
Characterisation of a 4A QTL for Metribuzin Resistance in Wheat by Developing Near-Isogenic Lines



Abstract
:1. Introduction
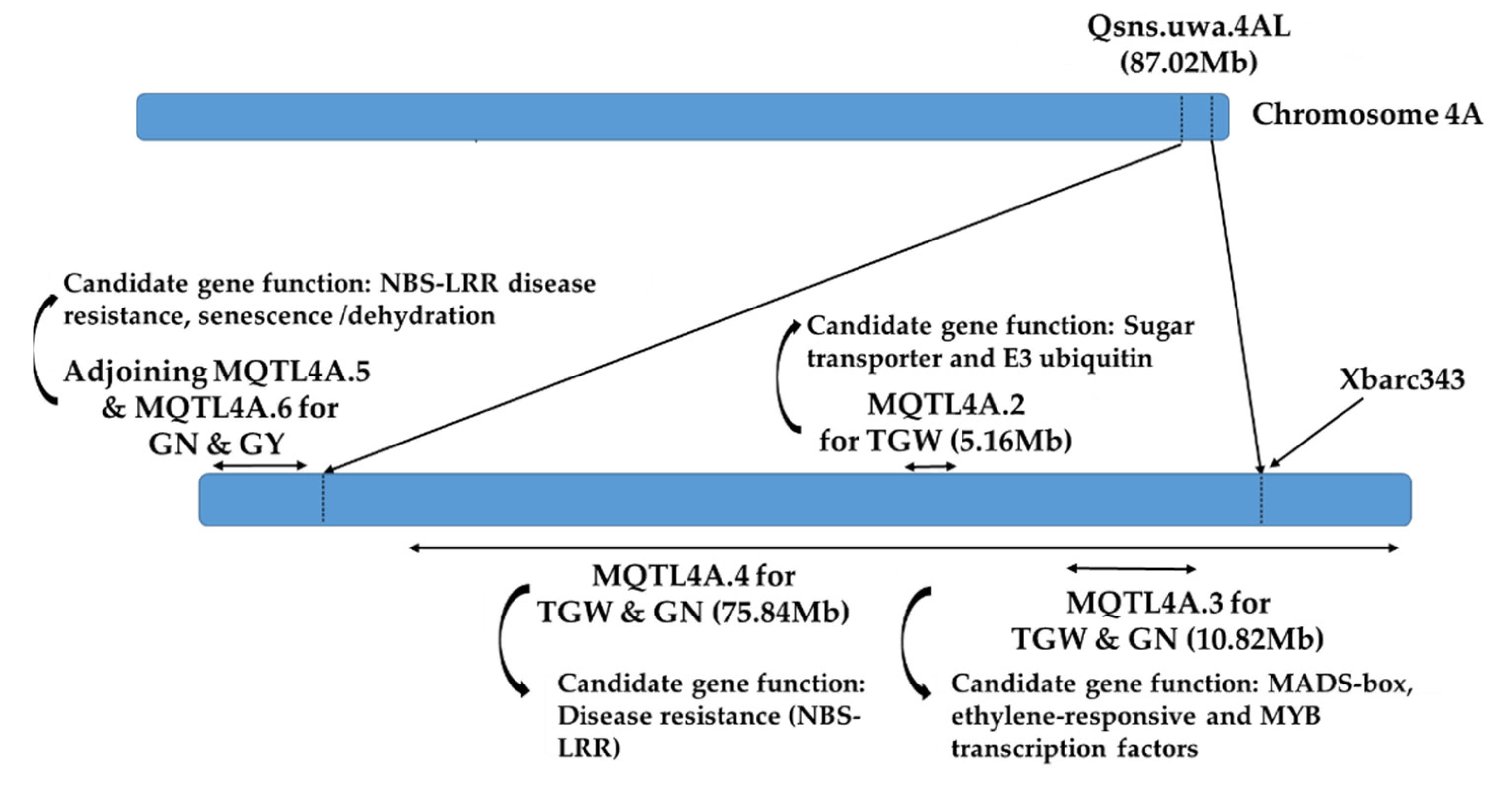
2. Results
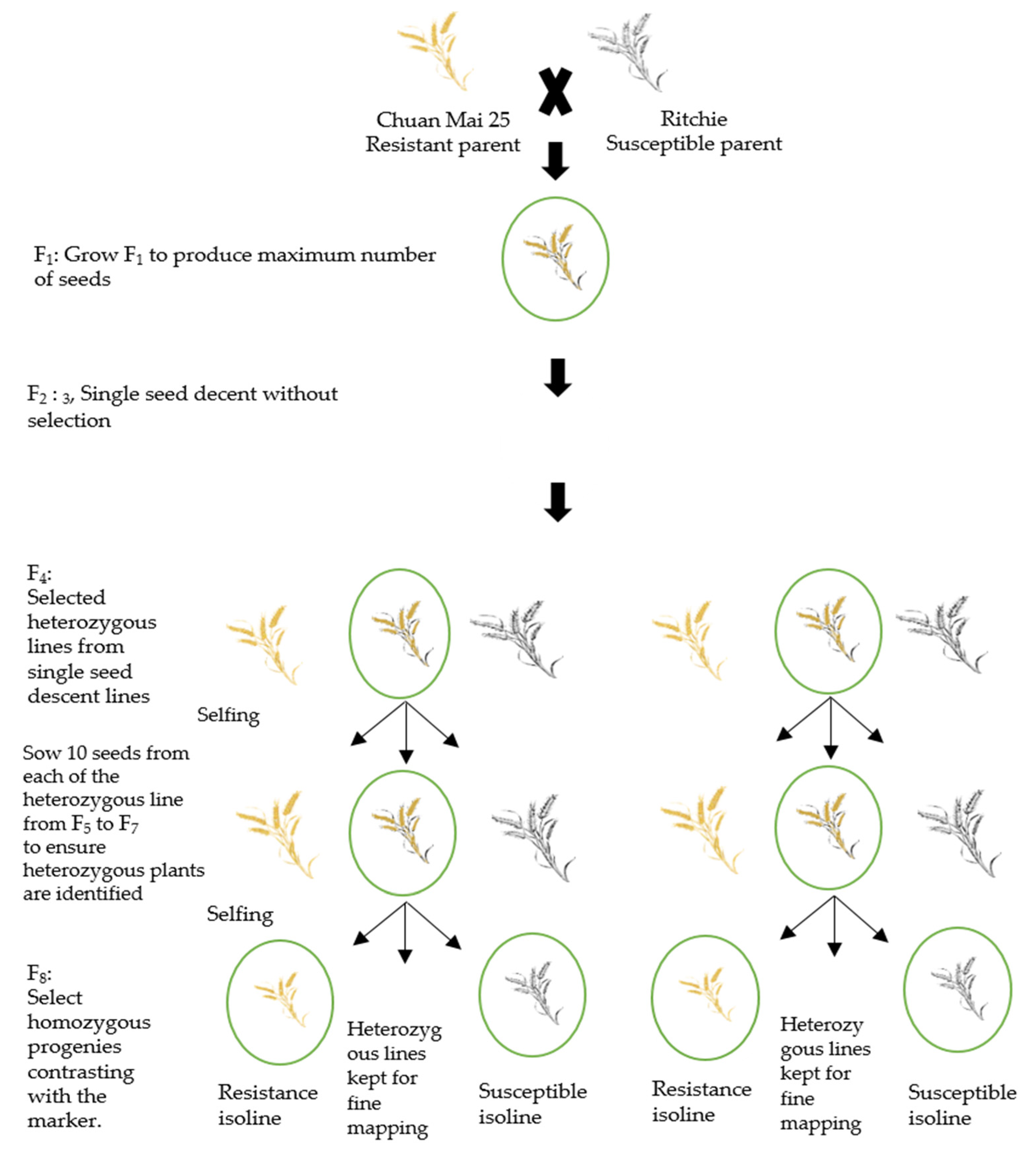
2.1. Development of Near-Isogenic Lines
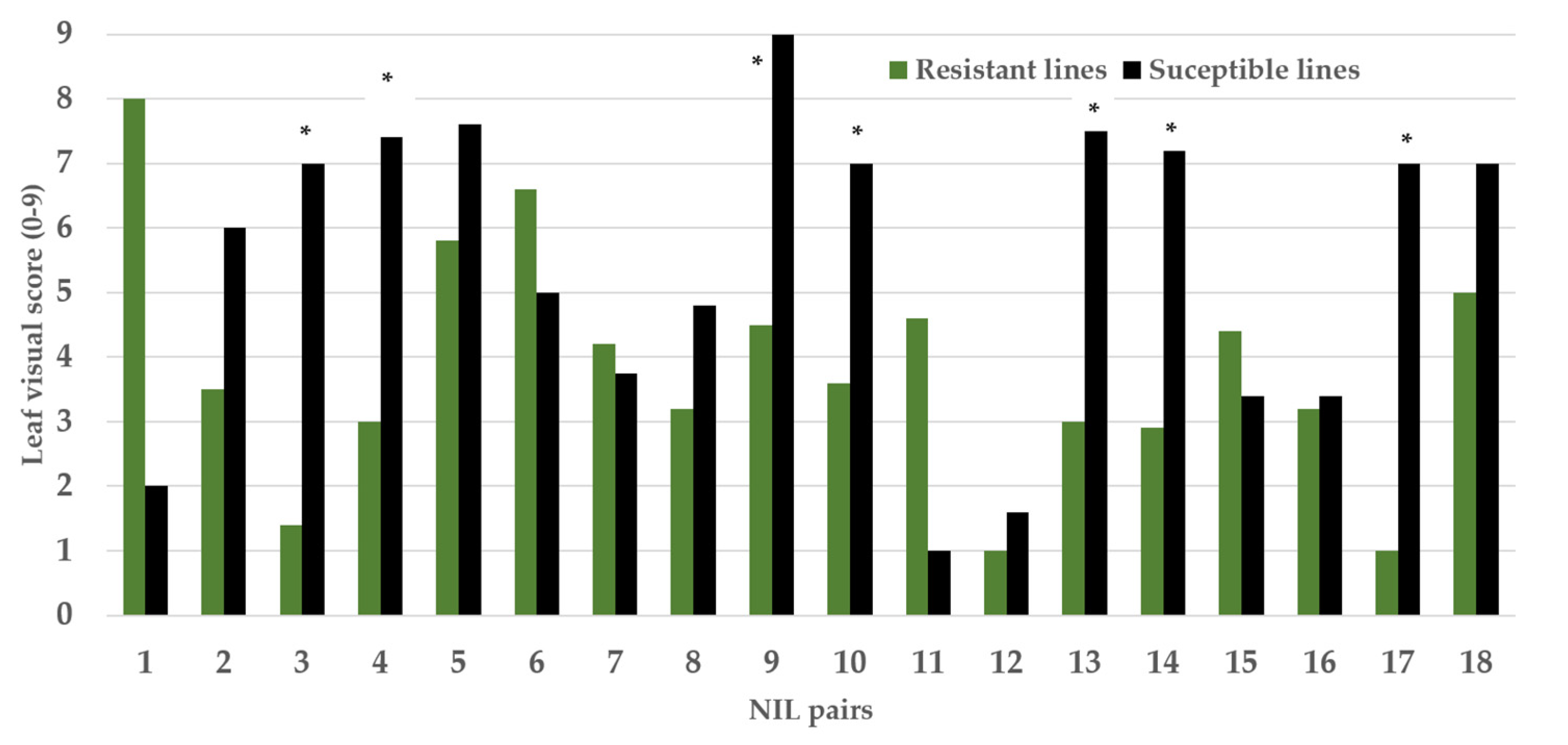
2.2. Evaluation of NIL Pairs Based on Visual Scoring (0–9) and SPAD Measurement after Metribuzin Treatment
2.3. Evaluation of NIL Pairs for Morphological Traits
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Development of NILs
4.3. Molecular Marker Analysis
4.4. Fast Generation-Cycling System
4.5. Metribuzin Assessment of Putative NILs
4.6. Phenotyping
4.7. Measurements of Other Morphological Traits
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carrubba, A.; Labruzzo, A.; Comparato, A.; Muccilli, S.; Spina, A. Use of plant water extracts for weed control in durum wheat (Triticum turgidum L. Subsp. durum Desf.). Agronomy 2020, 10, 364. [Google Scholar] [CrossRef] [Green Version]
- Kleemann, S.G.; Boutsalis, P.; Gill, G.S.; Preston, C. Applications of pre-emergent pyroxasulfone, flufenacet and their mixtures with triallate for the control of Bromus diandrus (ripgut brome) in no-till wheat (Triticum aestivum) crops of southern Australia. Crop. Prot. 2016, 80, 144–148. [Google Scholar] [CrossRef]
- Storrie, A.; McGillion, T. Herbicide resistance. In Integrated Weed Management in Australian Cropping Systems; GRDC: Canberra, Australia, 2014; pp. 28–52. [Google Scholar]
- Powles, S.B.; Gaines, T.A. Exploring the potential for a regulatory change to encourage diversity in herbicide use. Weed Sci. 2016, 64, 649–654. [Google Scholar] [CrossRef] [Green Version]
- Davis, A.; Lewis, S.; Brodie, J.; Benson, A. The potential benefits of herbicide regulation: A cautionary note for the Great Barrier Reef catchment area. Sci. Total Environ. 2014, 490, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Mets, L.; Thiel, A. Biochemistry and genetic control of the photosystem II herbicide target site. In Target Sites of Herbicide Action; CRC Press: Boca Raton, FL, USA, 2020; pp. 1–24. [Google Scholar]
- Bhoite, R.N.; Si, P.; Stefanova, K.T.; Siddique, K.H.; Yan, G. Identification of new metribuzin-tolerant wheat (Triticum spp.) genotypes. Crop. Pasture Sci. 2017, 68, 401–408. [Google Scholar] [CrossRef]
- Bhoite, R.; Onyemaobi, I.; Si, P.; Siddique, K.H.; Yan, G. Identification and validation of QTL and their associated genes for pre-emergent metribuzin tolerance in hexaploid wheat (Triticum aestivum L.). BMC Genet. 2018, 19, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Paterson, A.H. Molecular dissection of quantitative traits: Progress and prospects. Genome Res. 1995, 5, 321–333. [Google Scholar] [CrossRef] [Green Version]
- Tanksley, S.; Ganal, M.; Prince, J.; De Vicente, M.; Bonierbale, M.; Broun, P.; Fulton, T.; Giovannoni, J.; Grandillo, S.; Martin, G. High density molecular linkage maps of the tomato and potato genomes. Genetics 1992, 132, 1141–1160. [Google Scholar] [CrossRef]
- Paterson, A.H.; DeVerna, J.W.; Lanini, B.; Tanksley, S.D. Fine mapping of quantitative trait loci using selected overlapping recombinant chromosomes, in an interspecies cross of tomato. Genetics 1990, 124, 735–742. [Google Scholar] [CrossRef]
- Zheng, Z.; Ma, J.; Stiller, J.; Zhao, Q.; Feng, Q.; Choulet, F.; Feuillet, C.; Zheng, Y.-L.; Wei, Y.; Han, B. Fine mapping of a large-effect QTL conferring Fusarium crown rot resistance on the long arm of chromosome 3B in hexaploid wheat. BMC Genom. 2015, 16, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.-K.; Fu, B.-Y.; Gao, Y.-M.; Xu, J.-L.; Ali, J.; Lafitte, H.; Jiang, Y.-Z.; Rey, J.D.; Vijayakumar, C.; Maghirang, R. Genome-wide introgression lines and their use in genetic and molecular dissection of complex phenotypes in rice (Oryza sativa L.). Plant. Mol. Biol. 2005, 59, 33–52. [Google Scholar] [CrossRef] [PubMed]
- Habib, A.; Shabala, S.; Shabala, L.; Zhou, M.; Liu, C. Near-isogenic lines developed for a major QTL on chromosome arm 4HL conferring Fusarium crown rot resistance in barley. Euphytica 2016, 209, 555–563. [Google Scholar] [CrossRef]
- Chen, Z.; Zheng, Z.; Luo, W.; Zhou, H.; Ying, Z.; Liu, C. Detection of a major QTL conditioning trichome length and density on chromosome arm 4BL and development of near isogenic lines targeting this locus in bread wheat. Mol. Breed. 2021, 41, 1–9. [Google Scholar] [CrossRef]
- Mia, M.S.; Liu, H.; Wang, X.; Yan, G. Multiple near-isogenic lines targeting a QTL hotspot of drought tolerance showed contrasting performance under post-anthesis water stress. Front. Plant. Sci. 2019, 10, 271. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Liu, H.; Mia, M.S.; Siddique, K.H.; Yan, G. Development of near-isogenic lines targeting a major QTL on 3AL for pre-harvest sprouting resistance in bread wheat. Crop. Pasture Sci. 2018, 69, 864–872. [Google Scholar] [CrossRef]
- Tuinstra, M.; Ejeta, G.; Goldsbrough, P. Heterogeneous inbred family (HIF) analysis: A method for developing near-isogenic lines that differ at quantitative trait loci. Theor. Appl. Genet. 1997, 95, 1005–1011. [Google Scholar] [CrossRef]
- Liu, H.; Zwer, P.; Wang, H.; Liu, C.; Lu, Z.; Wang, Y.; Yan, G. A fast generation cycling system for oat and triticale breeding. Plant. Breed. 2016, 135, 574–579. [Google Scholar] [CrossRef]
- Yan, G.; Liu, H.; Wang, H.; Lu, Z.; Wang, Y.; Mullan, D.; Hamblin, J.; Liu, C. Accelerated generation of selfed pure line plants for gene identification and crop breeding. Front. Plant. Sci. 2017, 8, 1786. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Liu, H.; Kilian, A.; Bhoite, R.; Liu, G.; Si, P.; Wang, J.; Zhou, W.; Yan, G. QTL mapping using a high-density genetic map to identify candidate genes associated with metribuzin tolerance in hexaploid wheat (Triticum aestivum L.). Front. Plant. Sci. 2020, 11, 1440. [Google Scholar] [CrossRef]
- Araus, J.L.; Amaro, T.; Zuhair, Y.; Nachit, M. Effect of leaf structure and water status on carbon isotope discrimination in field-grown durum wheat. Plant. Cell Environ. 1997, 20, 1484–1494. [Google Scholar] [CrossRef]
- Pfister, K.; Arntzen, C.J. The mode of action of photosystem II-specific inhibitors in herbicide-resistant weed biotypes. Z. Für Nat. Schung C 1979, 34, 996–1009. [Google Scholar] [CrossRef]
- Bhoite, R.; Si, P.; Siddique, K.H.; Yan, G. Genes, Mechanisms and Novel EST-SSR Markers Associated with Metribuzin Tolerance in Wheat (Triticum aestivum L.): Targets for Improving Photosynthetic Capacity and Yield; Research Square: Durham, NC, USA, 2020. [Google Scholar]
- Sharma, A.; Kumar, V.; Shahzad, B.; Ramakrishnan, M.; Sidhu, G.P.S.; Bali, A.S.; Handa, N.; Kapoor, D.; Yadav, P.; Khanna, K. Photosynthetic response of plants under different abiotic stresses: A review. J. Plant. Growth Regul. 2019, 1–23. [Google Scholar] [CrossRef]
- Collard, B.C.; Mackill, D.J. Marker-assisted selection: An approach for precision plant breeding in the twenty-first century. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 557–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venuprasad, R.; Dalid, C.; Del Valle, M.; Zhao, D.; Espiritu, M.; Cruz, M.S.; Amante, M.; Kumar, A.; Atlin, G. Identification and characterization of large-effect quantitative trait loci for grain yield under lowland drought stress in rice using bulk-segregant analysis. Theor. Appl. Genet. 2009, 120, 177–190. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, Y.; Chen, G.; Tian, J. Genetic analysis of grain yield and leaf chlorophyll content in common wheat. Cereal Res. Commun. 2009, 37, 499–511. [Google Scholar] [CrossRef]
- Liu, H.; Mullan, D.; Zhang, C.; Zhao, S.; Li, X.; Zhang, A.; Lu, Z.; Wang, Y.; Yan, G. Major genomic regions responsible for wheat yield and its components as revealed by meta-QTL and genotype–phenotype association analyses. Planta 2020, 252, 1–22. [Google Scholar] [CrossRef]
- Daws, M.; Hall, J.; Flynn, S.; Pritchard, H. Do invasive species have bigger seeds? Evidence from intra-and inter-specific comparisons. South. Afr. J. Bot. 2007, 73, 138–143. [Google Scholar] [CrossRef] [Green Version]
- Ma, M.; Zhao, H.; Li, Z.; Hu, S.; Song, W.; Liu, X. TaCYP78A5 regulates seed size in wheat (Triticum aestivum). J. Exp. Bot. 2015, 67, 1397–1410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, M.; Wang, Q.; Li, Z.; Cheng, H.; Li, Z.; Liu, X.; Song, W.; Appels, R.; Zhao, H. Expression of Ta CYP 78A3, a gene encoding cytochrome P450 CYP 78A3 protein in wheat (Triticum aestivum L.), affects seed size. Plant. J. 2015, 83, 312–325. [Google Scholar] [CrossRef] [PubMed]
- Daws, M.I.; Pearson, T.R.; Burslem, D.F.P.; Mullins, C.E.; Dalling, J.W. Effects of topographic position, leaf litter and seed size on seedling demography in a semi-deciduous tropical forest in Panama. Plant. Ecol. 2005, 179, 93–105. [Google Scholar] [CrossRef]
- Leishman, M.R.; Wright, I.J.; Moles, A.T.; Westoby, M. The evolutionary ecology of seed size. Seeds Ecol. Regen. Plant. Communities 2000, 2, 31–57. [Google Scholar]
- Maity, A.; Singh, V.; Jessup, R.; Bagavathiannan, M. Seed traits correlate with herbicide resistance in Italian ryegrass (Lolium perenne ssp. multiflorum). Pest. Manag. Sci. 2021, 77, 2756–2765. [Google Scholar] [CrossRef] [PubMed]
- Courtois, B.; Ahmadi, N.; Khowaja, F.; Price, A.H.; Rami, J.-F.; Frouin, J.; Hamelin, C.; Ruiz, M. Rice root genetic architecture: Meta-analysis from a drought QTL database. Rice 2009, 2, 115–128. [Google Scholar] [CrossRef] [Green Version]
- Murray, M.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- MacFarland, T.W.; Yates, J.M. Mann–whitney u test. In Introduction to Nonparametric Statistics for the Biological Sciences Using R; Springer: Basel, Switzerland, 2016; pp. 103–132. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NIL Pairs as per Figure 2 | Line Name (QTL Reference: [8]) | Leaf Visual Score (0–9) | Leaf Visual Score (0–9) Difference in Pairs (%) | SPAD | SPAD Difference in Pairs (%) |
|---|---|---|---|---|---|
| 3 | Qsns.uwa.4AL.3R | 1.4 ± 0.07 ** | 68 | 27 ± 0.64 ** | 26 |
| Qsns.uwa.4AL.3S | 7.0 ± 0.18 | 15 ± 1.29 | |||
| 4 | Qsns.uwa.4AL.4R | 3.0 ± 0.28 ** | 69 | 28 ± 1.82 * | 26 |
| Qsns.uwa.4AL.4S | 7.4 ± 0.27 | 22 ± 0.85 | |||
| 9 | Qsns.uwa.4AL.9R | 4.5 ± 0.18 * | 85 | 22 ± 0.91 * | 21 |
| Qsns.uwa.4AL.9S | 9.0 ± 0.00 | 14 ± 1.31 | |||
| 10 | Qsns.uwa.4AL.10R | 3.6 ± 0.17 ** | 65 | 23 ± 0.57 ** | 22 |
| Qsns.uwa.4AL.10S | 7.0 ± 0.13 | 12 ± 0.81 | |||
| 13 | Qsns.uwa.4AL.13R | 3.0 ± 0.25 * | 63 | 30 ± 1.31 ** | 29 |
| Qsns.uwa.4AL.13S | 7.5 ± 0.22 | 20 ± 1.03 | |||
| 14 | Qsns.uwa.4AL.14R | 2.9 ± 0.37 ** | 68 | 25 ± 1.65 * | 24 |
| Qsns.uwa.4AL.14S | 7.2 ± 0.25 | 20 ± 0.75 | |||
| 17 | Qsns.uwa.4AL.17R | 1.0 ± 0.00 ** | 69 | 27 ± 1.31 ** | 26 |
| Qsns.uwa.4AL.17S | 7.0 ± 0.22 | 20 ± 1.10 |
| NIL Pairs | TGW (g) | Biomass (kg)/Plant | Plant Height (cm) | Tillers /Plant | Yield (g) /Plant | PM Visual Score (0–9) |
|---|---|---|---|---|---|---|
| 3R | 26.7 ± 0.35 ** | 0.45 ± 0.03 | 64 ± 0.33 ** | 4.0 ± 0.40 * | 10.50 ± 1.00 | 1.0 ± 0.00 |
| 3S | 23.4 ± 0.10 | 0.65 ± 0.02 | 59 ± 0.57 | 6.5 ± 0.28 | 11.70 ± 0.15 | 1.0 ± 0.00 |
| 4R | 34.1 ± 0.05 ** | 0.55 ± 0.05 | 60 ± 0.30 | 3.5 ± 0.28 | 6.75 ± 2.75 ** | 1.0 ± 0.00 ** |
| 4S | 29.1 ± 0.15 | 0.63 ± 0.01 | 60 ± 0.00 | 4.5 ± 0.28 | 21.20 ± 1.10 | 7.5 ± 0.50 |
| 9R | 38.3 ± 0.05 * | 0.65 ± 0.05 * | 63 ± 0.57 | 6.5 ± 0.28 ** | 31.60 ± 0.35 * | 1.0 ± 0.00 ** |
| 9S | 36.4 ± 0.20 | 0.48 ± 0.03 | 63 ± 1.73 | 3.0 ± 0.00 | 21.60 ± 0.40 | 7.0 ± 0.00 |
| 10R | 35.2 ± 0.05 ** | 0.50 ± 0.02 ** | 64 ± 0.33 | 4.0 ± 0.40 | 16.50 ± 1.00 * | 1.0 ± 0.00 ** |
| 10S | 30.4 ± 0.30 | 0.29 ± 0.05 | 64 ± 0.57 | 2.0 ± 0.00 | 11.20 ± 1.15 | 7.5 ± 0.00 |
| 13R | 40.1 ± 0.05 ** | 0.69 ± 0.04 ** | 65 ± 0.33 | 6.5 ± 0.28 * | 20.30 ± 0.25 * | 1.0 ± 0.00 |
| 13S | 36.2 ± 0.10 | 0.40 ± 0.02 | 67 ± 0.33 | 3.5 ± 0.28 | 13.20 ± 0.20 | 1.0 ± 0.00 |
| 14R | 28.6 ± 0.10 * | 0.65 ± 0.02 ** | 67 ± 0.57 ** | 6.5 ± 0.28 ** | 15.20 ± 0.16 | 1.0 ± 0.00 |
| 14S | 30.4 ± 0.20 | 0.26 ± 0.02 | 74 ± 1.15 | 2.0 ± 0.00 | 14.70 ± 0.85 | 1.0 ± 0.00 |
| 17R | 37.5 ± 0.25 ** | 0.35 ± 0.01 | 62 ± 1.15 ** | 3.0 ± 0.40 | 30.10 ± 0.08 | 1.0 ± 0.00 |
| 17S | 27.5 ± 0.25 | 0.33 ± 0.03 | 69 ± 0.57 | 2.0 ± 0.00 | 31.60 ± 1.15 | 1.0 ± 0.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhattarai, R.; Liu, H.; Siddique, K.H.M.; Yan, G. Characterisation of a 4A QTL for Metribuzin Resistance in Wheat by Developing Near-Isogenic Lines. Plants 2021, 10, 1856. https://doi.org/10.3390/plants10091856
Bhattarai R, Liu H, Siddique KHM, Yan G. Characterisation of a 4A QTL for Metribuzin Resistance in Wheat by Developing Near-Isogenic Lines. Plants. 2021; 10(9):1856. https://doi.org/10.3390/plants10091856
Chicago/Turabian StyleBhattarai, Rudra, Hui Liu, Kadambot H. M. Siddique, and Guijun Yan. 2021. "Characterisation of a 4A QTL for Metribuzin Resistance in Wheat by Developing Near-Isogenic Lines" Plants 10, no. 9: 1856. https://doi.org/10.3390/plants10091856
APA StyleBhattarai, R., Liu, H., Siddique, K. H. M., & Yan, G. (2021). Characterisation of a 4A QTL for Metribuzin Resistance in Wheat by Developing Near-Isogenic Lines. Plants, 10(9), 1856. https://doi.org/10.3390/plants10091856