
A Core35S Promoter of Cauliflower Mosaic Virus Drives More Efficient Replication of Turnip Crinkle Virus



{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
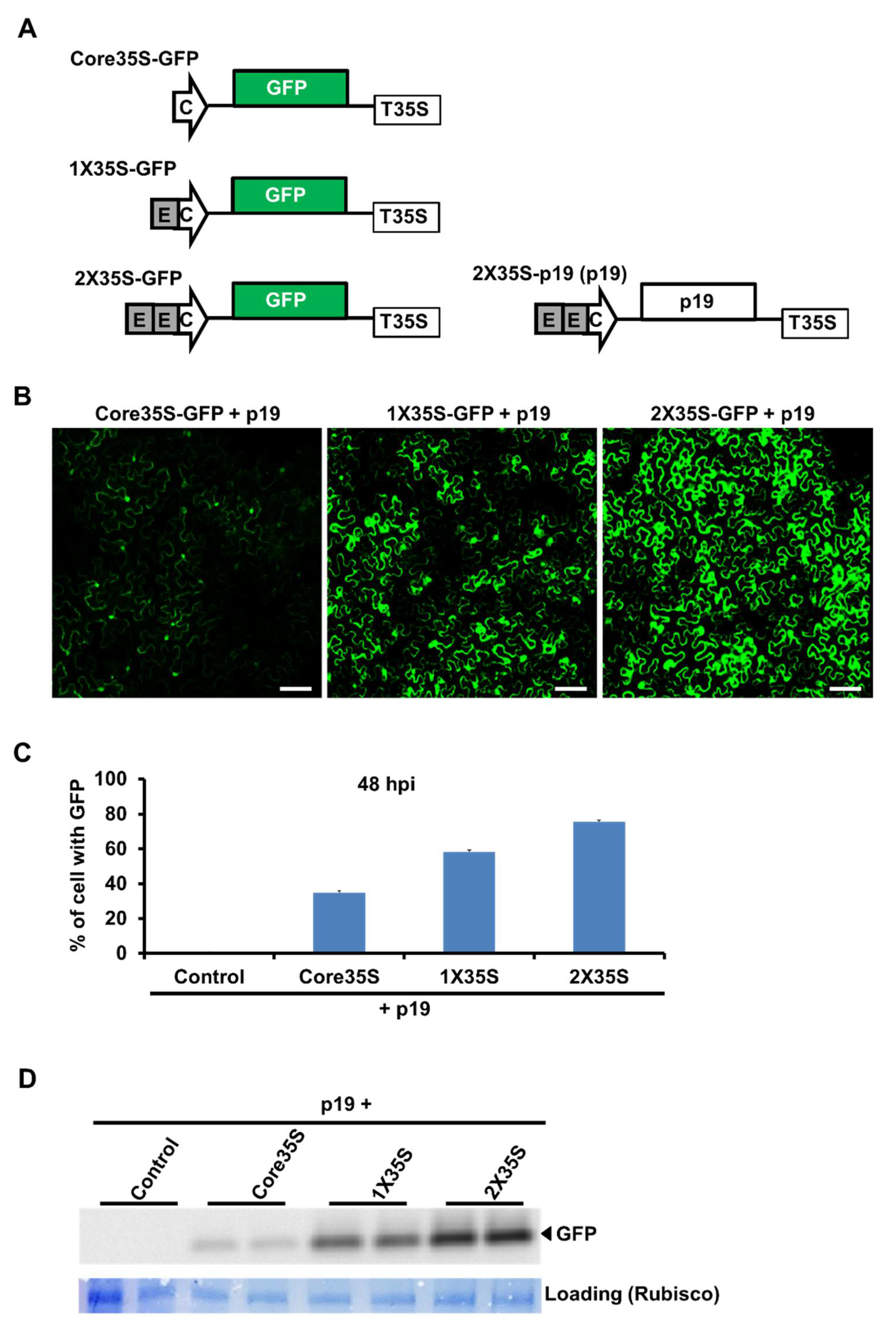
2.1. Strength Variation of Three Variants of 35S Promoters by GFP Expression
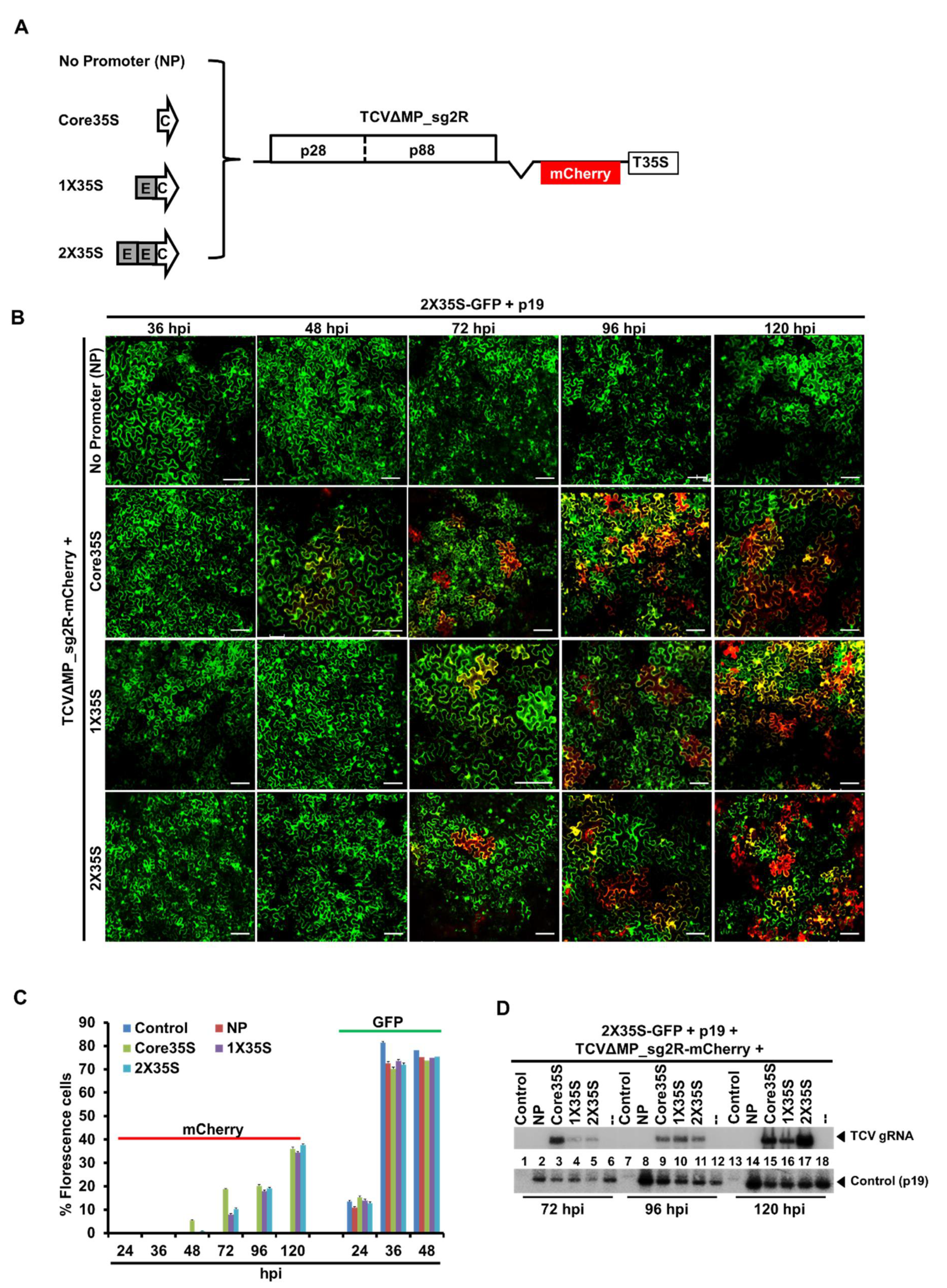
2.2. TCV-mCherry Replicon Is Variably Driven by the Three Promoters of Distinctive Strengths
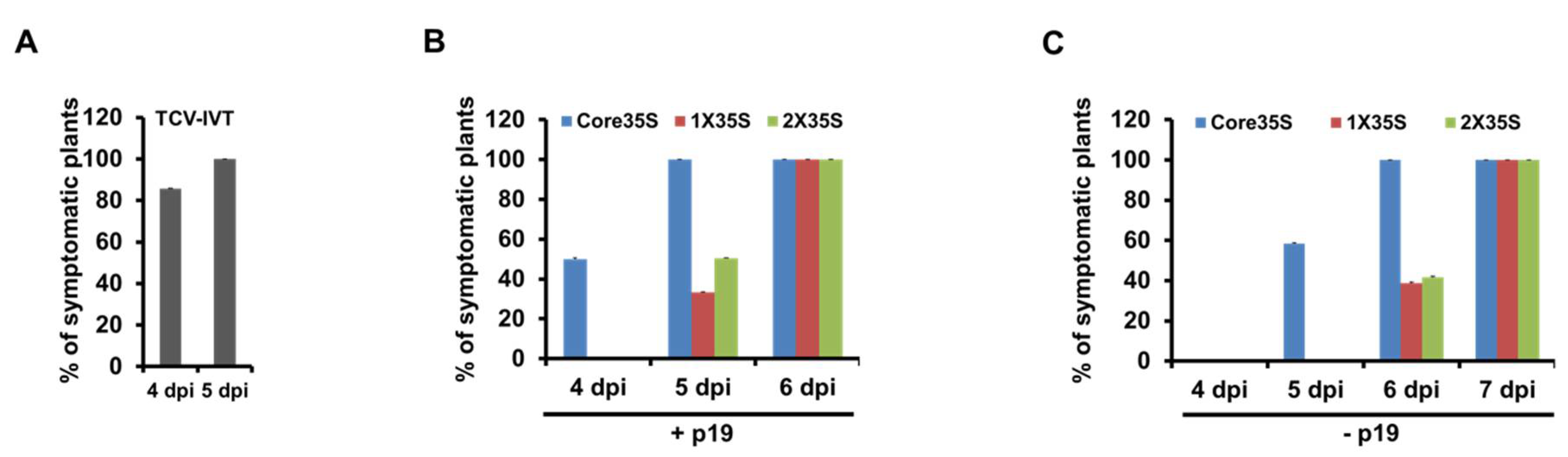
2.3. Symptom Expression Comparison of the Wildtype TCV cDNA by 35S Promoter Variants with In Vitro Transcript
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plant Materials
5.2. Plasmid Construction
5.3. Agroinfiltration of N. benthamiana Plants
5.4. Infection of N. benthamiana Plants with In Vitro Transcripts
5.5. RNA Blot Analysis
5.6. Western Blot Analysis
5.7. Confocal Microscopy
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Koziel, M.G.; Carozzi, N.B.; Desai, N. Optimizing expression of transgenes with an emphasis on post-transcriptional events. Plant Mol. Biol. 1996, 32, 393–405. [Google Scholar] [CrossRef] [PubMed]
- Potenza, C.; Aleman, L.; Sengupta-Gopalan, C. Targeting transgene expression in research, agricultural, and environmental applications: Promoters used in plant transformation. In Vitro Cell. Dev. Biol. Plant 2004, 40. [Google Scholar] [CrossRef]
- Juven-Gershon, T.; Kadonaga, J.T. Regulation of gene expression via the core promoter and the basal transcriptional machinery. Dev. Biol. 2010, 339, 225–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katagiri, F.; Chua, N.-H. Plant transcription factors: Present knowledge and future challenges. Trends Genet. 1992, 8, 22–27. [Google Scholar] [CrossRef]
- Porto, M.S.; Pinheiro, M.P.N.; Batista, V.G.L.; dos Santos, R.C.; de Albuquerque Melo Filho, P.; de Lima, L.M. Plant promoters: An approach of structure and function. Mol. Biotechnol. 2014, 56, 38–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guilley, H.; Dudley, R.K.; Jonard, G.; Balàzs, E.; Richards, K.E. Transcription of cauliflower mosaic virus DNA: Detection of promoter sequences, and characterization of transcripts. Cell 1982, 30, 763–773. [Google Scholar] [CrossRef]
- Benfey, P.N.; Chua, N.-H. The Cauliflower Mosaic Virus 35S Promoter: Combinatorial Regulation of Transcription in Plants. Science 1990, 250, 959–966. [Google Scholar] [CrossRef] [Green Version]
- Kay, R.; Chan, A.; Daly, M.; Mcpherson, J. Duplication of CaMV 35S Promoter Sequences Creates a Strong Enhancer for Plant Genes. Science 1987, 236, 1299–1302. [Google Scholar] [CrossRef]
- Odell, J.T.; Nagy, F.; Chua, N.-H. Identification of DNA sequences required for activity of the cauliflower mosaic virus 35S promoter. Nat. Cell Biol. 1985, 313, 810–812. [Google Scholar] [CrossRef]
- Martínez, F.; Sardanyés, J.; Elena, S.F.; Daròs, J.-A. Dynamics of a plant RNA virus intracellular accumulation: Stamping machine vs. geometric replication. Genetics 2011, 188, 637–646. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Ma, X.; Qian, S.; Zhou, X.; Sun, K.; Chen, X.; Zhou, X.; Jackson, A.O.; Li, Z. Rescue of a Plant Negative-Strand RNA Virus from Cloned cDNA: Insights into Enveloped Plant Virus Movement and Morphogenesis. PLoS Pathog. 2015, 11, e1005223. [Google Scholar] [CrossRef]
- Carrington, J.C.; Heaton, L.A.; Zuidema, D.; Hillman, B.I.; Morris, T. The genome structure of turnip crinkle virus. Virology 1989, 170, 219–226. [Google Scholar] [CrossRef]
- Qu, F.; Ren, T.; Morris, T.J. The Coat Protein of Turnip Crinkle Virus Suppresses Posttranscriptional Gene Silencing at an Early Initiation Step. J. Virol. 2003, 77, 511–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hacker, D.; Petty, I.; Wei, N.; Morris, T. Turnip crinkle virus genes required for RNA replication and virus movement. Virology 1992, 186. [Google Scholar] [CrossRef]
- Schnurr, J.A.; Guerra, D.J. The CaMV-35S promoter is sensitive to shortened photoperiod in transgenic tobacco. Plant Cell Rep. 2000, 19, 279–282. [Google Scholar] [CrossRef] [PubMed]
- Pret’Ová, A.; Obert, B.; Wetzstein, H. Leaf developmental stage and tissue location affect the detection of β-glucuronidase in transgenic tobacco plants. Biotechnol. Lett. 2001, 23, 555–558. [Google Scholar] [CrossRef]
- Driesen, M.; Benito-Moreno, R.-M.; Hohn, T.; Fütterer, J. Transcription from the CaMV 19 S Promoter and Autocatalysis of Translation from CaMV RNA. Virology 1993, 195, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Medberry, S.L.; Lockhart, B.; Olszewski, N.E. The Commelina yellow mottle virus promoter is a strong promoter in vascular and reproductive tissues. Plant Cell 1992, 4, 185–192. [Google Scholar]
- Medberry, S.L.; Olszewski, N.E. Identification of cis elements involved in Commelina yellow mottle virus promoter activity. Plant J. 1993, 3, 619–626. [Google Scholar] [CrossRef]
- Chen, G.; Müller, M.; Potrykus, I.; Hohn, T.; Fütterer, J. Rice tungro bacilliform virus: Transcription and translation in protoplasts. Virology 1994, 204, 91–100. [Google Scholar] [CrossRef]
- Lindbo, J.A. High-efficiency protein expression in plants from agroinfection-compatible Tobacco mosaic virus expression vectors. BMC Biotechnol. 2007, 7, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morilla, G.; Castillo, A.G.; Preiss, W.; Jeske, H.; Bejarano, E.R. A Versatile Transreplication-Based System To Identify Cellular Proteins Involved in Geminivirus Replication. J. Virol. 2006, 80, 3624–3633. [Google Scholar] [CrossRef] [Green Version]
- Meng, B.; Venkataraman, S.; Li, C.; Wang, W.; Dayan-Glick, C.; Mawassi, M. Construction and biological activities of the first infectious cDNA clones of the genus Foveavirus. Virology 2013, 435, 453–462. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.-Y.; Liu, K.-H.; Wang, Y.-C.; Wu, J.-F.; Chiu, W.-L.; Chen, C.-Y.; Wu, S.-H.; Sheen, J.; Lai, E.-M. AGROBEST: An efficient Agrobacterium-mediated transient expression method for versatile gene function analyses in Arabidopsis seedlings. Plant Methods 2014, 10, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Yang, Y.; Wang, X.; Yu, F.; Yu, C.; Chen, J.; Cheng, Y.; Yan, C.; Chen, J. Enhanced transgene expression in rice following selection controlled by weak promoters. BMC Biotechnol. 2013, 13, 29. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, A.; Garcia, H.G.; Jones, D.; Phillips, R.; Kondev, J. Effect of Promoter Architecture on the Cell-to-Cell Variability in Gene Expression. PLoS Comput. Biol. 2011, 7, e1001100. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-F.; Sun, R.; Guo, Q.; Zhang, S.; Meulia, T.; Halfmann, R.; Li, D.; Qu, F. A self-perpetuating repressive state of a viral replication protein blocks superinfection by the same virus. PLoS Pathog. 2017, 13, e1006253. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Sun, R.; Guo, Q.; Zhang, X.-F.; Qu, F. Repression of turnip crinkle virus replication by its replication protein p88. Virology 2019, 526, 165–172. [Google Scholar] [CrossRef]
- Lin, J.; Guo, J.; Finer, J.; Dorrance, A.E.; Redinbaugh, M.G.; Qu, F. The Bean pod mottle virus RNA2-encoded 58-kilodalton protein P58 is required in cis for RNA2 accumulation. J. Virol. 2014, 88, 3213–3222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Zhang, X.; Singh, J.; Li, D.; Qu, F. Temperature-Dependent Survival of Turnip Crinkle Virus-Infected Arabidopsis Plants Relies on an RNA Silencing-Based Defense That Requires DCL2, AGO2, and HEN1. J. Virol. 2012, 86, 6847–6854. [Google Scholar] [CrossRef] [Green Version]
- Cao, M.; Ye, X.; Willie, K.; Lin, J.; Zhang, X.; Redinbaugh, M.G.; Simon, A.E.; Morris, T.J.; Qu, F. The Capsid Protein of Turnip Crinkle Virus Overcomes Two Separate Defense Barriers to Facilitate Systemic Movement of the Virus in Arabidopsis. J. Virol. 2010, 84, 7793–7802. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, M.E.; Waliullah, S. A Core35S Promoter of Cauliflower Mosaic Virus Drives More Efficient Replication of Turnip Crinkle Virus. Plants 2021, 10, 1700. https://doi.org/10.3390/plants10081700
Ali ME, Waliullah S. A Core35S Promoter of Cauliflower Mosaic Virus Drives More Efficient Replication of Turnip Crinkle Virus. Plants. 2021; 10(8):1700. https://doi.org/10.3390/plants10081700
Chicago/Turabian StyleAli, Md Emran, and Sumyya Waliullah. 2021. "A Core35S Promoter of Cauliflower Mosaic Virus Drives More Efficient Replication of Turnip Crinkle Virus" Plants 10, no. 8: 1700. https://doi.org/10.3390/plants10081700
APA StyleAli, M. E., & Waliullah, S. (2021). A Core35S Promoter of Cauliflower Mosaic Virus Drives More Efficient Replication of Turnip Crinkle Virus. Plants, 10(8), 1700. https://doi.org/10.3390/plants10081700