger, S.; Moghaddam, S.M.; Geffroy, V.; Miklas, P.; Mamidi, S. White seed color in common bean (Phaseolus vulgaris) results from convergent evolution in the P (pigment) gene. New Phytol. 2018, 219, 1112–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murube, E.; Campa, A.; Song, Q.; McClean, P.; Ferreira, J.J. Toward validation of QTLs associated with pod and seed size in common bean using two nested recombinant inbred line populations. Mol. Breed. 2019, 40, 1–17. [Google Scholar] [CrossRef]
- Vázquez, V.; Patricia, M.L.; Muruaga Martínez, J.S.; Lépiz Ildefonso, R.; Pérez Guerrero, A. La colección INIFAP de frijol ayocote (Phaseolus coccineus L.) I: Distribución geográfica de sitios de colecta. Rev. Mex. Cienc. Agr. 2012, 3, 1247–1259. [Google Scholar]
- Hamburdă, S.; Munteanu, N.; Stoleru, V.; Teliban, G.; Butnariu, G.; Popa, L. Evaluation of the Possibilities of Using Runner Bean (Phaseolus coccineus L.) in Landscaping Design. 2007. Available online: https://www.uaiasi.ro/revista_horti/files/Nr1_2014/Vol%2057_1_2014(13) (accessed on 27 June 2021).
- Oyanagi, A.; Nakamoto, T.; Morita, S. The gravitropic response of roots and the shaping of the root system in cereal plants. Environ. Exp. Bot. 1993, 33, 141–158. [Google Scholar] [CrossRef]
- Bocianowski, J.; Łuczkiewicz, T.; Kozak, M.; Brzeskwiniewicz, H. Does Genetic Distance of Parental Forms Affect Specific Combining Ability and Heterosis Effects in F1 and F2 of Spring Rape Diallel Cross? Agric. Conspec. Sci. 2008, 73, 211–215. [Google Scholar]
- Meza-Vázquez, K.E.; Lépiz-Ildefonso, R.; López-Alcocer, J.D.J.; Morales-Rivera, M.M. Caracterización morfológica y fenológica de especies silvestres de frijol (Phaseolus). Rev. Fitotec. Mex. 2015, 38, 17–28. [Google Scholar] [CrossRef]
- Pando, J.B.; Fohouo, F.-N.T.; Tamesse, J.L. Foraging and pollination behaviour of Xylocopa calens Lepeletier (Hymenoptera: Apidae) on Phaseolus coccineus L. (Fabaceae) flowers at Yaounde (Cameroon). Entomol. Res. 2011, 41, 185–193. [Google Scholar] [CrossRef]
- Nechifor, B.; Filimon, R.; Szilagyi, L. Genetic variability, heritability and expected genetic advance as indices for yield and yield components selection in common bean (Phaseolus vulgaris L.). Sci. Pap. UASVM Buchar. Ser. A 2011, 54, 332. [Google Scholar]
- Mhlaba, Z.B.; Shimelis, H.; Amelework, B.; Modi, A.T.; Mashilo, J. Variance components and heritability of yield and yield-related traits in tepary bean (Phaseolus acutifolius). S. Afr. J. Plant Soil 2019, 36, 117–128. [Google Scholar] [CrossRef]
- Giurcă, D.M. Morphological and phenological differences between the two species of the Phaseolus genus (Phaseolus vulgaris and Phaseolus coccineus). Cercet. Agron. În Mold. 2009, 42, 39–45. [Google Scholar]
- Bezerra, A.C.; Neto, F.A.; Das Neves, A.C.; Maggioni, K. Comportamento morfoagronômico de feijão-caupi, cv. BRS Guariba, sob diferentes densidades de plantas. Rev. de Ciencias Agrárias 2012, 55, 184–189. [Google Scholar] [CrossRef]
- Sinkovič, L.; Pipan, B.; Sinkovič, E.; Meglič, V. Morphological Seed Characterization of Common (Phaseolus vulgaris L.) and Runner (Phaseolus coccineus L.) Bean Germplasm: A Slovenian Gene Bank Example. BioMed Res. Int. 2019, 2019, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nei, M. Analysis of Gene Diversity in Subdivided Populations. Proc. Natl. Acad. Sci. USA 1973, 70, 3321–3323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, S.N. Uso adecuado de fertilizantes en porotos, Zona centro sur. Bol. INIA—Inst. De Investig. Ag-Ropecuarias 2004, 116, 1–26. Available online: https://biblioteca.inia.cl/handle/123456789/7028 (accessed on 3 June 2021).
- Doyle, J.J.; Doyle, J.L. A rapid total DNA preparation procedure for fresh plant tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Carrasco, B.; Garcés, M.; Rojas, P.; Saud, G.; Herrera, R.; Retamales, J.B.; Caligari, P.D. The Chilean Strawberry [Fragaria chiloensis (L.) Duch.]: Genetic Diversity and Structure. J. Am. Soc. Hortic. Sci. 2007, 132, 501–506. [Google Scholar] [CrossRef] [Green Version]
- Hammer, Ø.; Harper, D.; Ryan, P. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 4–9. [Google Scholar]
- Di Rienzo, J.; Casanoves, F.; Balzarini, M.; Gonzalez, L.; Tablada, M.; Robledo, C. InfoStat versión 2016. Córdoba, InfoStat Group, Facultad de Ciencias Agropecuarias, Universidad Nacional de Córdoba. 2016. Available online: http://www.infostat.com.ar (accessed on 23 June 2021).
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Yeh, F.C.; Yang, R.C.; Boyle, T.B.; Ye, Z.H.; Mao, J.X. POPGENE, the user-friendly shareware for population genetic analysis. Molecular biology and biotechnology centre. Popul. Genet. Anal. 1997, 10, 295–301. [Google Scholar]
- Nei, M. Estimation of Average Heterozygosity and Genetic Distance from A Small Number of Individuals. Genetics 1978, 89, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Stephens, M.; Rosenberg, N.; Donnelly, P. Association Mapping in Structured Populations. Am. J. Hum. Genet. 2000, 67, 170–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evanno:, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lines | Germination Type | Color of Flower | Branch Type | Growth Habit |
|---|---|---|---|---|
| 1 | Hypogeal | White | Evenly distributed (upper and lower) | Indeterminate |
| 3 | Epigeal | Purple | Evenly distributed/lower * | Indeterminate/determinate * |
| 4 | Epigeal | White | Evenly distributed | Indeterminate |
| 6 | Epigeal | White | Evenly distributed | Indeterminate |
| 7 | Epigeal | White | Evenly distributed | Indeterminate |
| 8 | Hypogeal | White | Evenly distributed | Indeterminate |
| 19 | Hypogeal | Orange | Evenly distributed | Indeterminate |
| 20 | Hypogeal | White | Evenly distributed | Indeterminate |
| 27 | Hypogeal | Orange/White | Evenly distributed/upper * | Indeterminate |
| 28 | Hypogeal | Orange/White | Evenly distributed/upper * | Indeterminate |
| Pod-Related Traits | Seed-Related Traits | Root-Related Traits | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Lines | Color | Shape | Fibers | Color | Longitudinal Cut | Cross-Section Cut | Hilum Color | Color | Nodules | Type |
| 1 | Beige | Concave | Yes | White | Kidney | Narrow elliptical | White | Brown | Yes | Fibrous |
| 3 | Beige/Purple spots | Concave | Yes | Cream/Black spots | Kidney | Narrow elliptical | Cream | Brown | No | Fibrous |
| 4 | Cream/Brown spots | Concave | Yes | White | Kidney | Narrow elliptical | White | Brown | No | Fibrous |
| 6 | Cream | Concave | Yes | White | Elliptical Circular | Narrow elliptical | White | Brown | Yes | Fibrous |
| 7 | Beige | Concave | Yes | White | Elliptical Circular | Narrow elliptical | White | Brown | Yes | Fibrous |
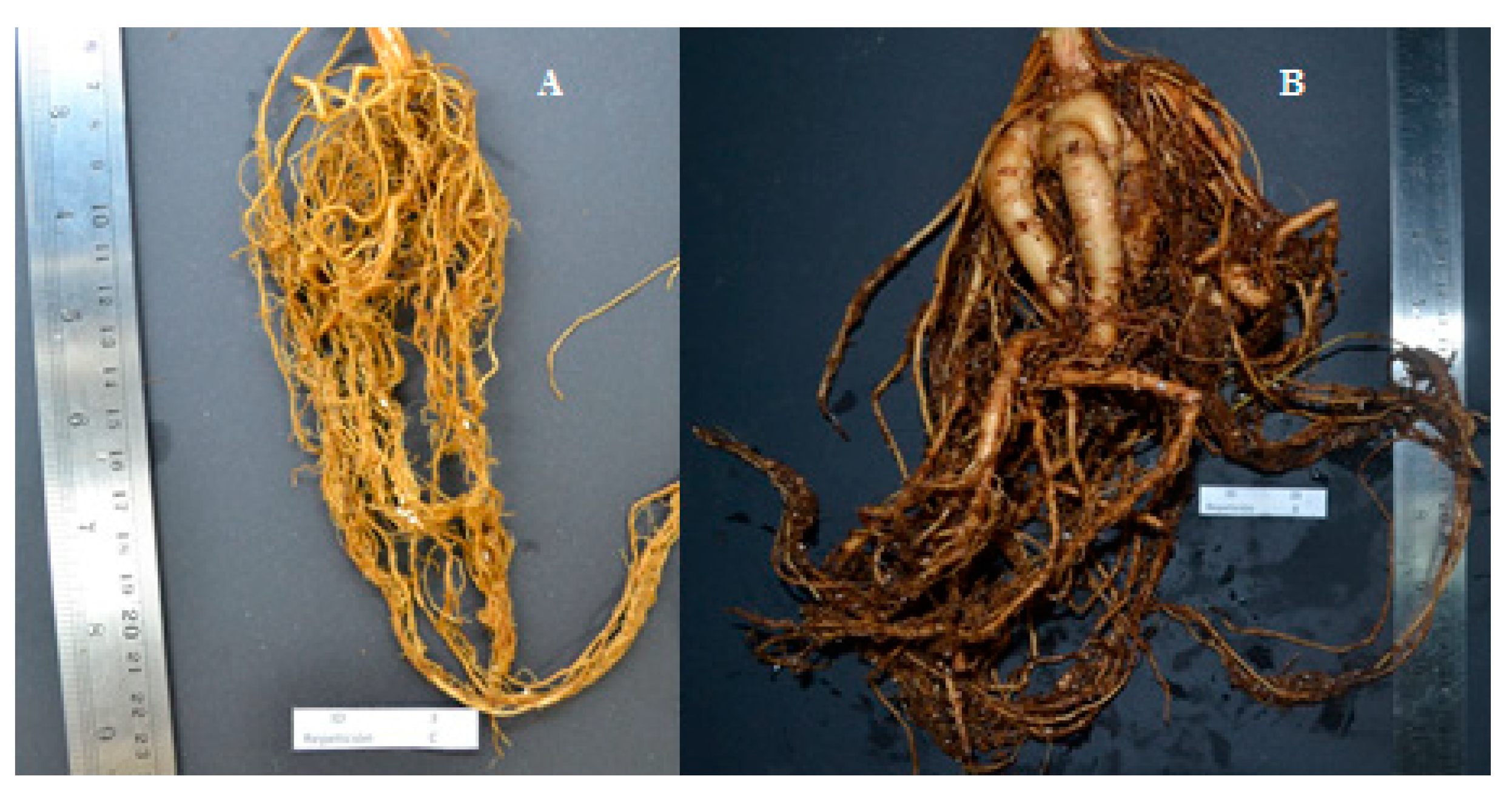
| 8 | Beige | Concave | Yes | White | Kidney | Elliptical | White | Cream | Yes | Tuberous |
| 19 | Beige | Concave | Yes | Purple/Black spots | Kidney | Circular | Cream | Cream | Yes | Tuberous |
| 20 | Beige | Concave | Yes | White | Rectangular | Elliptical | White | White | Yes | Tuberous |
| 27 | Beige | Concave | Yes | Purple/Black spots | Kidney | Elliptical | White | Cream | Yes | Tuberous |
| 28 | Beige | Concave | Yes | Purple/Black spots | Kidney | Elliptical | Cream | Cream | Yes | Tuberous |
| Trait | Sum of Squares | d.f. | Mean Squares | p Value | GDG |
|---|---|---|---|---|---|
| Days to V2 | 1429.280 | 9 | 158.809 | 0.007 | 55.85% |
| N° of seeds per pod | 42.600 | 9 | 8.088 | 0.001 | 67.83% |
| Pod length | 142.424 | 9 | 7.162 | 0.007 | 64.46% |
| Beak length | 18.674 | 9 | 2.074 | 3.69 × 10−16 | 95.04% |
| Seed width | 0.535 | 9 | 0.059 | 0.004 | 72.15% |
| Pod-Related Traits | Seed-Related Traits | |||||||
|---|---|---|---|---|---|---|---|---|
| Lines | N° of Seeds/Pod | Length (cm) | Length of Beak (cm) | Width (cm) | N° of Pods/Plant | Length (cm) | Width (cm) | Weight of 10 Seeds |
| 1 | 3.26 b | 13.46 ab | 1.76 ab | 1.5 | 35.67 | 1.95 | 1.08 bc | 8.5 |
| 3 | 4.92 a | 16.48 a | 1.49 b | 1.66 | 16.17 | 1.94 | 1.07 bc | 8.68 |
| 4 | 2.87 b | 11.76 bc | 2.06 a | 1.45 | 46.17 | 1.89 | 1.06 c | 8.07 |
| 6 | 2.78 b | 12.20 bc | 1.86 ab | 1.28 | 26.83 | 1.91 | 1.07 bc | 8.88 |
| 7 | 3.13 b | 12.59 bc | 1.70 b | 1.39 | 26.57 | 1.92 | 1.05 c | 8.54 |
| 8 | 2.19 b | 10.47 bc | 0.58 c | 1.53 | 3 | 2.06 | 1.23 abc | 13.55 |
| 19 | 1.77 b | 10.75 c | 0.52 c | 1.82 | 6 | 2.24 | 1.29 a | 15.46 |
| 20 | 2.02 b | 10.34 c | 0.59 c | 1.78 | 4 | 1.98 | 1.25 ab | 11.65 |
| 27 | 1.85 b | 11.84 bc | 0.69 c | 1.77 | 8.71 | 2.24 | 1.34 a | 14.4 |
| 28 | 2.50 b | 12.87 bc | 0.70 c | 1.87 | 2.4 | 2.07 | 1.23 abc | 11.89 |
| Species | Region | N | Na | Ne | I | h | P |
|---|---|---|---|---|---|---|---|
| P. coccineus | Central of Chile | 6 | 1.652 ± 0.477 | 1.399 ± 0.334 | 0.319 ± 0.262 | 0.208 ± 0.182 | 65.23% |
| Southern of Chile | 4 | 1.582 ± 0.494 | 1.366 ± 0.361 | 0.322 ± 0.286 | 0.216 ± 0.198 | 58.20% | |
| P. coccineus | Overall | 10 | 1.793 ± 0.406 | 1.378 ± 0.314 | 0.367 ± 0.237 | 0.237 ± 0.169 | 79.30% |
| P. vulgaris + P. lunatus | 5 | 1.656 ± 0.475 | 1.357 ± 0.358 | 0.327 ± 0.266 | 0.214 ± 0.187 | 65.62% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arriagada, O.; Schwember, A.R.; Greve, M.J.; Urban, M.O.; Cabeza, R.A.; Carrasco, B. Morphological and Molecular Characterization of Selected Chilean Runner Bean (Phaseolus coccineus L.) Genotypes Shows Moderate Agronomic and Genetic Variability. Plants 2021, 10, 1688. https://doi.org/10.3390/plants10081688
Arriagada O, Schwember AR, Greve MJ, Urban MO, Cabeza RA, Carrasco B. Morphological and Molecular Characterization of Selected Chilean Runner Bean (Phaseolus coccineus L.) Genotypes Shows Moderate Agronomic and Genetic Variability. Plants. 2021; 10(8):1688. https://doi.org/10.3390/plants10081688
Chicago/Turabian StyleArriagada, Osvin, Andrés R. Schwember, María Jesús Greve, Milan O. Urban, Ricardo A. Cabeza, and Basilio Carrasco. 2021. "Morphological and Molecular Characterization of Selected Chilean Runner Bean (Phaseolus coccineus L.) Genotypes Shows Moderate Agronomic and Genetic Variability" Plants 10, no. 8: 1688. https://doi.org/10.3390/plants10081688
APA StyleArriagada, O., Schwember, A. R., Greve, M. J., Urban, M. O., Cabeza, R. A., & Carrasco, B. (2021). Morphological and Molecular Characterization of Selected Chilean Runner Bean (Phaseolus coccineus L.) Genotypes Shows Moderate Agronomic and Genetic Variability. Plants, 10(8), 1688. https://doi.org/10.3390/plants10081688