
Chemical Diversity and Therapeutic Effects of Essential Oils of Aniba Species from the Amazon: A Review

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Economic and Traditional Uses and Conservation of Aniba Species
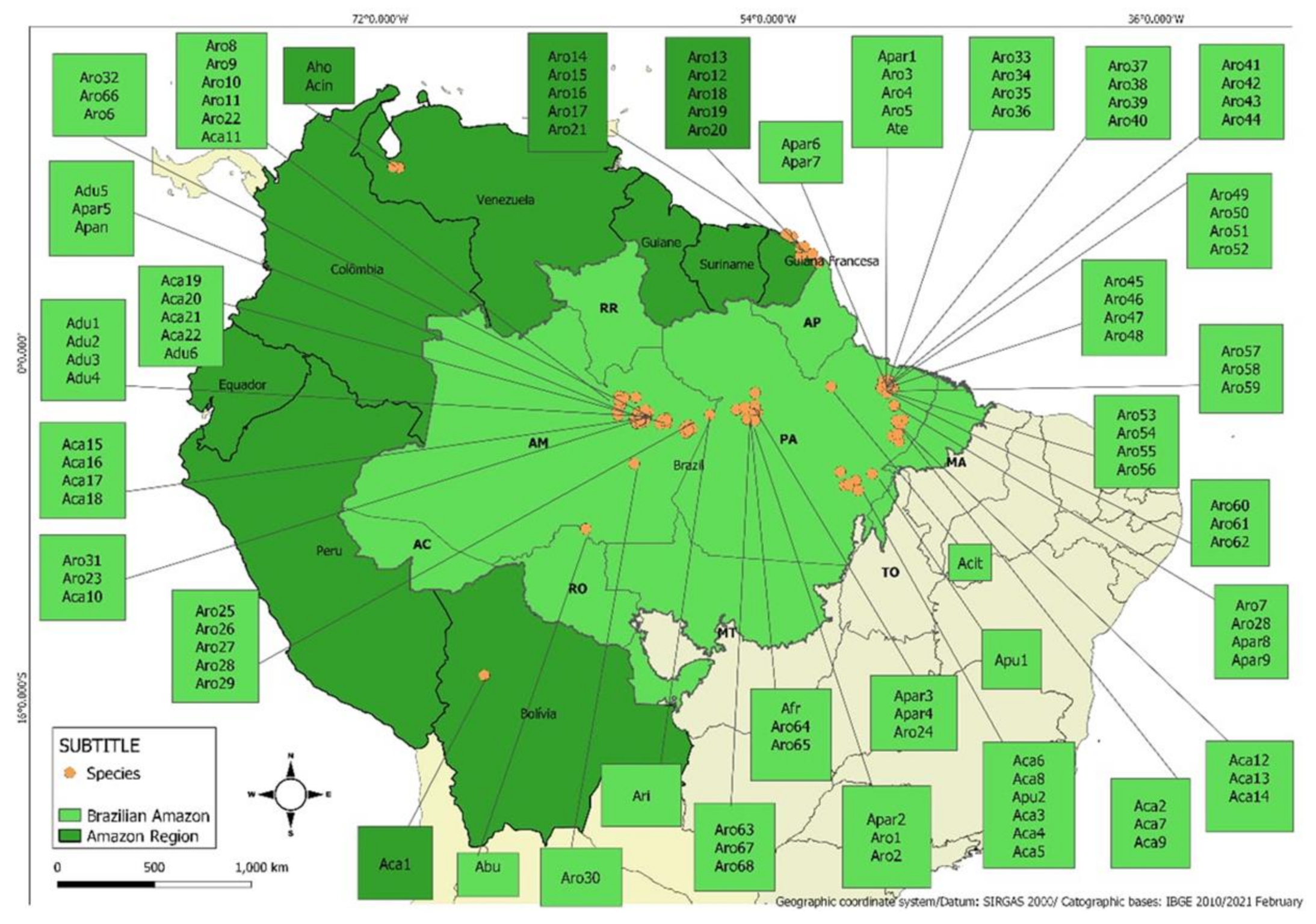
3. Scope of Collected Data
4. Multivariate Statistical Analysis Based on the Essential Oils of Aniba Species
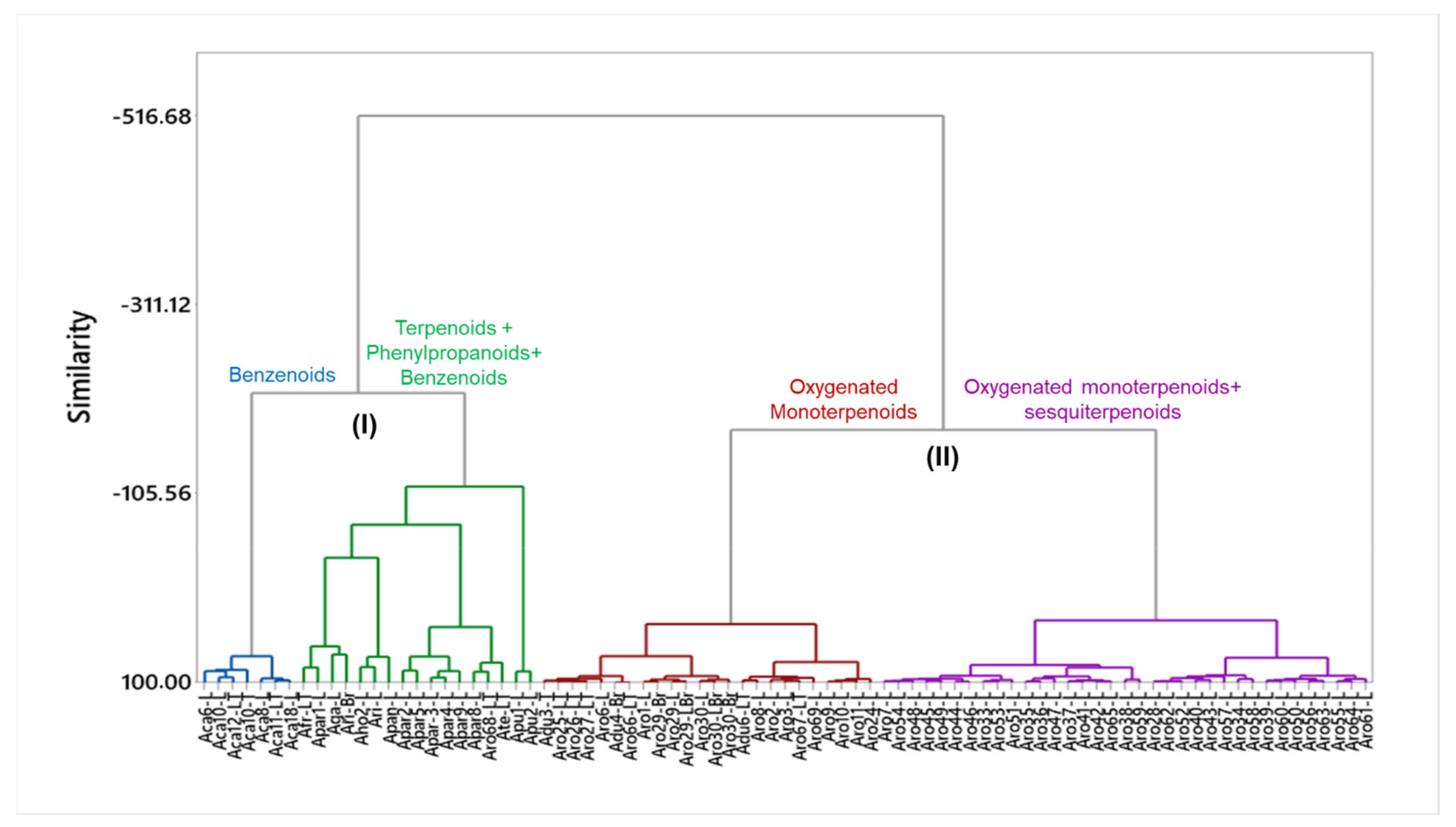
4.1. Essential Oils from Leaves, Thin Twigs and Branches of Aniba Species
4.1.1. Cluster I: Benzenoid-Rich Oils
4.1.2. Cluster I: Terpenoid, Phenylpropanoid and Benzenoid-Rich Oils
4.1.3. Cluster II: Oxygenated Monoterpene-Rich Oils
4.1.4. Cluster II: Oils Rich in Oxygenated Mono- and Sesquiterpenes
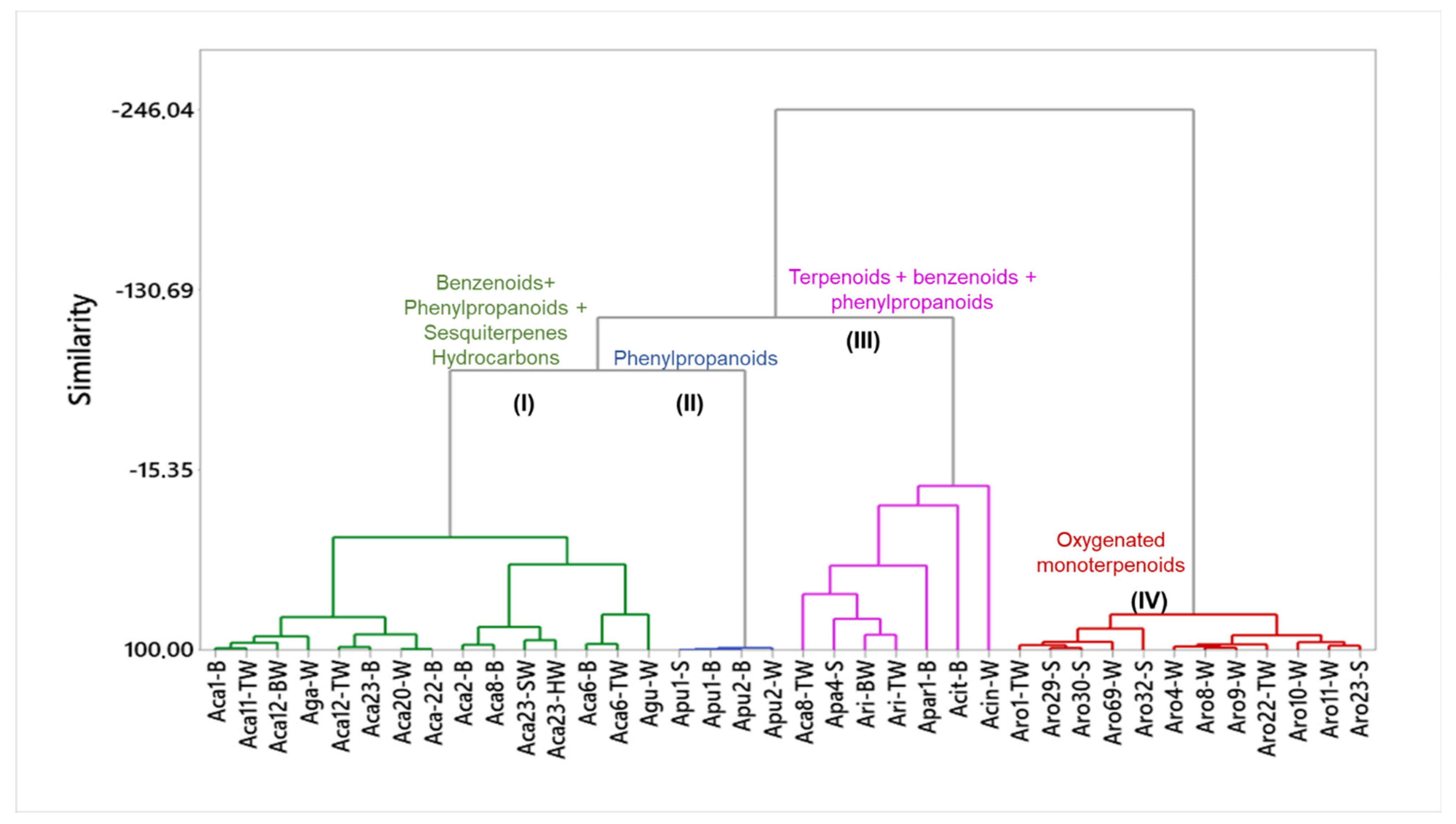
4.2. Essential Oils from Stem, Bark and Trunkwood of Aniba Species
4.2.1. Cluster I: Oils Rich in Benzenoids, Phenylpropanoids and Sesquiterpene Hydrocarbons
4.2.2. Cluster II: Phenylpropanoid-Rich Oils
4.2.3. Cluster III: Oils Rich in Terpenoids, Benzenoids and Phenylpropanoids
4.2.4. Cluster IV: Oxygenated Monoterpenoid-Rich Oils
5. Aniba Commercial Samples
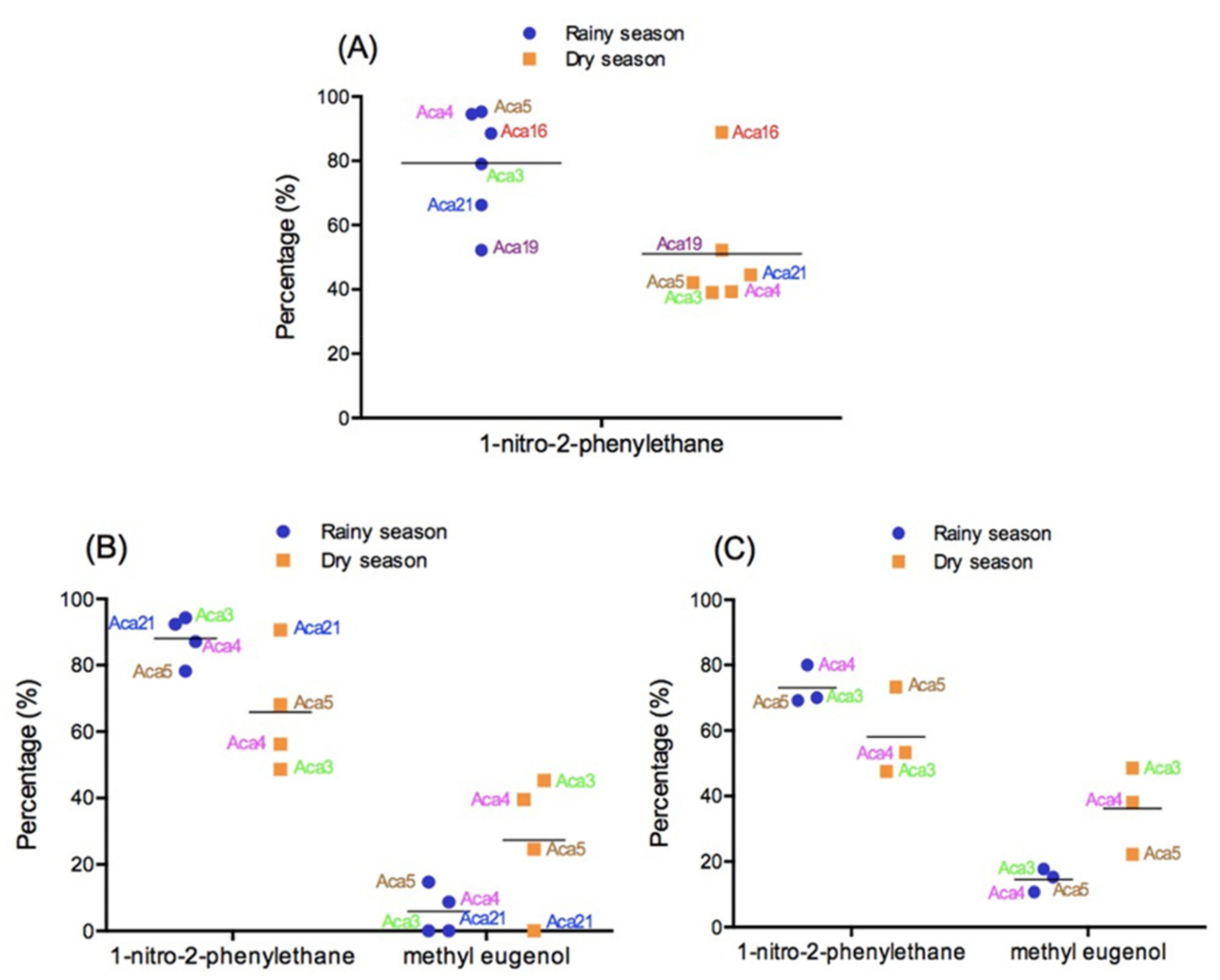
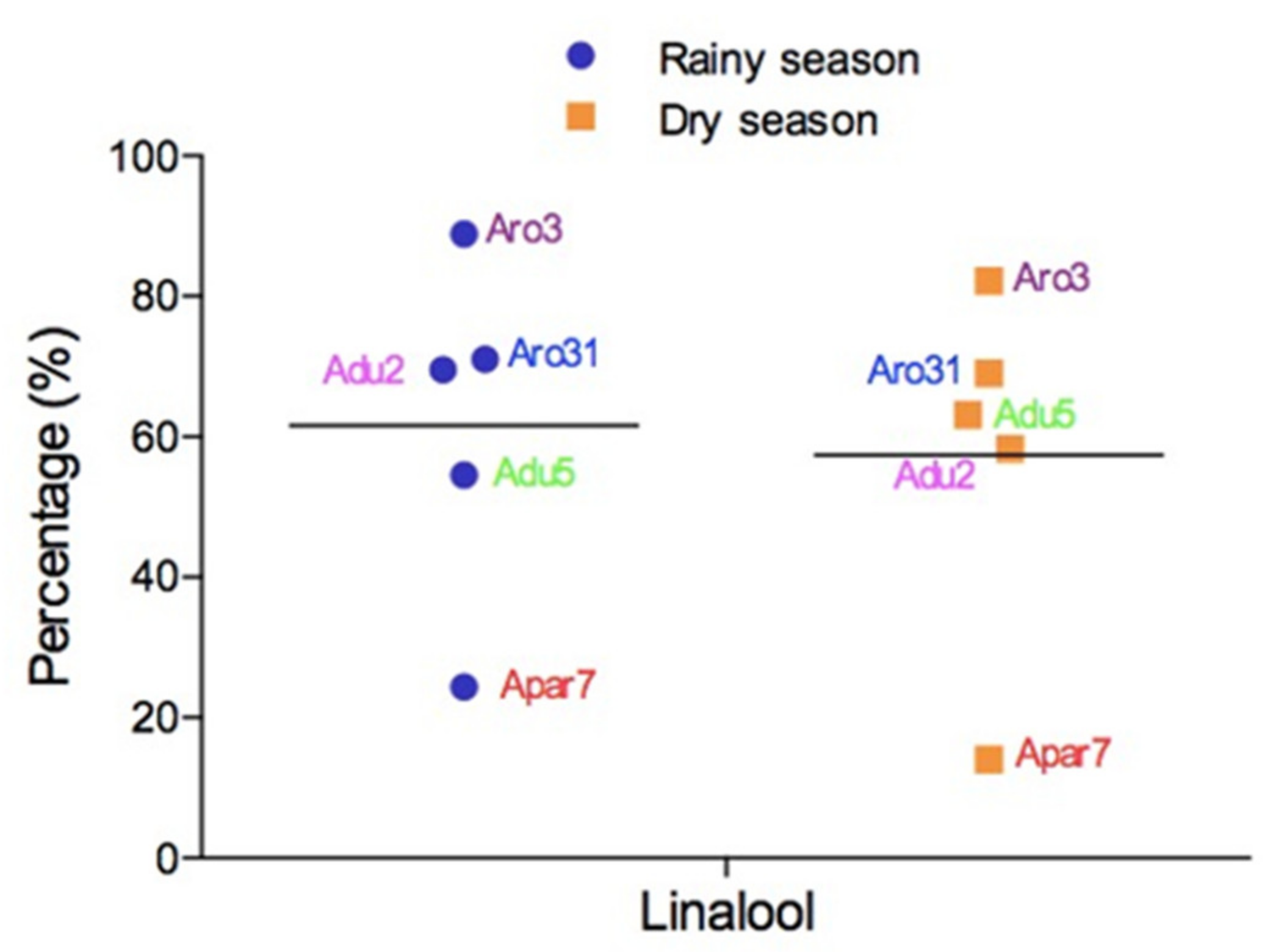
6. Seasonal Variation in the Aniba Volatile Constituents

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Collection Site | Plant Part | Extraction Type | Major Components | References |
|---|---|---|---|---|---|
| A. burchellii | Humaitá, AM, Brazil | Leaf | SD | Benzyl salicylate (45.6%), α-pinene (12,1%), and benzyl benzoate (5.2%) | [106] |
| A. canelilla | Manaus, AM, Brazil | Sapwood | HD | Methyleugenol (65.3%) and eugenol (34.4%) | [12] |
| A. canelilla | Manaus, AM, Brazil | Heartwood | HD | 1-Nitro-2-phenylethane (46.0%), methyleugenol (52.9%), and linalool (5.0%) | [12] |
| A. canelilla | Manaus, AM, Brazil | Bark | HD | 1-Nitro-2-phenylethane (72.6%) and methyleugenol (24.9%) | [12] |
| A. canelilla | Fatima de Chimanes, Bolivia | Bark | HD | 1-Nitro-2-phenylethane (89.8%), methyleugenol (2.90%), and safrole (2.6%) | [83] |
| A. canelilla | Carajás National Forest, Marabá, PA, Brazil | Leaf | not reported | 1-Nitro-2-phenylethane (95.3%) and limonene (1.2%) | [64] |
| A. canelilla | Carajás National Forest, Marabá, PA, Brazil | Bark | HD | 1-Nitro-2-phenylethane (58.2%), methyleugenol (34.7%), and epi-α-cadinol (1.5%) | [64] |
| A. canelilla | Carajás National Forest, Marabá, PA, Brazil | Trunkwood | HD | 1-Nitro-2-phenylethane (47.4%), epi-α-cadinol (19.9%), and methyleugenol (10.5%) | [64] |
| A. canelilla | Carajás National Forest, Marabá, PA, Brazil | Leaf | HD | 1-Nitro-2-phenylethane (68.5%), linalool (8.8%), and spathulenol (4.8%) | [64] |
| A. canelilla | Carajás National Forest, Marabá, PA, Brazil | Bark | HD | 1-Nitro-2-phenylethane (60.5%), methyleugenol (21.3%), and β-sesquiphellandrene (5.4%) | [64] |
| A. canelilla | Carajás National Forest, Marabá, PA, Brazil | Trunkwood | HD | 1-Nitro-2-phenylethane (50.2%), methyleugenol (23.0%), and β-sesquiphellandrene (6.4%) | [64] |
| A. canelilla | Carajás National Forest, Marabá, PA, Brazil | Leaf | HD | Rainy season, Mn Mine: 1-nitro-2-phenylethane (70.6%) and methyleugenol (3.4%) | [84] |
| A. canelilla | Carajás National Forest, Marabá, PA, Brazil | Leaf | HD | Rainy season: Cu Mine: 1-nitro-2-phenylethane (94.5%) and methyleugenol (0.2%) | [84] |
| Zoobotanic park: 1-nitro-2-phenylethane (95.3%) and methyleugenol (0.2%) | |||||
| A. canelilla | Carajás National Forest, Marabá, PA, Brazil | Leaf | HD | Dry season, Mn Mine: 1-nitro-2-phenylethane (39.0%) and methyleugenol (0.5%) | [84] |
| Cu Mine: 1-nitro-2-phenylethane (39.3%) and methyleugenol (0.5%) | |||||
| Zoobotanic park: 1-nitro-2-phenylethane (42.1%) and methyleugenol (0.6%) | |||||
| A. canelilla | Carajás National Forest, Marabá, PA, Brazil | Bark | HD | Rainy season, Mn Mine: 1-nitro-2-phenylethane (94.3%) and methyleugenol (1.0%) | [84] |
| Cu Mine: 1-nitro-2-phenylethane (87.1%) and methyleugenol (8.7%) | |||||
| Zoobotanic park: 1-nitro-2-phenylethane (78.2%) and methyleugenol (14.7%) | |||||
| A. canelilla | Carajás National Forest, Marabá, PA, Brazil | Bark | HD | Dry season: Mn Mine: 1-nitro-2-phenylethane (48.6%) and methyleugenol (45.3%) | [84] |
| Cu Mine: 1-nitro-2-phenylethane (56.2%) and methyleugenol (39.5%) | |||||
| Zoobotanic park: 1-nitro-2-phenylethane (68.1%) and methyleugenol (24.6%) | |||||
| A. canelilla | Carajás National Forest, Marabá, PA, Brazil | Trunkwood | HD | Rainy season, Mn Mine: 1-nitro-2-phenylethane (70.0%) and methyleugenol (17.7%) | [84] |
| A. canelilla | Carajás National Forest, Marabá, PA, Brazil | Trunkwood | HD | Rainy season, Cu Mine: 1-nitro-2-phenylethane (80.0%) and methyleugenol (10.7%) | [84] |
| Zoobotanic park: 1-nitro-2-phenylethane (69.2%) and methyleugenol (15.3%) | |||||
| A. canelilla | Carajás National Forest, Marabá, PA, Brazil | Trunkwood | HD | Dry season, Mn Mine: 1-nitro-2-phenylethane (47.5%) and methyleugenol (48.5%) | [84] |
| Cu Mine: 1-nitro-2-phenylethane (53.3%) and methyleugenol (38.0%) | |||||
| Zoobotanic park: 1-nitro-2-phenylethane (73.3%) and methyleugenol (22.2%) | |||||
| A. canelilla | Cauaxi River, Paragominas, PA, Brazil | Bark | HD | 1-Nitro-2-phenylethane (52.4%), methyleugenol (38.6%) and selin-11-en-4-α-ol (2.5%) | [84,85] |
| A. canelilla | Adolpho Ducke Forest Reserve, Manaus, AM, Brazil | Leaf | HD | 1-Nitro-2-phenylethane (71.2%), benzaldehyde (4.8%), and β-selinene (4.5%) | [65] |
| A. canelilla | Adolpho Ducke Forest Reserve, Manaus, AM, Brazil | Thin twig | HD | 1-Nitro-2-phenylethane (68.2%), eugenol (5.2%), and linalool (5.2%) | [65] |
| A. canelilla | Not reported | Leaf | SD | Commercial sample (market Ver-o-peso): 1-Nitro-2-phenylethane (99.1%) and eugenol (0.9%) | [77] |
| A. canelilla | Not reported | Wood | SD | Commercial sample: 1-Nitro-2-phenylethane (68.8%), methyleugenol (28.1%), and safrole (1.7%) | [77] |
| A. canelilla | Jatapu River, Novo Airão, AM, Brazil | Leaf and thin twig | HD | 1-Nitro-2-phenylethane (91.8%), β-caryophyllene (1.6%), and selin-11-en-4-α-ol (1.3%) | [27] |
| A. canelilla | Jatapu River, Novo Airão, AM, Brazil | Trunkwood | HD | 1-Nitro-2-phenylethane (92.1%), methyleugenol (4.30%), and eugenol (1.2%) | [27] |
| A. canelilla | Cauaxi River, Ulianópolis, PA, Brazil | Leaf and thin twig | HD | 1-Nitro-2-phenylethane (74.0%), linalool (7.6%), and β-caryophyllene (3.5%) | [27] |
| A. canelilla | Cauaxi River, Ulianópolis, PA, Brazil | Barkwood | HD | 1-Nitro-2-phenylethane (90.3%), selin-11-en-4-α-ol (3.5%), and methyleugenol (2.0%) | [27] |
| A. canelilla | Cauaxi River, Ulianópolis, PA, Brazil | Trunkwood | HD | 1-Nitro-2-phenylethane (70.2%), methyleugenol (25.8%), and selin-11-en-4-α-ol (1.2%) | [27] |
| A. canelilla | Adolpho Ducke Forest Reserve, Manaus, AM, Brazil | Leaf | HD | Dry season: 1-nitro-2-phenylethane (88.9%), β-caryophyllene (4.21%), and β-phellandrene (0.80%) | [22] |
| A. canelilla | Adolpho Ducke Forest Reserve, Manaus, AM, Brazil | Leaf | HD | Rainy season: 1-nitro-2-phenylethane (88.5%),β-caryophyllene (5.04%), and β-phellandrene (1.14%) | [22] |
| A. canelilla | Itacoatiara, AM, Brazil | Leaf | HD | 1-Nitro-2-phenylethane (52.2%), eugenol (8.71%) and methyleugenol (5.09%) | [102] |
| A. canelilla | Itacoatiara, AM, Brazil | Thin twig | HD | 1-Nitro-2-phenylethane (92.7%), eugenol (0.3%) and methyleugenol (0.33%) | [102] |
| A. canelilla | Itacoatiara, AM, Brazil | Leaf | HD | Rainy season: 1-nitro-2-phenylethane (31.22-84.33%), β-caryophyllene (1.89-16.34%), and β-selinene (0.77-8.82%) | [101] |
| A. canelilla | Itacoatiara, AM, Brazil | Leaf | HD | Dry season: 1-nitro-2-phenylethane (13.17–74.55%), β-caryophyllene (4.19–12.64%), and β-selinene (1.09–14.98%) | [101] |
| A. canelilla | Itacoatiara, AM, Brazil | Stem | HD | Rainy season: 1-nitro-2-phenylethane (90.94–93.58%), benzeneacetaldehyde (0.48–1.98%) and eugenol (0.07–1.45%) | [101] |
| A. canelilla | Itacoatiara, AM, Brazil | Stem | HD | Dry season: 1-nitro-2-phenylethane (87.85–94.16%), linalool (0.05–3.08%), eugenol (0.08–1.19%) | [101] |
| A. canelilla | Itacoatiara, AM, Brazil | Trunkwood | HD | 1-Nitro-2-phenylethane (83.68%), methyleugenol (14.83%) and neo-intermedeol (0.31%) | [86] |
| A. canelilla | Itacoatiara, AM, Brazil | Bark | HD | 1-Nitro-2-phenylethane (83.2%), methyleugenol (16.2%), and benzaldehyde (0.4%) | [87] |
| A. canelilla | Adolpho Ducke Forest Reserve, Manaus, AM, Brazil | Leaf | HD | 1-Nitro-2-phenylethane (87.34%), ethyl butanoate (3.10%), and α-humulene (0.68%) | [35] |
| A. cinnamomiflora | Los Andes, Merida, Venezuela | wood | HD | γ-Palmitolactone (54.0%), 1-epi-cubenol (9.56%), and δ-cadinene (6.05%) | [88] |
| A. citrifolia | Melgaço, PA, Brazil | Bark | HD | Safrole (16.7%), α-pinene (10.6%), and β-pinene (11.2%) | [73] |
| A. duckei | Manaus, AM, Brazil | Leaf | SD | Seasonal study, linalool (27.3–85.3%) and linalool oxides (5.3–19.7%) | [99] |
| A. duckei | Manaus, AM, Brazil | Thin twig | SD | Seasonal study, linalool (35.7–94.3%) and linalool oxides (3.0-18.7%) | [99] |
| A. duckei | Itacoatiara, AM, Brazil | Leaf and thin twig | HD | Linalool (81.8%), spathulenol (3.8%), and cis-linalool oxide (1.3%) | [51] |
| A. duckei | Adolpho Ducke Forest Reserve, Manaus, AM, Brazil | Leaf | HD | Rainy season: linalool (60.38%, 56.26%) | [103] |
| A. duckei | Adolpho Ducke Forest Reserve, Manaus, AM, Brazil | Leaf | HD | Dry season: linalool (62.40%, 76.69%) | [103] |
| A. duckei | Adolpho Ducke Forest Reserve, Manaus, AM, Brazil | Thin twig | HD | Linalool (89.34%), α-terpineol (3.06%), and cis-linalool oxide (1.94%) | [81] |
| A. duckei | Adolpho Ducke Forest Reserve, Manaus, AM, Brazil | Branch | HD | Linalool (93.60%), α-terpinolene (3.37%), and cis-linalool oxide (3.03%) | [29] |
| A. gardneri | Not reported | Leaf | HD | Benzyl benzoate (44.1%), cadinene (4.8%), and 1,8-cineole (3.6%) | [12] |
| A. gardneri | Not reported | Trunkwood | HD | Benzyl benzoate (78.1%) and phenylethyl benzoate (14.3%) | [12] |
| A. guianensis | Not reported | Leaf | HD | Benzyl benzoate (44.8%) and benzyl salicylate (16.7%) | [12] |
| A. guianensis | Not reported | Wood | HD | Benzyl benzoate (59.0%), benzyl salicylate (6.4%), and methylisoeugenol (5.0%) | [12] |
| A. fragrans | Curuá-una, PA, Brazil | Leaf and thin twig | HD | Linalool (32.4%), spathulenol (19.1%), and limonene (14.5%) | [51] |
| A. hostmanniana | Not reported | Bark | HD | (γ-asarone) 2,4,5-trimethoxyallylbenzene (94.5%) | [12] |
| A. hostmanniana | Not reported | Trunkwood | HD | (γ-asarone) 2,4,5-trimethoxyallylbenzene (98.6%) | [12] |
| A. hostmanniana | Forest of San Eusebio, Andrés Bello, Mérida, Venezuela | Leaf | HD | Benzyl benzoate (29.3%), δ-cadinene (12%), and β-caryophyllene (10.5%) | [68] |
| A. panurensis | Adolpho Ducke Forest Reserve, Manaus, AM, Brazil | Leaf | HD | β-Caryophyllene (33.5%), germacrene-D (25.4%), and α-copaene (7,5%) | [70] |
| A. parviflora | Zoobotanical Park of Emilio Goeldi Museum, Belém, PA, Brazil | Leaf | SD | Benzyl salicylate (34.3), benzyl benzoate (7.2%), and α-pinene (3.9%) | [106] |
| A. parviflora | Zoobotanical Park of Emilio Goeldi Museum, Belém, PA, Brazil | Root | SD | Benzyl benzoate (97.8%) and benzaldehyde (2.2%) | [106] |
| A. parviflora | Santarém, PA, Brazil | Leaf | HS-SPME | Linalool (29.6%), β-caryophyllene (10.9%), and α -phellandrene (10.5%) | [72] |
| A. parviflora | Tomé-Açu, PA, Brazil | Leaf | HD | Linalool (21.30%), β-phellandrene (21.06%), and α-phellandrene (7,25%) | [73] |
| A. parviflora | Tomé-Açu, PA, Brazil | Leaf | HD | β-Phellandrene (23,60%), linalool (12.64%), and spathulenol (4.73%) | [73] |
| A. parviflora | Belém, PA, Brazil | Leaf | HD | β-Phellandrene (15.1%), linalool (14.1%), and γ-eudesmol (12.9%) | [32] |
| A. parviflora | Belém, PA, Brazil | Branch | HD | γ-Eudesmol (16.8%), β-caryophyllene (15.7%), and linalool (12.4%) | [32] |
| A. parviflora | Santarém, PA, Brazil | Leaf | HD | Linalool (45.0%), β-phellandrene (17.3%), and α-phellandrene (4.1%) | [33,71] |
| A. parviflora | Santarém, PA, Brazil | Leaf | HD | Linalool (22.8%), β-caryophyllene (8.15%), and β-phellandrene (7.55%) | [34] |
| A. parviflora | Santarém, PA, Brazil | Thin wig | HD | Linalool (11.90%), aristolene (11.07%), and β-eudesmol (3.97%) | [34] |
| A. parviflora | Belém, PA, Brazil | Leaf | HD | Dry season: linalool (14,07%), spathulenol (7.0%), and oxygenated sesquiterpene (220) (6.7%) | [104] |
| A. parviflora | Belém, PA, Brazil | Leaf | HD | Rainy season: linalool (20.33%, 28.42%), p-cymene (13.04%, 17.54%), and α-phellandrene (12.74%, 14.87%) | [104] |
| A. parviflora | Adolpho Ducke Forest Reserve, Manaus, AM, Brazil | Leaf | HD | Linalool (40.02%), caryophyllene oxide (4.33%), 1,8-cineol (4.02%), and β-phellandrene (4.01%) | [35] |
| A. puchury-minor | Carajás National Forest, Marabá, PA, Brazil | Leaf | SD | Elemicin (23.46%), germacrene B (13.42%) and myristicin (8.43%) | [66] |
| A. puchury-minor | Carajás National Forest, Marabá, PA, Brazil | Stem | SD | (E)-Asarone (52.75%), methyleugenol (17.62%), and isolemicin (13.91%) | [66] |
| A. puchury-minor | Carajás National Forest, Marabá, PA, Brazil | Bark | SD | Methyleugenol (43.10%), (E)-asarone (29.95%), and isolemicin (11.87%) | [66] |
| A. puchury-minor | Carajás National Forest, Canaã dos Carajás, PA, Brazil | Leaf | HD | Elemicin (21.5%), bicyclogermacrene (15.4%), and spathulenol (11.3%) | [67] |
| A. puchury-minor | Carajás National Forest, Canaã dos Carajás, PA, Brazil | Bark | HD | (E)-Methylisoeugenol (43.1%), (E)-asarone (30.0%), (Z)-methylisoeugenol (9.0%) | [67] |
| A. puchury-minor | Carajás National Forest, Canaã dos Carajás, PA, Brazil | Wood | HD | (E)-Asarone (38.2%), (E)-isoelemicin (23.1%), (Z)-methylisoeugenol (14.8%) | [67] |
| A. riparia | Parintins, AM, Brazil | Leaf | HD | β-Caryophyllene (16.9%), α-humulene (14.9%) and bicyclogermacrene (14.1%) | [69] |
| A. riparia | Parintins, AM, Brazil | Branch | HD | (E)-Nerolidol (19.4%), elemol (16.2%), and α-humulene (10.9%) | [69] |
| A. riparia | Parintins, AM, Brazil | Barkwood | HD | Benzyl benzoate (30.9%), terpinen-4-ol (9.3%), and benzyl salicylate (7.9%) | [69] |
| A. riparia | Parintins, AM, Brazil | Trunkwood | HD | Benzyl benzoate (36.2%), benzyl salicylate (7.7%), and cis-calamenene (7.2%) | [69] |
| A. rosaeodora | not reported | Trunkwood | SD | Commercial samples: linalool (72.0-83.0%), cis-linalool oxide (1.10-5.80%), and trans-linalool oxide (1.10-5.20%) | [76] |
| A. rosaeodora | Curuá Una, Santarém, PA, Brazil | Leaf | SD | Linalool (73.0–78.0%), eremophyllene (4.50–6.0%), and cyclosatirene (1.40–2.70%) | [76] |
| 3R-(+)-linalool (77.8%) | |||||
| A. rosaeodora | Curuá Una, Santarém, PA, Brazil | Trunkwood | SD | Linalool (87.0–92.0%), cis-linalool oxide (1.0–1.68%), and trans-linalool oxide (0.90–1.60%) | [76] |
| 3R-(−)-linalool > 50.0% | |||||
| A. rosaeodora | Curuá Una, Santarém, PA, Brazil | Leaf | HD | Linalool (79.0%), oxygenated sesquiterpene (MW220) (5.4%), and β-selinene (2.0%) | [51] |
| A. rosaeodora | not reported | Not reported | HD | Commercial sample (Institute for Medicinal Plants Research Dr. Josif Pancic, Belgrade, Serbia): linalool (81.27%), α-terpineol (4.78%), trans-linalool oxide (2.10%) | [94] |
| A. rosaeodora | Not reported | Leaf | HD | Linalool (81.45%), β-selinene (1.65%), and α-selinene (1.39%) | [30] |
| 3R-(−)-linalool (29.3%) and 3S-(+)-linalool (70.7%) | |||||
| A. rosaeodora | Not reported | Trunkwood | HD | Linalool (85.0%), trans-linalool oxide (2.43%), and cis-linalool oxide (2.21%) | [30] |
| 3R-(−)-linalool (38.3%) and 3S-(+)-linalool (61.7%) | |||||
| A. rosaeodora | Presidente Figueiredo, AM, Brazil | Leaf | HS-SPME | Linalool (77.5%), cis-linalool oxide (7.7%), and trans-linalool oxide (5.6%) | [77] |
| A. rosaeodora | Presidente Figueiredo, AM, Brazil | Trunkwood | HS-SPME | Commercial sample: linalool (86.5%), cis-linalool oxide (4.9%), trans-linalool oxide (4.5%) | [77] |
| A. rosaeodora | Zoobotanical Park of Emilio Goeldi Museum, Belém, PA, Brazil | Leaf | HD | Seasonal study, linalool (68.0–96.1%) | [25] |
| A. rosaeodora | Zoobotanical Park of Emilio Goeldi Museum, Belém, PA, Brazil | Leaf | HD | Linalool (85.5%), β-selinene (0.9%), and trans-linalool oxide (0.8%) | [25] |
| A. rosaeodora | Zoobotanical Park of Emilio Goeldi Museum, Belém, PA, Brazil | Wood | HD | Linalool (84.8%), α-terpineol (2.9%), and geraniol (1.0%) | [25,91] |
| 3S-(−)-linalool (50.62%) and of 3R-(+)-linalool (49.38%) | [90] | ||||
| A. rosaeodora | Novo Airão, AM, Brazil | Leaf | HD | Tachi morphological type: linalool (82.1%), spathulenol (2.0%), caryophyllene oxide (2.0%) | [25] |
| A. rosaeodora | Novo Airão, AM, Brazil | Trunkwood | HD | Tachi morphological type: linalool (84.8%), cis-linalool oxide (1.8%), and trans-linalool oxide (1.8%) | [25] |
| A. rosaeodora | Novo Airão, AM, Brazil | Leaf | HD | Preciosa morphological type: linalool (83.9%), caryophyllene oxide (2.0%), and trans-linalool oxide (1.7%) | [25] |
| A. rosaeodora | Novo Airão, AM, Brazil | Trunkwood | HD | Preciosa morphological type: linalool (79.7%), α-terpineol (5.6%), and cis-linalool oxide (2.7%) | [25] |
| A. rosaeodora | Novo Airão, AM, Brazil) | Leaf | HD | Itaúba morphological type: linalool (79.7%), caryophyllene oxide (3.2%), and β-selinene (1.6%) | [25] |
| A. rosaeodora | Novo Airão, AM, Brazil | Trunkwood | HD | Itaúba morphological type: linalool (78.9%), benzyl benzoate (2.7%), and β-selinene (1.0%) | [25] |
| A. rosaeodora | Novo Airão, AM, Brazil | Leaf | HD | Imbaúba morphological type: linalool (79.8%), spathulenol (4.0%), and trans-linalool oxide (1.7%) | [25] |
| A. rosaeodora | Novo Airão, AM, Brazil | Trunkwood | HD | Imbaúba morphological type: linalool (76.9%), α-terpineol (3.8%), and cis-linalool oxide (1.6%) | [25] |
| A. rosaeodora | French Guiana | Leaf | HD | 3S-(+)-linalool (78–89%) | [107] |
| A. rosaeodora | French Guiana | Branch | HD | 3S-(+)-linalool (5–28%) | [107] |
| A. rosaeodora | French Guiana | Trunkwood | HD | 3R-(−)-linalool (95–99%) | [107] |
| A. rosaeodora | French Guiana | Trunkwood | HD | 3S-(+)-linalool (73.0%), α-terpineol (18.8%), nerol (1.7%) | [107] |
| A. rosaeodora | Novo Airão, AM, Brazil | Trunkwood | HD | Linalool (87.7%), α-terpineol (3.1%), trans-linalool oxide (1.5%), and cis-linalool oxide (1.5%) | [92] |
| A. rosaeodora | Adolpho Ducke Forest Reserve, Manaus, AM, Brazil | Stem | HD | Linalool (86.0%), caryophyllene oxide (2.8%), and trans-linalool oxide (1.5%) | [70] |
| A. rosaeodora | not reported | Not reported | SD | Commercial sample (Erbamea-Istrana, Treviso, Italy): linalool (60.1%), limonene (19.2%), and geraniol (7.8%) | [95] |
| A. rosaeodora | not reported | Not reported | SD | Commercial sample: linalool (80%), and α-terpineol (4.5%) | [97] |
| A. rosaeodora | Maués, AM, Brazil | Leaf and thin twig | SD | A four years sample: linalool (82.15%), α-terpineol (3.60%), and geraniol (1.33%) | [78,79] |
| A. rosaeodora | Maués, AM, Brazil | Leaf and thin twig | SD | A ten years sample: linalool (90.5%), α-terpineol (1.11%), and α-selinene (0.73%) | [79] |
| A. rosaeodora | Maués, AM, Brazil | Leaf and thin twig | SD | A twenty years sample: linalool (87.1%), benzyl benzoate (1.61%), and α-terpineol (1.21%) | [79] |
| A. rosaeodora | Tomé-Açu, PA, Brazil | Leaf | HD | Linalool (50.81%), spathulenol (4.46%), and β-selinene (4.35%) | [73] |
| A. rosaeodora | Tomé-Açu, PA, Brazil | Leaf | HD | Linalool (55.91%), cis-linalool furanoxide (5.59%), and trans-linalool furanoxide (4.43%) | [73] |
| A. rosaeodora | Belém, PA, Brazil | Leaf | HD | Linalool (51.75%), spathulenol (3.82%), and β-selinene (3.79%) | [73] |
| A. rosaeodora | Belém, PA, Brazil | Leaf | HD | Linalool (57.17%), cis-linalool furanoid (4.21%), spathulenol (3.92%) | [73] |
| A. rosaeodora | Belém, PA, Brazil | Leaf | HD | Linalool (43.02%), β-selinene (5.84%), and spathulenol (4.46%) | [73] |
| A. rosaeodora | Belém, PA, Brazil | Leaf | HD | Linalool (42.61%), β-selinene (5.23%), and α-selinene (4.26%) | [73] |
| A. rosaeodora | Belém, PA, Brazil | Leaf | HD | Linalool (45.28%), β-selinene (5.11%), and α-selinene (4.11%) | [73] |
| A. rosaeodora | Belém, PA, Brazil | Leaf | HD | Linalool (42.67%), spathulenol (7.20%), and β-selinene (4.88%) | [73] |
| A. rosaeodora | Belém, PA, Brazil | Leaf | HD | Linalool (52.22%), β-selinene (4.24%), and α-selinene (3.53%) | [73] |
| A. rosaeodora | Belém, PA, Brazil | Leaf | HD | Linalool (55.10%), cis-linalool furanoid (4.28%), and spathulenol (4.25%) | [73] |
| A. rosaeodora | Belém, PA, Brazil | Leaf | HD | Linalool (43.96%), β-selinene (5.25%), and α-selinene (4.24%) | [73] |
| A. rosaeodora | Belém, PA, Brazil | Leaf | HD | Linalool (44.66%), β-selinene (5.40%), and α-selinene (4.49%) | [73] |
| A. rosaeodora | Belém, PA, Brazil | Leaf | HD | Linalool (56.29%), cis-linalool furanoid (4.79%), and spathulenol (3.87%) | [73] |
| A. rosaeodora | Belém, PA, Brazil | Leaf | HD | Linalool (52.72%), spathulenol (5.12%), and β-selinene (4.0%) | [73] |
| A. rosaeodora | Belém, PA, Brazil | Leaf | HD | Linalool (46.90%), β-selinene (4.92%), and spathulenol (4.53%) | [73] |
| A. rosaeodora | Belém, PA, Brazil | Leaf | HD | Linalool (49.24%), spathulenol (6.47%), and β-selinene (4.09%) | [73] |
| A. rosaeodora | Belém, PA, Brazil | Leaf | HD | Linalool (38.48%), β-selinene (6.41%), and α-selinene (5.58%) | [73] |
| A. rosaeodora | Belém, PA, Brazil | Leaf | HD | Linalool (47.49%), benzyl benzoate (6.25%), and β-selinene (4.02%) | [73] |
| A. rosaeodora | Belém, PA, Brazil | Leaf | HD | Linalool (45.41%), β-selinene (4.31%), and spathulenol (4.70%) | [73] |
| A. rosaeodora | Belém, PA, Brazil | Leaf | HD | Linalool (45.99%), α-copaene (5.06%), and β-selinene (4.04%) | [73] |
| A. rosaeodora | Belém, PA, Brazil | Leaf | HD | Linalool (57.22%), spathulenol (6.53%), and α-copaene (5.48%) | [73] |
| A. rosaeodora | Belém, PA, Brazil | Leaf | HD | Linalool (64.26%), α-copaene (3.27%), and spathulenol (3.26%) | [73] |
| A. rosaeodora | Belém, PA, Brazil | Leaf | HD | Linalool (52.68%), spathulenol (4.56%), and β-selinene (4.19%) | [73] |
| A. rosaeodora | Belém, PA, Brazil | Leaf | HD | Linalool (49.93%), spathulenol (4.44%), and β-selinene (3.89%) | [73] |
| A. rosaeodora | Belém, PA, Brazil | Leaf | HD | Linalool (51.88%), β-selinene (4.12%), and cis-linalool furanoid (3.18%) | [73] |
| A. rosaeodora | Belém, PA, Brazil | Leaf | HD | Linalool (54.36%), β-selinene (4.13%), and α-copaene (3.95%) | [73] |
| A. rosaeodora | Belém, PA, Brazil | Leaf | HD | Linalool (71.05%), spathulenol (3.73%), and α-copaene (2.90%) | [73] |
| A. rosaeodora | Belém, PA, Brazil | Leaf | HD | Linalool (70.3%), spathulenol (6.20%), and α-copaene (2.07%) | [73] |
| A. rosaeodora | Belém, PA, Brazil | Leaf | HD | Linalool (43.66%), β-selinene (6.29%), and spathulenol (6.18%) | [73] |
| A. rosaeodora | Belém, PA, Brazil | Leaf | HD | Linalool (50.35%), β-selinene (4.22%), and α-selinene (3.50%) | [73] |
| A. rosaeodora | Belém, PA, Brazil | Leaf | HD | Linalool (48.39%), α-copaene (3.73%), and β-selinene (3.55%) | [73] |
| A. rosaeodora | Belém, PA, Brazil | Leaf | HD | Linalool (61.96%), spathulenol (3.75%), and β-selinene (3.10%) | [73] |
| A. rosaeodora | Curuá-una, Santarém, PA, Brazil | Leaf | HD | Linalool (55.09%), β-selinene (4.49%), and α-selinene (4.14%) | [73] |
| A. rosaeodora | Curuá-una, Santarém, PA, Brazil | Leaf | HD | Linalool (55.93%), β-selinene (3.79%), and α-selinene (3.18%) | [73] |
| A. rosaeodora | Curuá-una, Santarém, PA, Brazil | Leaf | HD | Linalool (48.10%), β-selinene (4.81%), and spathulenol (4.62%) | [73] |
| A. rosaeodora | Santarém, PA, Brazil | Leaf | HD | Linalool (88.6%), cis-linalool oxide (1.5%), and (E)-nerolidyl acetate (1.5%) | [33,71] |
| A. rosaeodora | Brazil | Trunkwood | SD | Commercial sample (Oshadhi Ltd. (Cambridge, England): linalool (44.4%), linalyl acetate (31.4%), and β-caryophyllene (10.5%) | [96] |
| A. rosaeodora | not reported | Trunkwood | SD | Commercial sample: linalool (86.23%), cis-linalool oxide (1.06%), and β-selinene (0.95%) | [98] |
| A. rosaeodora | Maués, AM, Brazil | Leaf | HD | Linalool (81.32%, 83.59%), spathulenol (2.52%, 1.98%), β-selinene (1.4%, 1.4%), and camphene (0.04%, 0.30%) | [80] |
| A. rosaeodora | Maués, AM, Brazil | Branch | HD | Linalool (83.88%, 79.49%), β-selinene (0.38%, 0.63%), spathulenol (0.37%, 1.04%), and camphene (0.17%, 1.03%) | [80] |
| A. rosaeodora | Maués, AM, Brazil | Leaf and branch | HD | Linalool (82.11%, 81.33%), spathulenol (1.01%, 2.06%), β-selinene (0.64%, 1.15%), camphene (0.04%, 0.71%) | [80] |
| A. rosaeodora | Maués, AM, Brazil | Stem | HD | Linalool (86.12%), β-selinene (0.43%), camphene (0.11%), and spathulenol (0.11%) | [80] |
| A. rosaeodora | Novo Aripuanã, AM, Brazil | Leaf | HD | Linalool (71.76%, 73.89%), spathulenol (3.16%, 2.13%), β-selinene (2.71%, 2.19%), and camphene (0.76%) | [80] |
| A. rosaeodora | Novo Aripuanã, AM, Brazil | Branch | HD | Linalool (81.53%, 65.08%), β-selinene (0.61%, 0.76%), camphene (0.29%, 1.62%), and spathulenol (0.34%, 0.93%) | [80] |
| A. rosaeodora | Novo Aripuanã, AM, Brazil | Leaf and branch | HD | Linalool (75.57%, 73.24%), spathulenol (2.03%, 2.15%), β-selinene (1.62%, 1.79%), and camphene (0.97%) | [80] |
| A. rosaeodora | Novo Aripuanã, AM, Brazil | Stem | HD | Linalool (81.77%), β-selinene (0.64%), spathulenol (0.24%), and camphene (0.16%) | [80] |
| A. rosaeodora | Manaus, AM, Brazil | Leaf | HD | Dry season: linalool (69.0%), β-phellandrene (2.9%), and bulnesol (1.8%) | [28] |
| A. rosaeodora | Manaus, AM, Brazil | Leaf | HD | Rainy season: linalool (71.0%), butanoic acid (3.5%), and trans-linalool oxide (2.8%) | [28] |
| A. rosaeodora | Manaus, AM, Brazil | Thin twig | HD | Dry season: linalool (78.0%), β-phellandrene (1.5%), and α-eudesmol (1.4%) | [28] |
| A. rosaeodora | Manaus, AM, Brazil | Thin twig | HD | Rainy season: linalool (84.0%), butanoic acid (3.0%), and cis-linalool oxide (2.0%) | [28] |
| A. rosaeodora | Floresta Nacional do Tapajós- Rurópolis (PA, Brazil) | Leaf and thin twig | HD | Linalool (83.7%), aromadendrene oxide (2.5%), and spathulenol (1.6%) | [74] |
| A. rosaeodora | Reserva Extrativista Tapajós-Arapiuns (PA, Brazil) | Leaf and thin twig | HD | Linalool (39.6%), α-phellandrene (22.8%) and p-cymene (7.0%) | [74] |
| A. rosaeodora | Adolpho Ducke Forest Reserve, Manaus, AM, Brazil | Leaf and thin twig | HD | Linalool (93.60%), α-terpinolene (3.37%), and cis-linalool oxide (3.03%) | [81,82] |
| A. rosaeodora | Adolpho Ducke Forest Reserve, Manaus, AM, Brazil | Stem | HD | Linalool (63.16%), trans-linalool oxide (9.73%) and cis-linalool oxide (7.69%) | [93] |
| A. terminalis | Zoobotanical Park of Emilio Goeldi Museum, Belém, PA, Brazil | Leaf and thin twig | HD | α-Phellandrene (32.8%), linalool (21.7%), and p-cymene (16.7%) | [75] |
| A. terminalis | Zoobotanical Park of Emilio Goeldi Museum, Belém, PA, Brazil | Inflorescence | HD | Linalool (36.2%), α-phellandrene (30.7%), and (E)-β-ocimene (22.3%) | [75] |
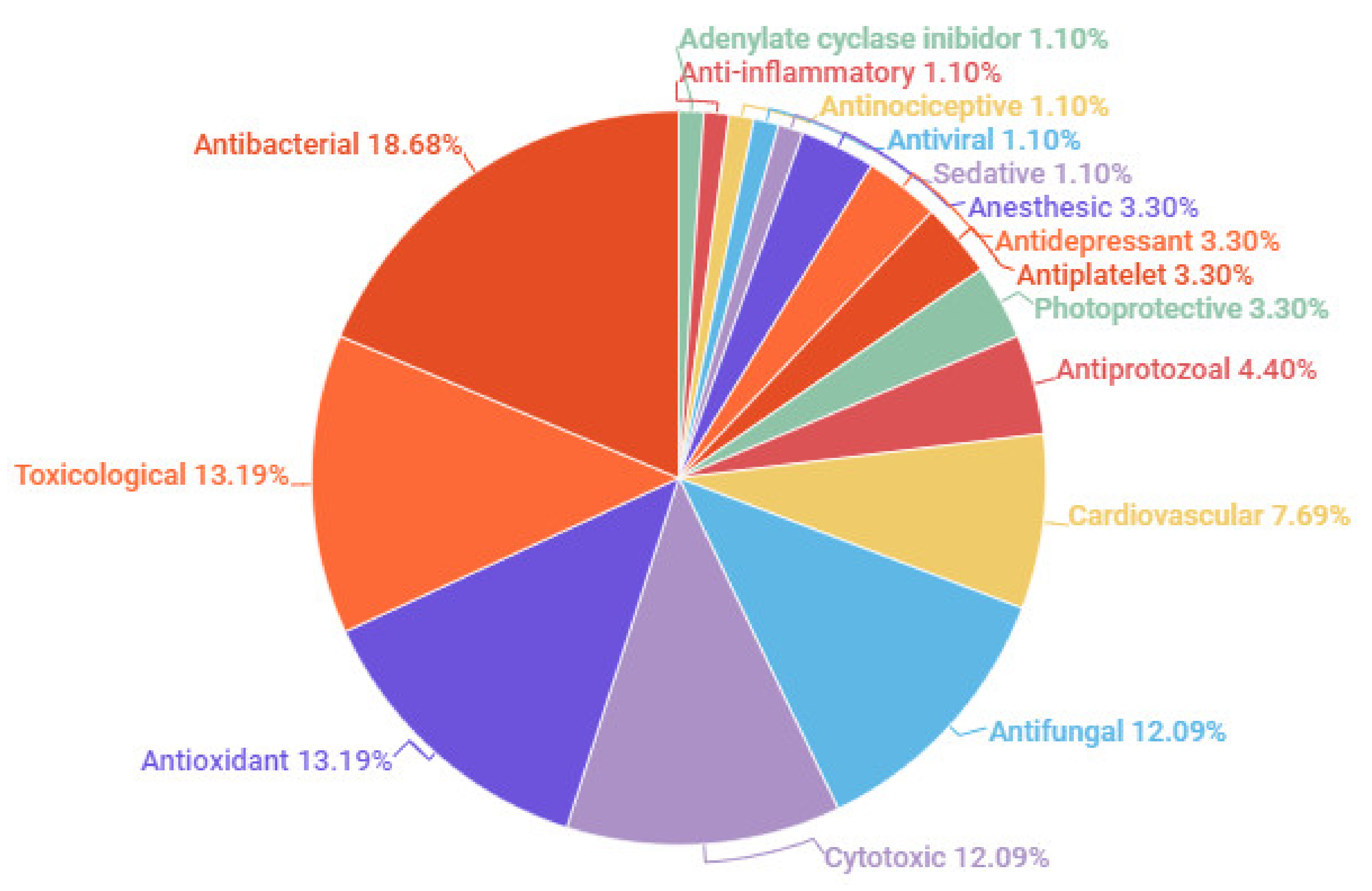
7. Biological Activities
7.1. Antibacterial Activity
| Aniba Species | Collection Site | Plant Part | Major Componentes | Bioactivities | References |
|---|---|---|---|---|---|
| A. canelilla | Adolpho Ducke Forest Reserve, Manaus, AM, Brazil | Leaf | 1-Nitro-2-phenylethane (88.9%), β-caryophyllene (4.21%), and β-phellandrene (0.80%) | Anti-leishmanial (IC50 40 μg/mL, Leishmania amazonensis promastigotes); cytotoxic (mice BALB-c macrophage, MTT assay, IC50 9.3 μg/mL); toxicological (LC50 68.37 μg/mL, Artemia salina lethality) | [22] |
| A. canelilla | Adolpho Ducke Forest Reserve, Manaus, AM, Brazil | Leaf | 1-Nitro-2-phenylethane (88.9%), β-caryophyllene (4.21%), and β-phellandrene (0.80%) | Anti-leishmanial (IC50 40 μg/mL, Leishmania amazonensis promastigotes); cytotoxic (mice BALB-c macrophage, MTT assay, IC50 9.3 μg/mL); toxicological (LC50 68.37 μg/mL, Artemia salina lethality) | [22] |
| A. canelilla | Cauaxi River, Paragominas, PA, Brazil | Bark | 1-Nitro-2-phenylethane (52.4%), methyleugenol (38.6%) and selin-11-en-4-α-ol (2.5%); | Cardiovascular, hypotension and bradycardia, EO at 1–10 mg/kg; vasorelaxant effects, IC50 19 μg/mL; | [23] |
| A. canelilla | Cauaxi River, Paragominas, PA, Brazil | Bark | 1-Nitro-2-phenylethane, isolated | Cardiovascular, hypotension and bradycardia, EO at 1–20 mg/kg; vasorelaxant effects, IC50 29.6 μg/mL | [23] |
| A. canelilla | Cauaxi River, Paragominas, PA, Brazil | Bark | 1-Nitro-2-phenylethane (52.4%), methyleugenol (38.6%) and selin-11-en-4α-ol (2.5%), and 1-nitro-2-phenylethane, isolated | Cardiovascular, vasorelaxant effects (IC50 294.19 μg/mL) and 1-nitro-2-phenylethane isolated (IC50 501.27 μg/mL) | [24] |
| A. canelilla | Jatapu River, Novo Airão, AM, Brazil | Trunk wood | 1-Nitro-2-phenylethane (92.1%), methyleugenol (4.30%), and eugenol (1.2%) | Antioxidant (EC50 223.81 μg/mL, DPPH method); toxicological (LC50 21.61 μg/mL, Artemia salina lethality) | [27] |
| A. canelilla | Cauaxi River, Ulianópolis, PA, Brazil | Trunk wood | 1-Nitro-2-phenylethane (70.2%), methyleugenol (25.8%), and selin-11-en-4α-ol (1.2%) | Antioxidant (EC50 172.52 μg/mL, DPPH method); toxicological (LC50 21.61 μg/mL, Artemia salina lethality assay) | [27] |
| A. canelilla | Adolpho Ducke Forest Reserve, Manaus, AM, Brazil | Leaf | 1-Nitro-2-phenylethane (87.34%), ethyl butanoate (3.10%), and α-humulene (0.68%) | Antifungal (Aspergillus flavus, Colletotrichum guaranicola, MIC 0.15 μg/mL; A. niger, MIC 0.3 μg/mL; Fusarium oxysporum, F. solani, C. gloeosporioides, C. musae, MIC 0.62 μg/mL; Alternaria alternata MIC 5.0 μg/mL, agar-well diffusion method) | [35] |
| A. canelilla | not reported | Bark stem | 1-Nitro-2-phenylethane (73.0%), methyleugenol (19.2%), safrole (3.7%), and eugenol (1.5%) | Antimicrobial (Staphylococcus aureus, Candida albicans, C. parapsilosis and C. krusei, agar diffusion method) | [77] |
| A. canelilla | Cauaxi River, Paragominas, PA, Brazil | Bark | 1-Nitro-2-phenylethane (52.4%), methyleugenol (38.6%) and selin-11-en-4α-ol (2.5%) | Cardiovascular (Rattus norvegicus male Wistar rat model), hypotension and bradycardia EO at 1, 5, 10, and 20 mg/kg; vasorelaxant effects, IC50 109.5 μg/mL | [85] |
| A. canelilla | Itacoatiara, AM, Brazil | Wood | 1-Nitro-2-phenylethane (83.68%), methyleugenol (14.83%) and neointermedeol (0.31%) | Trypanocide (Trypanosoma evansi, mortality after 6 h, EO at 0.5 to 2.0%); Cytotoxic (human lymphocytes, EO at 0.5 to 2.0%, MTT assay) | [86] |
| A. canelilla | Cauaxi River, Ulianópolis, PA, Brazil | Bark wood | 1-Nitro-2-phenylethane | Antinociceptive and anti-inflammatory (abdominal writhing method) | [110] |
| A. canelilla | Cauaxi River, Ulianópolis, PA, Brazil | Trunk wood | 1-Nitro-2-phenylethane, isolated | Anti-inflammatory (25 and 50 mg/kg, paw and ear edema in male Swiss mice and Wistar rats) | [111] |
| A. canelilla | Adolpho Ducke Forest Reserve, Manaus, AM, Brazil. | Leaf | Not reported | Photoprotective (dry season, FPS 7.54; rainy season FPS 14.08, spectrophotometric method) | [112] |
| A. canelilla | Adolpho Ducke Forest Reserve, Manaus, AM, Brazil. | Thin twig | Not reported | Photoprotective (dry season, FPS 5.49; rainy season FPS 6.93, spectrophotometric method) | [112] |
| A. canelilla | Cauaxi River, Paragominas, PA, Brazil | Bark | 1-Nitro-2-phenylethane, isolated | Cardiovascular (male Wistar rat model), induced hypotensive and bradycardic, EO at 1–10 mg/kg; vasorelaxant effects, IC50 60.1 μg/mL | [113] |
| A. canelilla | Cauaxi River, Paragominas, PA, Brazil | Bark | 1-Nitro-2-phenylethane, isolated | Cardiovascular (Male Wistar rat model); vasorelaxant effects IC50 203.1 μM, contractions induced by phenylephrine; Hyp9 IC50 119.0 μM and phorbol 12,13-dibutyrate IC50 43 μM | [114] |
| A. canelilla | not reported | Not reported | 1-Nitro-2-phenylethane, isolated | Cardiovascular, vasorelaxant effects at concentration of 0.1–100 μg/mL | [115] |
| A. canelilla | Cauaxi River, Ulianópolis, PA, Brazil | Trunk wood | 1-Nitro-2-phenylethane (70.2%), methyleugenol (25.8%), and selin-11-en-4α-ol (1.2%); 1-nitro-2-phenylethane, isolated | Acetylcholinesterase (detection limit of 0.01 ng to EO and 1-nitro-2-phenylethane, isolated, TLC bioautography method) | [116] |
| A. duckei | Adolpho Ducke Forest Reserve, Manaus, AM, Brazil | Twig | Linalool (93.6%), α-terpinolene (3.37%), and cis-linalool oxide (3.03%) | Antifungal (Colletotrichum gloesporioides and Fusarium oxysporum, mycelial growth inhibition in 100% at 0.4%) | [29] |
| A. duckei | Adolpho Ducke Forest Reserve, Manaus, AM, Brazil | Twig | Linalool (89.34%), α-terpineol (3.06%), and cis-linalool oxide (1.94%) | Toxicological (Aedes aegypti, LC50 250.61 μg/mL, EO; LC50 279.89 μg/mL, 3R-(–)-linalool; LC50 346 μg/mL, (±)-linalool | [81] |
| 3R-(–)-linalool and (±)-linalool standard | |||||
| A. duckei | Adolpho Ducke Forest Reserve, Manaus, AM, Brazil | Leaf | Not reported | Toxicological (Artemia franciscana, 100% of larval mortality after 10min, EO at 2 μg/mL) | [117] |
| A. duckei | Adolpho Ducke Forest Reserve, Manaus, AM, Brazil | Twig | Not reported | Toxicological (Aedes aegypti, LC90 54 × 103 μg/mL and LC50 2.2 × 102 μg/mL) | [117] |
| A. hostmanniana | Forest of San Eusebio, Andrés Bello, Mérida, Venezuela | Leaf | Benzyl benzoate (29.3%), δ-cadinene (12%), and β-caryophyllene (10.5%) | Antibacterial (Pseudomonas aeruginosa and Staphylococcus aureus, MIC 900 μg/mL, Klebsiella pneumonia, MIC1250 μg/mL, broth microdilution method) | [68] |
| A. panurensis | Adolpho Ducke Forest Reserve, Manaus, AM, Brazil | Leaf | β-Caryophyllene (33.5%), germacrene-D (25.4%), α-copaene (7.5%) and β-bourbonene (7.1%) | Antioxidant (EC50 > 1000 μg/mL, DPPH method); Antiplatelet activity (3.57%) | [70] |
| A. parviflora | Belém (PA, Brazil) | Leaf | β-Phellandrene (15.1%), linalool (14.1%), and γ-eudesmol (12.9%) | Antibacterial (Escherichia coli, MIC 19.5 μg/mL; Bacillus cereus, MIC: 312.5 μg/mL; Staphylococcus aureus, S. epidermidis, MIC: 625 μg/mL, Pseudomonas aeruginosa, MIC 1250 μg/mL, microdilution method); antioxidant (90.1–287.9 mg TE/mL, DPPH method); cytotoxic (MCF-7 mammary adenocarcinoma, IC50 67.9 μg/mL, MTT assay) | [32] |
| A. parviflora | Belém (PA, Brazil) | Branch | γ-Eudesmol (16.8%), β-caryophyllene (15.7%), and linalool (12.4%) | Antibacterial (Escherichia coli, MIC 19.5 μg/mL; Bacillus cereus, MIC: 1250 μg/mL; Staphylococcus aureus MIC: 625 μg/mL, S. epidermidis 1250 μg/mL, Pseudomonas aeruginosa, MIC 1250 μg/mL, microdilution method); antioxidant (94.1–358.4 mgTE/mL, DPPH method); cytotoxic (MCF-7 mammary adenocarcinoma, IC50 102.2 μg/mL MTT assay) | [32] |
| A. parviflora | Curuá-una, Santarém, PA, Brazil | Leaf | Linalool (45.0%), β-phellandrene (17.3%), and α-phellandrene (4.1%) | Antibacterial (Klesbsiella pneumoniae Enterococcus faecalis, Staphylococcus aureus and S. epidermidis MIC > 10 μL/mL, Streptococcus pyogenes MIC 1.3 μL/mL, agar disk diffusion method) | [33] |
| A. parviflora | Santarém, PA, Brazil | Leaf | Linalool (22.8%), caryophyllene (8.15 %), β-phellandrene (7.55%), and o-cymene (6.19%) | Antibacterial (Staphylococcus aureus and Enterococcus faecalis, MIC 2,0 mg/mL, agar disk diffusion method) | [34] |
| Twig | Linalool (11.90%), aristolene (11.7%),β-eudesmol (3.97%), and spathulenol (3.51%) | ||||
| A. parviflora | Adolpho Ducke Forest Reserve, Manaus, AM, Brazil | Leaf | Linalool (40.02%), caryophyllene oxide (4.33%), 1,8-cineole (4.02%), and β-phellandrene (4.01%) | Antifungal (Aspergillus flavus, Fusarium solani, F. oxysporum, MIC 0.62 µL/mL; A. niger, Colletotrichum guaranicola, MIC 2.5 µL/mL; C. gloeosporioides, MIC 0.62 µL/mL; Alternaria alternata, C. musae, MIC 5.0 µL/mL, agar-well diffusion method) | [35] |
| A. parviflora | Curuá-una, Santarém, PA, Brazil | Leaf | Linalool (45.0%), β-phellandrene (17.3%), α-phellandrene (4.1%), and (E)-caryophyllene (3.9%) | Antidepressant activity (Male Wistar rats, EO at 85 mg/kg) | [71] |
| A. parviflora | Santarém, PA, Brazil | Leaf | Linalool (29.60%), β-caryophyllene (10.9%), and α-phellandrene (10.5%) | Anesthetic potential to the fish species Colossoma macropomum. EO at 0.1 μL/mL light sedation (46.4 s), deep sedation (120.8 s), deep anesthesia (333.2 s) | [118] |
| A. parviflora | Curauá, Santarém, PA, Brazil | Bark | Linalool (16.3%), α-humulene (14.5%), δ-cadinene (10.2%), α-copaene (9.51%) and germacrene B (7.58%) | Cytotoxic (Human hepatocellular carcinoma cells HepG2; IC50 values of 9.05 μg/mL; xenograft model). | [119] |
| A. rosaeodora | Manaus, AM, Brazil | Leaf | Dry season: linalool (69.0%), β-phellandrene (2.9%), and bulnesol (1.8%) | Antifungal activity (Colletotrichum gloeosporioides, MIC 1.25 μL/mL, Colletotrichum sp., MIC 2.5 μL/mL; C. guaranicola, MIC 0.62 μL/mL; Alternaria alternata, MIC 1.25 μL/mL, agar-well diffusion method) | [28] |
| Rainy season: linalool (71.0%), butanoic acid (3.5%), and trans-linalool oxide (2.8%) | Antifungal activity (Colletotrichum gloeosporioides rainy season, MIC 2.5 μL/mL; Colletotrichum sp., MIC 5.0 μL/mL; C. guaranicola, MIC 1.25 μL/mL; Alternaria alternata, MIC 2.5 μL/mL, agar well diffusion method) | ||||
| A. rosaeodora | Manaus, AM, Brazil | Thin twig | Dry season: linalool (78.0%), β-phellandrene (1.5%), and α-eudesmol (1.4%) | Antifungal activity (Colletotrichum gloeosporioides, MIC 1.25 μL/mL; Colletotrichum sp., dry season, MIC 2.5 μL/mL; C. guaranicola, MIC 1.25 μL/mL; Alternaria alternata, MIC 1.25 μL/mL, agar well diffusion method) | [28] |
| Rainy season: linalool (84.0%), butanoic acid (3.0%), and cis-linalool oxide (2.0%) | Antifungal activity (Colletotrichum gloeosporioides, MIC 2.5 μL/mL; Colletotrichum sp., MIC 5.0 μL/mL; C. guaranicola, MIC 1.25 μL/mL; Alternaria alternata, MIC 2.5 μL/mL, agar well diffusion method) | ||||
| A. rosaeodora | Curuá-una, Santarém, PA, Brazil | Leaf | Linalool (88.6%), cis-linalool oxide (1.5%), and (E)-nerolidyl acetate (1.5%) | Antibacterial (Escherichia coli and Klesbsiella pneumoniae MIC > 10 µL/mL; Enterococcus faecalis and Staphylococcus epidermidis MIC 5 µL/mL, S. aureus and Streptococcus pyogenes MIC 1.3 µL/mL, agar disk diffusion method) | [33] |
| A. rosaeodora | Adolpho Ducke Forest Reserve, Manaus, AM, Brazil | Stem | Linalool (86.0%), caryophyllene oxide (2.8%), and trans-linalool oxide (1.5%) | Antioxidant (EC50 733 µg/mL, DPPH method); Antiplatelet activity (5.19%) | [70] |
| A. rosaeodora | Curuá-una, Santarém, PA, Brazil | Leaf | Linalool (88.6%), cis-linalool oxide (1.5%), and (E)-nerolidyl acetate (1.5%) | Isolated linalool at 0.1 µL/mL, light sedation (61.8 s), deep sedation (92.1 s) and deep anesthesia (464.3 s) | [71] |
| A. rosaeodora | Adolpho Ducke Forest Reserve, Manaus, AM, Brazil | Leaves and branches | Linalool (93.60%), α-terpinolene (3.37%), cis-linalool oxide (3.03%) and standard linalool | Antibacterial activity (MIC to EO 250–450 µg/mL and to standard linalool 550–650 µg/mL, disk-diffusion method); antioxidant activity (IC50: 15.46 μg/mL, ABTS method); trypanocide (Trypanosoma cruzi, IC50 to EO 150.5–911.6 µg/mL and to standard linalool 198.6–249.6 µg/mL); cytotoxicity (peritoneal macrophages from Balb/C mice, CC50 > 1000 µg/mL, MTT assay). | [81] |
| A. rosaeodora | Adolpho Ducke Forest Reserve, Manaus, AM, Brazil | Stem | Linalool (93.60%), α-terpinolene (3.37%), and cis-linalool oxide (3.03%) | Antibacterial activity (E. coli and S. aureus, MIC 200 and 150 µg/mL, respectively, disk-diffusion method); toxicological (Artemia salina, LC50: 282 mg/L). | [82] |
| A. rosaeodora | Zoobotanical Park of Emilio Goeldi Museum, Belém, PA, Brazil | Trunk wood | Linalool (84.8%), α-terpineol (2.9%), and geraniol (1.0%) | Relaxant and anticonvulsant (inhibition of cAMP, EO, IC50 130 µg/mL; 3R-(−)-linalool, IC50 310 µM; (±)-linalool, IC50: 300 µM) | [90] |
| Mixture racemic, linalool: 3R-(–)-linalool (50.62%) and 3S-(+)-linalool (49.38%) | |||||
| A. rosaeodora | Zoobotanical Park of Emilio Goeldi Museum, Belém, PA, Brazil | Trunk wood | Linalool (84.8%), α-terpineol (2.9%), and geraniol (1.0%) | Antibacterial (Escherichia coli, EO at 0.3% chicken gastrointestinal tract) | [91] |
| A. rosaeodora | Jatapu River, Novo Airão, AM, Brazil | Trunk wood | Linalool (87.7%), α-terpineol (3.1%), trans-linalool oxide (1.5%), and cis-linalool oxide (1.5%) | Sedative potential (Male Swiss albino mice) EO at 100-300 mg/kg; EO at 2 and 100 µg/mL, percentages of blocking effect sciatic nerves from 75 to 95% | [92] |
| A. rosaeodora | Adolpho Ducke Forest Reserve, Manaus, AM, Brazil | Stem | Linalool (63.16%), trans-linalool furanoid (9.73%) and cis-linalool furanoid (7.69%) | Antioxidant activity (IC50: 48.67 and 40.06 µg/mL, ABTS and DPPH methods, respectively); toxicological (Artemia salina, LC50 282-582 µg/mL; Aedes aegypti, LC50 41.07 µg/mL) | [93] |
| A. rosaeodora | Maués, AM, Brazil | Leaf | Linalool (90.5%), synthetic linalool and isolated linalool | Anesthetic potential to the fish species Colossoma macropomum. EO at 0.050 µL/mL light sedation (68.3 s), deep sedation (204.1 s) and deep anesthesia (636.4 s) | [118] |
| A. rosaeodora | Maués, AM, Brazil | Leaf | Linalool (90.5%), synthetic linalool and isolated linalool | Synthetic linalool at 0.1 µL/mL, light sedation (32.7 s), deep sedation (68.9 s) and deep anesthesia (198.7 s) | [118] |
| Isolated linalool at 0.1 µL/mL, light sedation (61.8 s), deep sedation (92.1 s) and deep anesthesia (464.3 s) | |||||
| A. rosaeodora | Zoobotanical Park of Emilio Goeldi Museum, Belém, PA, Brazil | Trunk wood | Not reported | Antiviral (Avian metapneumovirus, EC50: 20.86 µg/mL); cytotoxicity, cells derived from bovine kidney (CRIB), chicken embryo (CRER), mouse fibroblast cell (L929) and feline kidney cell lines (CRFK) CC50: 104.8 µg/mL | [120] |
| A. rosaeodora | Jatapu River, Novo Airão, AM, Brazil | Trunk wood | Linalool (87.7%), α-terpineol (3.1%), trans-linalool oxide (1.5%), and cis-linalool oxide (1.5%) | Cardiovascular (Male Wistar rat model), hypotension and bradycardia, EO at 10–20 mg/kg; vasorelaxant effects, IC50: 72.35 µg/mL | [121] |
7.2. Antifungal Activity
7.3. Anti-Inflammatory and Antinociceptive Activities
7.4. Antioxidant and Photoprotective Activities
7.5. Cardiovascular Activity
7.6. Cytotoxic Activity
7.7. Nervous System Activity
7.8. Toxicological Studies
7.9. Other Activities
7.10. Biological Activities from Commercial Aniba Rosaeodora Essential Oils
8. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- The Angiosperm Phylogeny Group. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Brasil, F.D.; Lauraceae in Flora do Brasil 2020 em Construção. Lauraceae in Flora do Brasil 2020 em construção. In Jard. Botânico do Rio Janeiro; 2020. Available online: http://www.floradobrasil.jbrj.gov.br/reflora/floradobrasil/FB23347 (accessed on 23 April 2020).
- Kubitzki, K.; Renner, S. Lauraceae I (Aniba and Aiouea). Flora Neotrop. 1982, 31, 1–124. Available online: http://www.jstor.org/stable/4393757 (accessed on 2 September 2021).
- De Moraes, P.L.R. Flora das cangas da Serra dos Carajás, Pará, Brasil: Lauraceae. Rodriguésia 2018, 69, 81–117. [Google Scholar] [CrossRef] [Green Version]
- Bastos, A.M. Os paus rosas da indústria da essência. Rodriguesia 1943, 7, 45–54. [Google Scholar]
- Richter, H.G. Wood and Bark Anatomy of Lauraceae. I. Aniba Aublet. IAWA J. 1981, 2, 79–87. [Google Scholar] [CrossRef]
- Ribeiro, J.E.L.S.; Hopkins, M.J.G.; Vicentini, A.; Sothers, C.A.; Costa, M.A.S.; Brito, J.M.; Souza, M.A.D.; Martins, L.H.P.; Lohmann, L.G.; Assunção, P.A.C.L.; et al. (Eds.) Flora da Reserva Ducke: Guia de Identificação das Plantas Vasculares de uma Floresta de Terra Firme na Amazônia Central; INPA-DFID: Manaus, AM, USA, 1999; p. 800. [Google Scholar]
- Quinet, A. Sinopse taxonômica da família Lauraceae no Estado do Rio de Janeiro, Brasil. Acta Bot. Bras. 2005, 19, 563–572. [Google Scholar] [CrossRef] [Green Version]
- Vattimo, Í.D. Anatomia da folha das espécies conhecidas de pau-rosa. X—Citomorfologia das epidermes superior e inferior de Aniba permollis (Nees) Mez (Lauraceae). Acta Amaz. 1975, 5, 45–50. [Google Scholar] [CrossRef] [Green Version]
- Loureiro, A.A. Estudo anatômico macro e microscópico de 10 espécies do Gênero Aniba (Lauraceae) da Amazônia. Acta Amaz. 1976, 6, 5–85. [Google Scholar] [CrossRef] [Green Version]
- Urbinati, C.V.; Da Silva, R.C.R.; Da Silva, S.S.; Lisboa, P.L.B.; Mori, F.A. Structure of the secondary xylem of Aniba Aubl. species from the Brazilian Amazon. Acta Bot. Bras. 2014, 28, 484–488. [Google Scholar] [CrossRef] [Green Version]
- De Morais, A.A.; Rezende, C.M.A.D.M.; Mourão, J.C.; Gottlieb, O.R.; Marx, M.C.; Von Bülow, M.V.; Da Rocha, A.I.; Magalhaes, M.T. Óleos essenciais de espécies do gênero Aniba. Acta Amaz. 1972, 2, 41–44. [Google Scholar] [CrossRef]
- Gottlieb, O.R.; Kubitzki, K. Chemosystematics of Aniba. Biochem. Syst. Ecol. 1981, 9, 5–12. [Google Scholar] [CrossRef]
- Chanderbali, A.S.; Van der Werff, H.; Renner, S.S. Phylogeny and historical biogeography of Lauraceae: Evidence from the chloroplast and nuclear genomes. Ann. Missouri. Bot. Gard. 2001, 88, 104–134. [Google Scholar] [CrossRef] [Green Version]
- Angrizani, R.C.; Contim, L.A.S.; Lemes, M.R. Development and Characterization of Microsatellite Markers for the Endangered Amazonian Tree Aniba rosaeodora (Lauraceae). Appl. Plant. Sci. 2013, 1, 1200516. [Google Scholar] [CrossRef] [PubMed]
- Xavier, J.; Maia, L.; Figueiredo, P.; Folador, A.; Ramos, A.; Andrade, E.; Maia, J.; Setzer, W.; da Silva, J. Essential Oil Composition and DNA Barcode and Identification of Aniba species (Lauraceae) Growing in the Amazon Region. Molecules 2021, 26, 1914. [Google Scholar] [CrossRef]
- Franciscon, C.H.; Miranda, I.D.S. Distribution and conservation of Aniba Aubl. (Lauraceae Jussieu) species in Brazil. Biota Neotropica 2018, 18. [Google Scholar] [CrossRef]
- Gottlieb, O. Chemosystematics of the lauraceae. Phytochemistry 1972, 11, 1537–1570. [Google Scholar] [CrossRef]
- Fernandes, J.B.; Gottlieb, O.R.; Xavier, L.M. Chemosystematic implications of flavonoids in Aniba riparia. Biochem. Syst. Ecol. 1978, 6, 55–58. [Google Scholar] [CrossRef]
- Rossi, M.H.; Yoshida, M.; Maia, J.G.S. Neolignans, styrylpyrones and flavonoids from an Aniba species. Phytochemistry 1997, 45, 1263–1269. [Google Scholar] [CrossRef]
- Ferreira, Z.S.; Gottlieb, O.R.; Roque, N.F. Chemosystematic implications of benzyltetrahydroisoquinolines in Aniba. Biochem. Syst. Ecol. 1980, 8, 51–54. [Google Scholar] [CrossRef]
- Silva, J.R.D.A.; Carmo, D.F.M.D.; Reis, M.; Machado, G.M.C.; Leon, L.L.; da Silva, B.O.; Ferreira, J.L.P.; Amaral, A.C.F. Chemical and biological evaluation of essential oils with economic value from Lauraceae species. J. Braz. Chem. Soc. 2009, 20, 1071–1076. [Google Scholar] [CrossRef] [Green Version]
- Interaminense, L.D.F.L.; De Siqueira, R.J.B.; Xavier, F.E.; Duarte, G.P.; Magalhães, P.J.C.; Da Silva, J.K.; Maia, J.G.S.; Sousa, P.J.D.C.; Leal-Cardoso, J.H.; Lahlou, S. Cardiovascular effects of 1-nitro-2-phenylethane, the main constituent of the essential oil of Aniba canelilla, in spontaneously hypertensive rats. Fundam. Clin. Pharmacol. 2010, 25, 661–669. [Google Scholar] [CrossRef]
- Interaminense, L.D.F.L.; dos Ramos-Alves, F.E.; de Siqueira, R.J.B.; Xavier, F.; Duarte, G.P.; Magalhaes, P.; Maia, J.G.S.; Sousa, P.J.D.C.; Lahlou, S. Vasorelaxant effects of 1-nitro-2-phenylethane, the main constituent of the essential oil of Aniba canelilla, in superior mesenteric arteries from spontaneously hypertensive rats. Eur. J. Pharm. Sci. 2013, 48, 709–716. [Google Scholar] [CrossRef]
- Maia, J.G.S.; Andrade, E.H.A.; Couto, H.A.R.; Da Silva, A.C.M.; Marx, F.; Henke, C. Plant sources of amazon rosewood oil. Quim Nova 2007, 30, 1906–1910. [Google Scholar] [CrossRef]
- Maia, J.G.S.; Andrade, E.H.D.A. Database of the Amazon aromatic plants and their essential oils. Quim Nova 2009, 32, 595–622. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, J.K.R.; Sousa, P.J.C.; Andrade, E.H.A.; Maia, J.G.S. Antioxidant Capacity and Cytotoxicity of Essential Oil and Methanol Extract of Aniba canelilla (H.B.K.) Mez. J. Agric. Food Chem. 2007, 55, 9422–9426. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, R.B.; Souza, D.P.; Albuquerque, P.M.; Fernandes, A.V.; Santos, A.S.; Duvoisin, S.; Gonçalves, J.F. Variability and antifungal activity of volatile compounds from Aniba rosaeodora Ducke, harvested from Central Amazonia in two different seasons. Ind. Crop. Prod. 2018, 123, 1–9. [Google Scholar] [CrossRef]
- Gomes, P.R.B.; Cunha, M.J.O.; DE Sousa, D.A.; Reis, J.B.; Melo, A.V.; De Freitas, A.C.; Hunaldo, V.K.L.; Fontenele, M.A.; De Paula, M.D.L.; Louzeiro, H.C.; et al. Chemical Study and Antifungal Activity of the Essential Oil of the Branches of Aniba duckei Kostermans. J. Essent. Oil Bear. Plants 2019, 22, 1554–1561. [Google Scholar] [CrossRef]
- Zellner, B.D.; Presti, M.L.; Barata, L.E.S.; Dugo, P.; Dugo, G.; Mondello, L. Evaluation of Leaf-Derived Extracts as an Environmentally Sustainable Source of Essential Oils by Using Gas Chromatography−Mass Spectrometry and Enantioselective Gas Chromatography—Olfactometry. Anal. Chem. 2005, 78, 883–890. [Google Scholar] [CrossRef] [PubMed]
- Tranchida, P.Q.; Souza RC, Z.; Barata LE, S.; Mondello, M.; Dugo, P.; Dugo, G.; Mondello, L. Analysis of Macacaporanga (Aniba parviflora) leaf essential oil by using comprehensive two-dimensional gas chromatography combined with rapid scan-ning quadrupole mass spectrometry. Chromatogr Today 2008, 95, 5–9. [Google Scholar]
- Da Silva, J.K.R.; Maia, J.G.S.; Dosoky, N.S.; Setzer, W.N. Antioxidant, Antimicrobial, and Cytotoxic Properties of Aniba parviflora Essential Oils from the Amazon. Nat. Prod. Commun. 2016, 11, 1025–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarrazin, S.; Oliveira, R.; Maia, J.; Mourão, R. Antibacterial Activity of the Rosewood (Aniba rosaeodora and A. parviflora) Linalool-rich Oils from the Amazon. Eur. J. Med. Plants 2016, 12, 1–9. [Google Scholar] [CrossRef]
- Batista, L.; Sarrazin, S.L.; De Moura, V.; Dos Santos, I.G.; Junior, S.D.; Albuquerque, P.M. Composição química, atividade antimicrobiana e antioxidante do óleo essencial de Aniba parviflora (Meisn) Mez. Rev. Fitos 2019, 13, 181–191. [Google Scholar] [CrossRef]
- De Souza, D.P.; Pimentel, R.B.; Santos, A.S.; Albuquerque, P.M.; Fernandes, A.V.; Junior, S.D.; Oliveira, J.T.; Ramos, M.V.; Rathinasabapathi, B.; Gonçalves, J.F. Fungicidal properties and insights on the mechanisms of the action of volatile oils from Amazonian Aniba trees. Ind. Crop. Prod. 2019, 143, 111914. [Google Scholar] [CrossRef]
- May, P.H.; Barata, L.E.S. Rosewood Exploitation in the Brazilian Amazon: Options for Sustainable Production. Econ. Bot. 2004, 58, 257–265. [Google Scholar] [CrossRef]
- Lima, P.G.C.; Coelho-Ferreira, M.; Oliveira, R. Plantas medicinais em feiras e mercados públicos do Distrito Florestal Sustentável da BR-163, estado do Pará, Brasil. Acta Bot. Bras. 2011, 25, 422–434. [Google Scholar] [CrossRef] [Green Version]
- Gonzaga, A.L. Madeira: Uso e Conservação; IPHAN/Monumenta: Brasília, Brazil, 2006; p. 246. [Google Scholar]
- Marques, C.A. Importância econômica da família Lauraceae. Floram 2001, 8, 195–206. [Google Scholar]
- Lawless, J. The Encyclopedia of Essential Oils: The Complete Guide to the Use of Aromatic Oils in Aromatherapy, Herbalism, Health, and Well Being; Conari Press: San Francisco, CA, USA, 2013. [Google Scholar]
- Maia, J.G.S.; Mourão, R.H.V. Amazon Rosewood (Aniba rosaeodora Ducke) Oils. In Essential Oils in Food Preservation, Flavor and Safety; Elsevier: Amsterdam, The Netherlands, 2016; pp. 193–201. [Google Scholar] [CrossRef]
- Berg, M.E.V.D.; Silva, M.H.L. Ethnobotany of a traditional ablution in Pará, Brazil. Bol. do Mus Para Emílio Goeldi 1986, 2, 213–218. [Google Scholar]
- Price, S.; Price, L. Aromatherapy for Health Professionals, 3rd ed.; Churchill Livingstone: London, UK, 2007. [Google Scholar]
- de Moura, V.M.; Freitas-De-Sousa, L.; Dos-Santos, M.C.; Raposo, J.D.A.; Lima, A.E.; de Oliveira, R.B.; da Silva, M.N.; Mourão, R.H.V. Plants used to treat snakebites in Santarém, western Pará, Brazil: An assessment of their effectiveness in inhibiting hemorrhagic activity induced by Bothrops jararaca venom. J. Ethnopharmacol. 2015, 161, 224–232. [Google Scholar] [CrossRef]
- Corrêa, M.P. Dicionário das plantas úteis do Brasil e das exóticas cultivadas: HL, Volume 4; Ministério da Agricultura, Instituto Brasileiro de Desenvolvimento Florestal: Rio de Janeiro, Brazil, 1984. [Google Scholar]
- Martins, E.C. Plantas Medicinais de uso na Amazônia; Graficentro/Cejup: Belém, Brazil, 1889. [Google Scholar]
- Almeida, E.R. Plantas Medicinais Brasileiras: Conhecimentos Populares e Científicos, 1st ed.; Hemus Ltd.: São Paulo, Brazil, 1993. [Google Scholar]
- Pedrollo, C.T.; Kinupp, V.F.; Shepard, G.; Heinrich, M. Medicinal plants at Rio Jauaperi, Brazilian Amazon: Ethnobotanical survey and environmental conservation. J. Ethnopharmacol. 2016, 186, 111–124. [Google Scholar] [CrossRef]
- Veja, M. Etnobotánica de la Amazonia Peruana; Ediciones Aby Yala: Quito, Ecuador, 2001. [Google Scholar]
- Santos, J.; Reis, A.; Parry, S.; Leão, F.; Carvalho, J. Caracterização Etnobotânica de Essências Florestais com Fins Medicinais Utilizadas pela Etnia Xipaya, no Município de Altamira-PA. Biota Amaz. 2016, 6, 1–8. [Google Scholar] [CrossRef]
- Maia, J.G.S.; Zoghbi, M.G.B.; Andrade, E.H.A. Plantas Aromáticas na Amazônia e Seus Óleos Essenciais; Museu Emílio Goeldi: Belém, Brazil, 2001; p. 230. [Google Scholar]
- Normah, M.N.; Chin, H.F.; Reed, B.M. (Eds.) Conservation of Tropical Plant. Species, 3rd ed.; Springer: New York, NY, USA, 2013. [Google Scholar] [CrossRef]
- IUCN. The IUCN Red List of Threatened Species. Available online: https://www.iucnredlist.org (accessed on 17 January 2021).
- Martinelli, G.; Moraes, M.A. Livro Vermelho da Flora do Brasil, 1rd ed.; Andrea Jacobsson: Rio de Janeiro, Brasil, 2013; p. 1100. [Google Scholar]
- Mitja, D.; Lescure, J.-P.; Emperaire, L. Du bois pour du parfum: Le bois de rose doit-il disparaître? In Le for~ t en Jeu l’Extrativisme en Amazonie Centrale; IRD Éditions: Paris, France, 1996; pp. 93–102. [Google Scholar] [CrossRef]
- Contim, L.A.S.; Contim, L.S.R. A tecnologia produtiva do pau-rosa (Aniba rosaeodora Ducke) como aliada ao desenvolvi-mento sustentável da região amazônica. Inclusão Soc. 2018, 12, 199–207. [Google Scholar]
- IBAMA. Portaria No. 037/92-N: Lista oficial de Espécies de Flora Brasileira Ameaçadas de Extinção. 1992. Available online: https://www.mma.gov.br/estruturas/179/_arquivos/179_05122008033627.pdf (accessed on 16 August 2020).
- IBAMA. Portaria No. 01/98. 1998. Available online: http://snif.florestal.gov.br/images/pdf/legislacao/portarias/portaria_ibama_01_1998.pdf (accessed on 16 August 2020).
- Sampaio, P.T.B. Propagação Vegetativa do PAU-Rosa (Aniba rosaeodora Ducke) Pelo Método da Estaquia. Ph.D. Thesis, Universidade Federal do Amazonas, Manaus, Brazil, 1987. [Google Scholar]
- Sampaio, P.T.B.; Ferraz, I.D.K.; Camargo, J.L.C. Pau-rosa, Aniba rosaeodora Ducke. In Manual de Se-Mentes da Amazônia, 1st ed.; Ferraz, I.D.K., Ed.; INPA: Manaus, Brazil, 2003; p. 6. [Google Scholar]
- Magalhães, L.M.S.; Alencar, J.D.C. Fenologia do pau-rosa (Aniba duckei Kostermans), Lauraceae, em floresta primária na Amazônia Central. Acta Amaz. 1979, 9, 227–232. [Google Scholar] [CrossRef] [Green Version]
- Spironello, W.R.; Sampaio, P.D.T.B.; Ronchi-Teles, B. Produção e predação de frutos em Aniba rosaeodora Ducke var. amazonica Ducke (Lauraceae) em sistema de plantio sob floresta de terra firme na Amazônia Central. Acta Bot. Bras. 2004, 18, 801–807. [Google Scholar] [CrossRef]
- Jardim, L.S.; Sampaio, P.D.T.B.; Costa, S.D.S.; Gonçalves, C.D.Q.B.; Brandão, H.L.M. Efeito de diferentes reguladores de crescimento na regeneração in vitro de pau-rosa (Aniba rosaeodora Ducke). Acta Amaz. 2010, 40, 275–279. [Google Scholar] [CrossRef]
- Taveira, F.S.N. Plantas Aromáticas da Serra de Carajás Como Prováveis Marcadores Biogeoquímicos de Solos Mineralizados. Ph.D. Thesis, Universidade Federal do Pará, Belém, Brazil, 1991. [Google Scholar]
- Lima, M.D.P.; Silva, T.M.D.; Da Silva, J.D.; Zoghbi, M.D.G.B.; Andrade, E.H.A. Essential oil composition of leaf and fine stem of Aniba canelilla (Kunth) Mez from Manaus, Brazil. Acta Amaz. 2004, 34, 329–330. [Google Scholar] [CrossRef] [Green Version]
- Silva, M.D.F.D.; Taveira, F.S.; Maia, J.G.S.; Conserva, L.M.; Yoshida, M.; Gottlieb, O.R. The natural occurrence of magnosalicin diastereomers. Phytochemistry 1997, 45, 1527–1528. [Google Scholar] [CrossRef]
- Taveira, F.S.; Silva, M.D.F.D.; Maia, J.G.S. Essential Oil of Aniba puchury-minor (Mart.) Mez Occurring in the Amazon. J. Essent. Oil Bear. Plants 2003, 6, 27–30. [Google Scholar] [CrossRef]
- De Lima, W.; Rojas-Fermin, L.-B.; Koteich-Khatib, S.; Lucena, M.E.; Arzola, J.C. Volatile Constituents of the Leaves of Aniba hostmanniana (Lauraceae) and their Antibacterial Activities. Nat. Prod. Commun. 2015, 10, 1321–1322. [Google Scholar] [CrossRef] [Green Version]
- Luz, A.I.R.; Da Silva, J.D.; Zoghbi, M.D.G.B.; Andrade, E.H.D.A.; Maia, J.G.S. Essential Oil from Aniba riparia (Nees) Mez. J. Essent. Oil Res. 2002, 14, 218–219. [Google Scholar] [CrossRef]
- Alcântara, J.M.; Yamaguchi, K.K.D.L.; Junior, V.F.D.V.; Lima, E.S. Composição química de óleos essenciais de espécies de Aniba e Licaria e suas atividades antioxidante e antiagregante plaquetária. Química Nova 2010, 33, 141–145. [Google Scholar] [CrossRef]
- dos Santos, R.Q.; Maia, C.S.F.; Junior, E.A.F.; Melo, A.S.; Pinheiro, B.; Maia, J.G.S. Linalool-rich essential oils from the Amazon display antidepressant-type effect in rodents. J. Ethnopharmacol. 2018, 212, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Souza, R.D.C.Z.D. Avaliação das Frações Voláteis de Espécies de Aniba por Microextração em Fase Sólida Acoplada a Cromatografia Gasosa (SPME-GC) e Cromatografia Gasosa Bidimensional Abrangente (GC × GC). Ph.D. Thesis, Universidade Estadual de Campinas, Campinas, Brazil, 2011. Available online: http://www.repositorio.unicamp.br/handle/REPOSIP/248376 (accessed on 18 August 2019).
- Zoghbi, M.G.B.; Ohashi, S.T.; Salomão, R.O.; Guilhon, G.M.S.P. Chemical variability of Aniba rosaeodora oils. Glob. J. Sci. Front. Res. B Chem. 2015, 15, 780. [Google Scholar]
- Amazonas, D.R.; Oliveira, C.; Barata, L.E.S.; Tepe, E.J.; Kato, M.J.; Mourão, R.H.V.; Yamaguchi, L.F. Chemical and Genotypic Variations in Aniba rosiodora from the Brazilian Amazon Forest. Molecules 2020, 26, 69. [Google Scholar] [CrossRef] [PubMed]
- Andrade, E.H.A.; Zoghbi, M.D.G.B.; Maia, J.G.S. Volatiles from Aniba terminalis Ducke. J. Essent. Oil Res. 2003, 15, 81–82. [Google Scholar] [CrossRef]
- Santana, A.; Ohashi, S.; De Rosa, L.; Green, C. Brazilian rosewood oil the prospect for sustainable production and oil quality management. Int. J. Aromather. 1997, 8, 16–20. [Google Scholar] [CrossRef]
- Lupe, F.A. Estudo da Composição Química de Óleos Essenciais de Plantas Aromáticas da Amazônia. Ph.D. Thesis, Universidade Estadual de Campinas, São Paulo, Brazil, 2007. [Google Scholar]
- Fidelis, C.H.; Augusto, F.; Sampaio, P.T.; Krainovic, P.; Barata, L.E. Chemical characterization of rosewood (Aniba rosaeodora Ducke) leaf essential oil by comprehensive two-dimensional gas chromatography coupled with quadrupole mass spectrometry. J. Essent. Oil Res. 2012, 24, 245–251. [Google Scholar] [CrossRef]
- Fidelis, C.H.; Sampaio, P.T.; Krainovic, P.; Augusto, F.; Barata, L.E. Correlation between maturity of tree and GC × GC–qMS chemical profiles of essential oil from leaves of Aniba rosaeodora Ducke. Microchem. J. 2013, 109, 73–77. [Google Scholar] [CrossRef]
- Krainovic, P.M.; de Almeida, D.R.A.; Junior, V.F.D.V.; Sampaio, P.D.T.B. Changes in rosewood (Aniba rosaeodora Ducke) essential oil in response to management of commercial plantations in Central Amazonia. For. Ecol. Manag. 2018, 429, 143–157. [Google Scholar] [CrossRef]
- Teles, R.D.M.; Filho, V.E.M.; De Souza, A.G. Chemical Characterization and Larvicidal Activity of Essential Oil from Aniba duckei Kostermans against Aedes aegypti. Int. J. Life Sci. Sci. Res. 2017, 3, 1495–1499. [Google Scholar] [CrossRef]
- Everton, G.O.; Júnior, P.S.S.; Araújo, R.J.P.; Ferreira, A.M.; Gomes, P.R.B.; Rosa, P.V.S.; Pereira, A.P.M.; Filho, V.E.M. Chemical profile and antimicrobial potential of essential oils of Cymbopogon citratus (DC.) Stapf, Ocimum basilicum Linn and Aniba rosaeodora Ducke. Sci. Plena 2020, 16, 061502. [Google Scholar] [CrossRef]
- Oger, J.-M.; Richomme, P.; Guinaudeau, H.; Bouchara, J.P.; Fournet, A. Aniba canelilla (H.B.K.) Mez Essential Oil: Analysis of Chemical Constituents, Fungistatic Properties. J. Essent. Oil Res. 1994, 6, 493–497. [Google Scholar] [CrossRef]
- Taveira, F.; de Lima, W.; Andrade, E.; Maia, J. Seasonal essential oil variation of Aniba canelilla. Biochem. Syst. Ecol. 2002, 31, 69–75. [Google Scholar] [CrossRef]
- Lahlou, S.; Magalhaes, P.; de Siqueira, R.J.B.; Figueiredo, A.F.; Interaminense, L.F.L.; Maia, J.G.S.; Sousa, P.J.D.C. Cardiovascular Effects of the Essential Oil of Aniba canelilla Bark in Normotensive Rats. J. Cardiovasc. Pharmacol. 2005, 46, 412–421. [Google Scholar] [CrossRef]
- Giongo, J.L.; Vaucher, R.A.; Da Silva, A.S.; Oliveira, C.B.; de Mattos, C.B.; Baldissera, M.D.; Sagrillo, M.R.; Monteiro, S.G.; Custódio, D.L.; de Matos, M.S.; et al. Trypanocidal activity of the compounds present in Aniba canelilla oil against Trypanosoma evansi and its effects on viability of lymphocytes. Microb. Pathog. 2017, 103, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Kreutz, T.; Lucca, L.G.; Loureiro-Paes, O.A.; Teixeira, H.F.; Veiga, V.F.; Limberger, R.P.; Ortega, G.G.; Koester, L.S. Optimization, validation and application of headspace solid-phase microextraction gas chromatography for the determination of 1-nitro-2-phenylethane and methyleugenol from Aniba canelilla (H.B.K.) Mez essential oil in skin permeation samples. J. Chromatogr. A 2018, 1564, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Marcano, E.E.; Padilla-Baretic, A.; Rojas-Fermín, L.; Mora-Vivas, F.D.; Ferrer-Pereira, H. Chemical composition of wood essential oil of Aniba cinnamomiflora C. K. Allen from venezuelan andes. Colomb. For. 2016, 19, 233–238. [Google Scholar] [CrossRef] [Green Version]
- Zoghbi, M.G.B.; Andrade, E.H.A.; Santos, A.S.; Da Silva, M.H.L.; Maia, J.G.S. Constituintes voláteis de espécies de Lauraceae com ocorrência na Floresta Nacional de Caxiuanã—Melgaço—PA. In Estação Científica Ferreira Penna; Dez Anos de Pesquisa: Belém, Brazil, 2003; pp. 1–3. [Google Scholar]
- Sampaio, L.D.F.S.; Maia, J.G.S.; Parijós, A.M.; Souza, R.Z.; Barata, L.E.S. Linalool from Rosewood (Aniba rosaeodora Ducke) Oil Inhibits Adenylate Cyclase in the Retina, Contributing to Understanding its Biological Activity. Phytother. Res. 2011, 26, 73–77. [Google Scholar] [CrossRef]
- Aguilar, C.A.L.; Lima, K.R.D.S.; Manno, M.C.; Maia, J.G.S.; Neto, D.L.F.; Tavares, F.B.; Roque, T.J.L.R.; Mendonça, R.D.C.A.D.; Carmo, E.S.N.D. Rosewood (Aniba rosaeodora Ducke) oil in broiler chickens diet. Rev. Bras. Saúde Prod. Anim. 2014, 15, 108–119. [Google Scholar] [CrossRef] [Green Version]
- De Almeida, R.N.; Araújo, D.A.M.; Gonçalves, J.C.R.; Montenegro, F.C.; de Sousa, D.P.; Leite, J.R.; Mattei, R.; Benedito, M.A.C.; de Carvalho, J.G.B.; Cruz, J.S.; et al. Rosewood oil induces sedation and inhibits compound action potential in rodents. J. Ethnopharmacol. 2009, 124, 440–443. [Google Scholar] [CrossRef]
- Ferreira, A.M.; Filho, V.E.M.; Mafra, N.S.C.; Sales, E.H.; Júnior, P.S.S.; Everton, G.O. Constituintes químicos, toxicidade, potencial antioxidante e atividade larvicida frente a larvas de Aedes aegypti do óleo essencial de Aniba rosaeodora Ducke. Res. Soc. Dev. 2020, 9, e520985663. [Google Scholar] [CrossRef]
- Simić, A.; Soković, M.D.; Ristić, M.; Grujić-Jovanović, S.; Vukojevic, J.; Marin, P.D. The chemical composition of some Lauraceae essential oils and their antifungal activities. Phytotherapy Res. 2004, 18, 713–717. [Google Scholar] [CrossRef] [PubMed]
- Rosato, A.; Piarulli, M.; Corbo, F.; Muraglia, M.; Carone, A.; Vitali, M.E.; Vitali, C. In Vitro Synergistic Action of Certain Combinations of Gentamicin and Essential Oils. Curr. Med. Chem. 2010, 17, 3289–3295. [Google Scholar] [CrossRef] [PubMed]
- Chiluwal, K.; Kim, J.; Bae, S.D.; Park, C.G. Essential oils from selected wooden species and their major components as repellents and oviposition deterrents of Callosobruchus chinensis (L.). J. Asia Pac. Èntomol. 2017, 20, 1447–1453. [Google Scholar] [CrossRef]
- Soeur, J.; Marrot, L.; Perez, P.; Iraqui, I.; Kienda, G.; Dardalhon, M.; Meunier, J.-R.; Averbeck, D.; Huang, M.-E. Selective cyto-toxicity of Aniba rosaeodora essential oil towards epidermoid cancer cells through induction of apoptosis. Mutat. Res. Toxicol. Environ. Mutagen. 2011, 718, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Kizak, V.; Can, E.; Danabaş, D.; Can, S. Evaluation of anesthetic potential of rosewood (Aniba rosaeodora) oil as a new anesthetic agent for goldfish (Carassius auratus). Aquaculture 2018, 493, 296–301. [Google Scholar] [CrossRef]
- De Araujo, V.C.; Corrêa, G.C.; Maia, J.G.S.; Da Silva, M.L.; Gottlieb, O.R.; Marx, M.C.; Magalhães, M.T. Óleos essenciais da Amazonia contendo linalol. Acta Amaz. 1971, 1, 45–47. [Google Scholar] [CrossRef] [Green Version]
- Sangwan, R.S.; Farooqi, A.; Shabih, F. Regulation of essential oil production in plants. Plant. Growth Regul. 2001, 34, 3–21. [Google Scholar] [CrossRef]
- Barbosa, P.C.S.; Fernandes, K.S.; Manhães, A.P.; Carneiro, S.B.; Sampaio, P.D.T.B.; Wiedemann, L.S.M.; Junior, V.F.D.V. New and sustainable essential oils obtained from the long-term explored cinnamomum-like Aniba canelilla. J. Appl. Res. Med. Aromat. Plants 2017, 5, 60–71. [Google Scholar] [CrossRef]
- Manhães, A.P.; Da Veiga-Júnior, V.F.; Wiedemann, L.S.M.; Fernandes, K.S.; Sampaio, P.D.T.B. Biomass production and essential oil yield from leaves, fine stems and resprouts using pruning the crown of Aniba canelilla (H.B.K.) (Lauraceae) in the Central Amazon. Acta Amaz. 2012, 42, 355–362. [Google Scholar] [CrossRef] [Green Version]
- Da Cunha, L.N. Influência sazonal no teor de linalol do óleo essencial da Aniba duckei Kostermans cultivada em ambiente natural na reserva florestal Ducke. Ciênc. Nat. 2011, 33, 7–15. Available online: https://www.redalyc.org/articulo.oa?id=467546359001 (accessed on 2 September 2021).
- Da Silva, I.G.R.; De Sousa, E.M.; De Moraes, A.A.B.; De Sarges, M.D.S.R.; Cascaes, M.M.; Nascimento, L.D.D.; Andrade, E.H.D.A. Avaliação sazonal do rendimento e composição química do óleo essencial das folhas de Aniba parviflora (Meisn) Mez. (Lauraceae). Braz. J. Dev. 2020, 6, 41334–41345. [Google Scholar] [CrossRef]
- Sadgrove, N.J. Comparing essential oils from Australia’s ‘Victorian Christmas Bush’ (Prostanthera lasianthos Labill., Lamiaceae) to closely allied new species: Phenotypic plasticity and taxonomic variability. Phytochemistry 2020, 176, 112403. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, O.R.; Koketsu, M.; Magalhães, M.T.; Maia, J.G.S.; Mendes, P.H.; Da Rocha, A.I.; Da Silva, M.L.; Wilberg, V.C. Óleos essenciais da Amazônia VII. Acta Amaz. 1981, 11, 143–148. [Google Scholar] [CrossRef] [Green Version]
- Chantraine, J.-M.; Dhénin, J.-M.; Moretti, C. Chemical Variability of Rosewood (Aniba rosaeodora Ducke) Essential Oil in French Guiana. J. Essent. Oil Res. 2009, 21, 486–495. [Google Scholar] [CrossRef]
- Carović-Stanko, K.; Orlic, S.; Politeo, O.; Strikić, F.; Kolak, I.; Milos, M.; Šatović, Z. Composition and antibacterial activities of essential oils of seven Ocimum taxa. Food Chem. 2010, 119, 196–201. [Google Scholar] [CrossRef]
- Sartoratto, A.; Machado, A.L.M.; Delarmelina, C.; Figueira, G.M.; Duarte, M.C.T.; Rehder, V.L.G. Composition and antimicrobial activity of essential oils from aromatic plants used in Brazil. Braz. J. Microbiol. 2004, 35, 275–280. [Google Scholar] [CrossRef] [Green Version]
- de Lima, A.B.; Santana, M.; Cardoso, A.S.; da Silva, J.K.R.; Maia, J.G.S.; Carvalho, J.C.T.; Sousa, P.J.C. Antinociceptive activity of 1-nitro-2-phenylethane, the main component of Aniba canelilla essential oil. Phytomedicine 2009, 16, 555–559. [Google Scholar] [CrossRef]
- Vale, J.K.L.; Lima, A.B.; Pinheiro, B.G.; Cardoso, A.S.; Silva, J.K.R.; Maia, J.G.S.; de Sousa, G.E.P.; da Silva, A.B.F.; Sousa, P.J.C.; Borges, R.S. Evaluation and Theoretical Study on the Anti-inflammatory Mechanism of 1-Nitro-2-phenylethane. Planta Medica 2013, 79, 628–633. [Google Scholar] [CrossRef]
- Júnior, E.Q.D.F.; Albuquerque, P.M.; Da Silva, G.F. Estudo Fitoquímico E Análise De Fotoproteção Dos Extratos E Óleos Essenciais De Aniba canelilla (H.B.K) MEZ. J. Eng. Exact Sci. 2017, 3, 0614–0620. [Google Scholar] [CrossRef]
- de Siqueira, R.J.B.; Macedo, F.I.B.; Interaminense, L.D.F.L.; Duarte, G.P.; Magalhaes, P.; Brito, T.S.; da Silva, J.K.R.; Maia, J.G.S.; Sousa, P.J.D.C.; Leal-Cardoso, J.H.; et al. 1-Nitro-2-phenylethane, the main constituent of the essential oil of Aniba canelilla, elicits a vago-vagal bradycardiac and depressor reflex in normotensive rats. Eur. J. Pharmacol. 2010, 638, 90–98. [Google Scholar] [CrossRef]
- Brito, T.S.; Lima, F.J.; Aragão, K.S.; de Siqueira, R.J.; Sousa, P.J.; Maia, J.G.; Filho, J.D.; Lahlou, S.; Magalhães, P.J. The vasorelaxant effects of 1-nitro-2-phenylethane involve stimulation of the soluble guanylate cyclase-cGMP pathway. Biochem. Pharmacol. 2012, 85, 780–788. [Google Scholar] [CrossRef] [Green Version]
- Arruda-Barbosa, L.; Rodrigues, K.M.S.; Souza-Neto, F.D.C.V.; Duarte, G.P.; Borges, R.S.; Magalhães, P.J.C.; Lahlou, S. Vasorelaxant effects of 1-nitro-2-phenylethene in rat isolated aortic rings. Vasc. Pharmacol. 2014, 63, 55–62. [Google Scholar] [CrossRef]
- Silva, N.N.S.; Silva, J.; Alves, C.N.; Andrade, E.H.D.A.; Da Silva, J.K.R.; Maia, J.G.S. Acetylcholinesterase Inhibitory Activity and Molecular Docking Study of 1-Nitro-2-Phenylethane, the Main Constituent of Aniba canelilla Essential Oil. Chem. Biol. Drug Des. 2014, 84, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Souza, K.S.; Chaar, J.S.; Oliveira, K.M.T.; Gomes, E.O.; Portela, C.N.; Pohlit, A.M.; Quignard, E.L.J.; Nunomura, S.M.; Tadei, W.P.; Mouchrek Filho, V.E.; et al. Atividade biológica de extratos, hidrolatos e óleos voláteis de pau-rosa (Aniba duckei Kostermans) e quantificação de linalol no hidrolato de folhas. Rev. Bras. Plantas Med. 2007, 9, 1–7. [Google Scholar]
- Baldisserotto, B.; Barata, L.E.S.; Silva, A.S.; Lobato, W.F.F.; Silva, L.L.; Toni, C.; Silva, L.V.F. Anesthesia of tambaqui Colossoma macropomum (Characiformes: Serrasalmidae) with the essential oils of Aniba rosaeodora and Aniba parviflora and their major compound, linalool. Neotrop. Ichthyol. 2018, 16, e170128. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira, F.P.; Rodrigues, A.C.B.D.C.; de Lima, E.J.S.P.; Silva, V.R.; Santos, L.D.S.; da Anunciação, T.A.; Nogueira, M.L.; Soares, M.B.P.; Dias, R.B.; Rocha, C.A.G.; et al. Essential Oil from Bark of Aniba parviflora (Meisn) Mez (Lauraceae) Reduces HepG2 Cell Proliferation and Inhibits Tumor Development in a Xenograft Model. Chem. Biodivers. 2021, 18, e2000938. [Google Scholar] [CrossRef] [PubMed]
- Kohn, L.K.; Queiroga, C.L.; Martini, M.C.; Barata, L.E.; Porto, P.S.S.; Souza, L.; Arns, C.W. In vitroantiviral activity of Brazilian plants (Maytenus ilicifolia and Aniba rosaeodora) against bovine herpesvirus type 5 and avian metapneumovirus. Pharm. Biol. 2012, 50, 1269–1275. [Google Scholar] [CrossRef] [PubMed]
- De Siqueira, R.J.; Rodrigues, K.M.S.; Da Silva, M.T.B.; Junior, C.A.B.C.; Duarte, G.P.; Magalhaes, P.; dos Santos, A.A.; Maia, J.G.S.; Da Cunha, P.J.S.; Lahlou, S. Linalool-rich Rosewood Oil Induces Vago-vagal Bradycardic and Depressor Reflex in Rats. Phytother. Res. 2013, 28, 42–48. [Google Scholar] [CrossRef] [Green Version]
- Amorati, R.; Foti, M.C.; Valgimigli, L. Antioxidant Activity of Essential Oils. J. Agric. Food Chem. 2013, 61, 10835–10847. [Google Scholar] [CrossRef] [PubMed]
- Mansur, J.S.; Breder, M.V.; Mansur, M.C.A.; Azuley, R.D. Determinação do fator de proteção solar por espectrofotometria. An. Bras. Dermatol. 1986, 61, 121–124. [Google Scholar]
- Agência Nacional de Vigilância Sanitária (ANVISA). Resolução RDC no 30, de 1 de junho de 2012. Aprova o regulamento técnico Mercosul sobre Protetores Solares em Cosméticos e dá Outras Providências. In Agência Nacional de Vigilância Sanitária (ANVISA); Diário Oficial da União: Brasília, Brazil, 2012. [Google Scholar]
- Dobetsberger, C.; Buchbauer, G. Actions of essential oils on the central nervous system: An updated review. Flavour Fragr. J. 2011, 26, 300–316. [Google Scholar] [CrossRef]
- Aprotosoaie, A.C.; Hancianu, M.; Costache, I.-I.; Miron, A. Linalool: A review on a key odorant molecule with valuable biological properties. Flavour Fragr. J. 2014, 29, 193–219. [Google Scholar] [CrossRef]
- Dolabela, M.F. Triagem In Vitro Para Atividade Antitumoral e Anti Trypanossoma Cruzi de Extratos Vegetais, Produtos Naturais e Susbstâncias Sintéticas. Ph.D. Thesis, Universidade Federal de Minas Gerais, Belo Horizonte, Brazil, 1997. [Google Scholar]
- Dias, C.N.; Moraes, D.F.C. Essential oils and their compounds as Aedes aegypti L. (Diptera: Culicidae) larvicides: Review. Parasitol. Res. 2013, 113, 565–592. [Google Scholar] [CrossRef]
- Janssen, A.M.; Scheffer, J.J.C.; Svendsen, A.B. Antimicrobial Activity of Essential Oils: A 1976–1986 Literature Review. Aspects of the Test Methods. Planta Med. 1987, 53, 395–398. [Google Scholar] [CrossRef] [Green Version]
- Hili, P.; Evans, C.S.; Veness, R.G. Antimicrobial action of essential oils: The effect of dimethylsulphoxide on the activity of cinnamon oil. Lett. Appl. Microbiol. 1997, 24, 269–275. [Google Scholar] [CrossRef] [Green Version]
- Hammer, K.A.; Carson, C.; Riley, T.V. Antimicrobial activity of essential oils and other plant extracts. J. Appl. Microbiol. 1999, 86, 985–990. [Google Scholar] [CrossRef] [Green Version]
- Chao, S.C.; Young, D.G.; Oberg, C.J. Screening for Inhibitory Activity of Essential Oils on Selected Bacteria, Fungi and Viruses. J. Essent. Oil Res. 2000, 12, 639–649. [Google Scholar] [CrossRef]
- Starliper, C.E.; Ketola, H.G.; Noyes, A.D.; Schill, W.; Henson, F.G.; Chalupnicki, M.A.; Dittman, D. An investigation of the bactericidal activity of selected essential oils to Aeromonas spp. J. Adv. Res. 2014, 6, 89–97. [Google Scholar] [CrossRef] [Green Version]
- Corthouts, J.; Michiels, C.W. Inhibition of nutrient- and high pressure-induced germination of Bacillus cereus spores by plant essential oils. Innov. Food Sci. Emerg. Technol. 2016, 34, 250–258. [Google Scholar] [CrossRef]
- Regnier, T.; Combrinck, S.; Veldman, W.; Du Plooy, W. Application of essential oils as multi-target fungicides for the control of Geotrichum citri-aurantii and other postharvest pathogens of citrus. Ind. Crop. Prod. 2014, 61, 151–159. [Google Scholar] [CrossRef]
- Kong, J.-O.; Lee, S.-M.; Moon, Y.-S.; Ahn, Y.-J. Nematicidal Activity of Plant Essential Oils against Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae). J. Asia Pacific Èntomol. 2006, 9, 173–178. [Google Scholar] [CrossRef]
- Abbott, W.S. A Method of Computing the Effectiveness of an Insecticide. J. Econ. Èntomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Uniyal, A.; Tikar, S.N.; Singh, R.; Shukla, S.V.; Agrawal, O.P.; Veer, V.; Sukumaran, D. Repellent effect, knockdown study and electrophysiological responses of essential oils against Aedes aegypti. J. Entomol. Zool Stud. 2014, 2, 351–357. [Google Scholar]
- O’Neal, S.T.; Johnson, E.J.; Rault, L.C.; Anderson, T.D. Vapor delivery of plant essential oils alters pyrethroid efficacy and detoxification enzyme activity in mosquitoes. Pestic. Biochem. Physiol. 2019, 157, 88–98. [Google Scholar] [CrossRef] [Green Version]
- Sheng, Z.; Jian, R.; Xie, F.; Chen, B.; Zhang, K.; Li, D.; Chen, W.; Huang, C.; Zhang, Y.; Hu, L.; et al. Screening of larvicidal activity of 53 essential oils and their synergistic effect for the improvement of deltamethrin efficacy against Aedes albopictus. Ind. Crop. Prod. 2020, 145, 112131. [Google Scholar] [CrossRef]
- de Sousa, D.; Nóbrega, F.F.F.; Santos, C.C.M.P.; De Almeida, R.N. Anticonvulsant Activity of the Linalool Enantiomers and Racemate: Investigation of Chiral Influence. Nat. Prod. Commun. 2010, 5, 1847–1851. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Trindade, R.C.S.; Xavier, J.K.A.M.; Setzer, W.N.; Maia, J.G.S.; da Silva, J.K.R. Chemical Diversity and Therapeutic Effects of Essential Oils of Aniba Species from the Amazon: A Review. Plants 2021, 10, 1854. https://doi.org/10.3390/plants10091854
da Trindade RCS, Xavier JKAM, Setzer WN, Maia JGS, da Silva JKR. Chemical Diversity and Therapeutic Effects of Essential Oils of Aniba Species from the Amazon: A Review. Plants. 2021; 10(9):1854. https://doi.org/10.3390/plants10091854
Chicago/Turabian Styleda Trindade, Rafaela C. S., Júlia Karla A. M. Xavier, William N. Setzer, José Guilherme S. Maia, and Joyce Kelly R. da Silva. 2021. "Chemical Diversity and Therapeutic Effects of Essential Oils of Aniba Species from the Amazon: A Review" Plants 10, no. 9: 1854. https://doi.org/10.3390/plants10091854
APA Styleda Trindade, R. C. S., Xavier, J. K. A. M., Setzer, W. N., Maia, J. G. S., & da Silva, J. K. R. (2021). Chemical Diversity and Therapeutic Effects of Essential Oils of Aniba Species from the Amazon: A Review. Plants, 10(9), 1854. https://doi.org/10.3390/plants10091854