
Polyploidization Increases the Lipid Content and Improves the Nutritional Quality of Rice

Abstract
:1. Introduction
2. Materials and Methods
2.1. Rice Samples
2.2. Reagents and Instruments
2.3. Sample Preparation and Extraction
2.4. HPLC Conditions and ESI-Q TRAP-MS/MS
2.5. Qualitative and Quantitative Analysis of Metabolites
2.6. Metabolite Data Analysis
3. Results
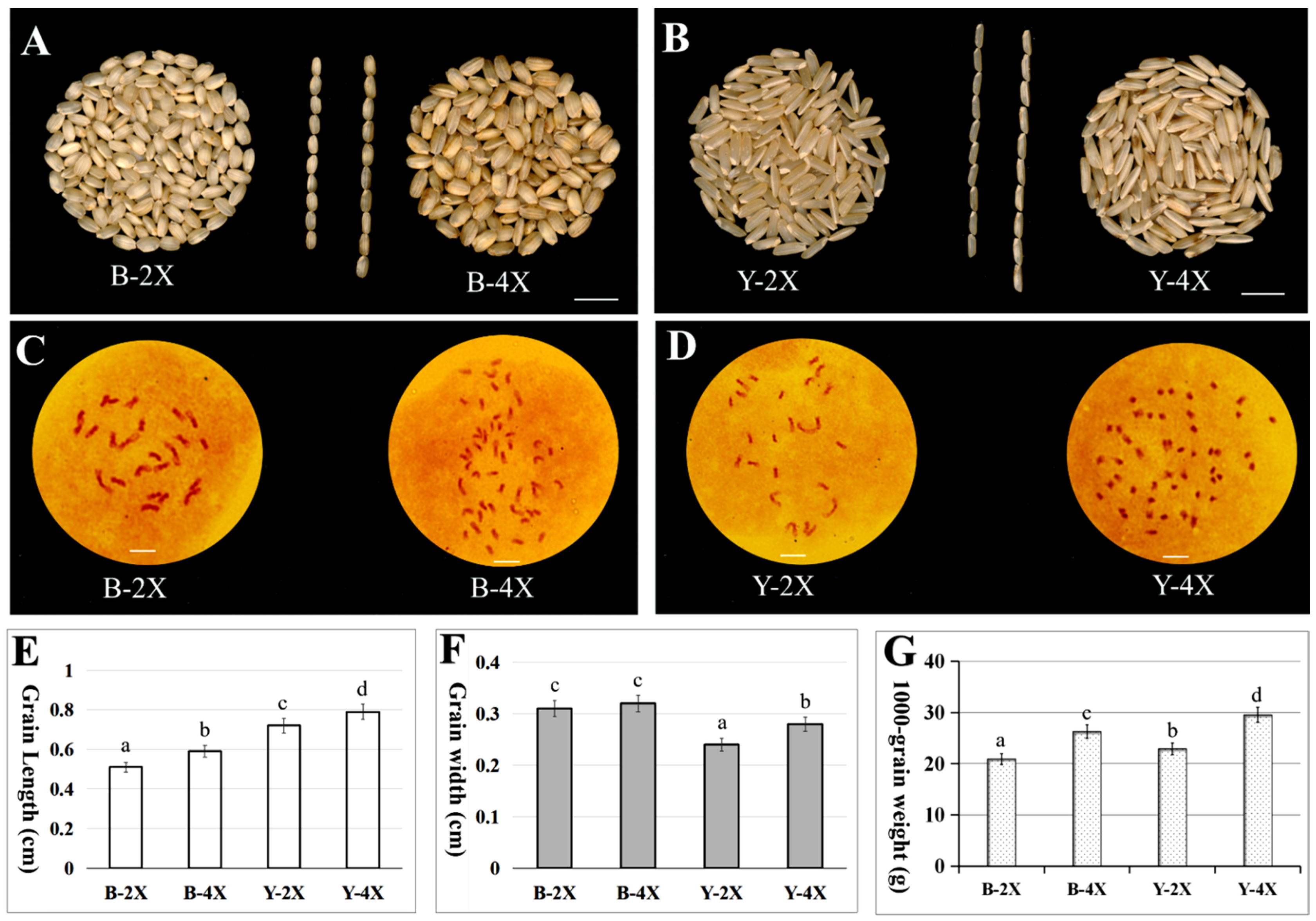
3.1. Phenotype Comparison and Chromosome Identification of Diploid and Tetraploid Rice
3.2. Widely Targeted Metabolic Profiling of Diploid and Tetraploid Brown Rice Based on LC-MS/MS
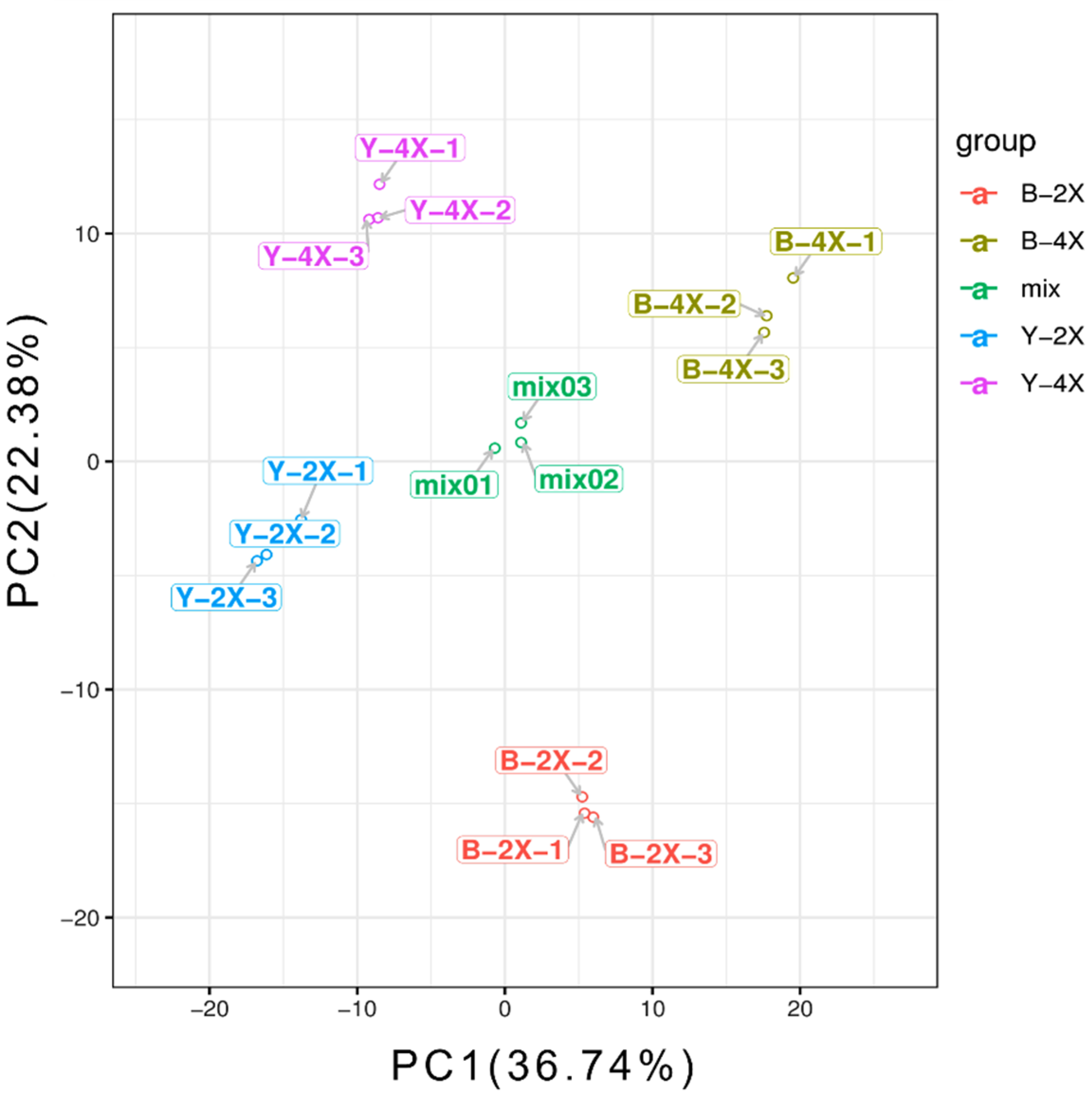
3.3. PCA for Diploid–Tetraploid Groups
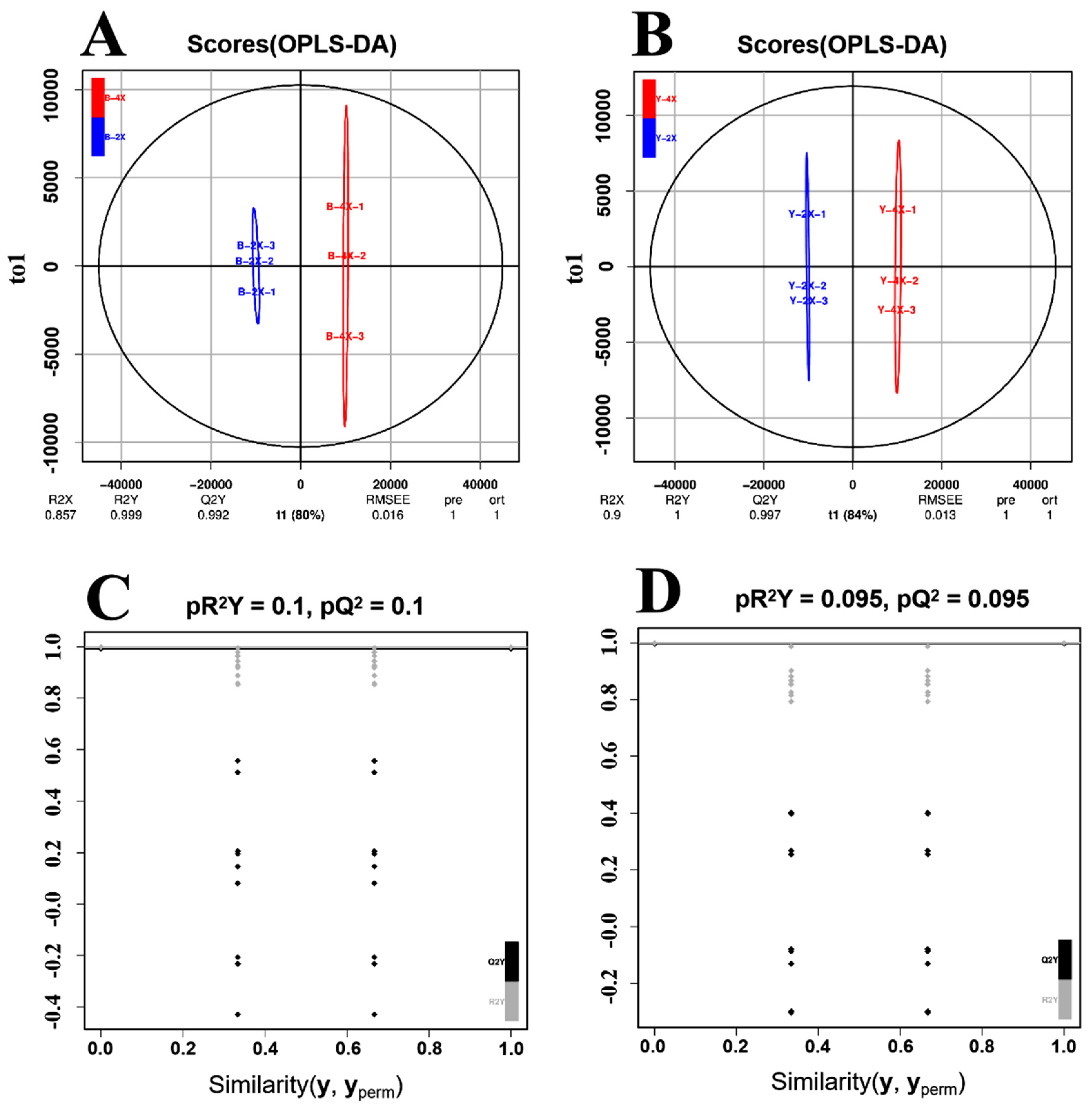
3.4. Orthogonal Projections to Latent Structures–Discriminant Analysis (OPLS-DA) for Diploid vs. Tetraploid Groups
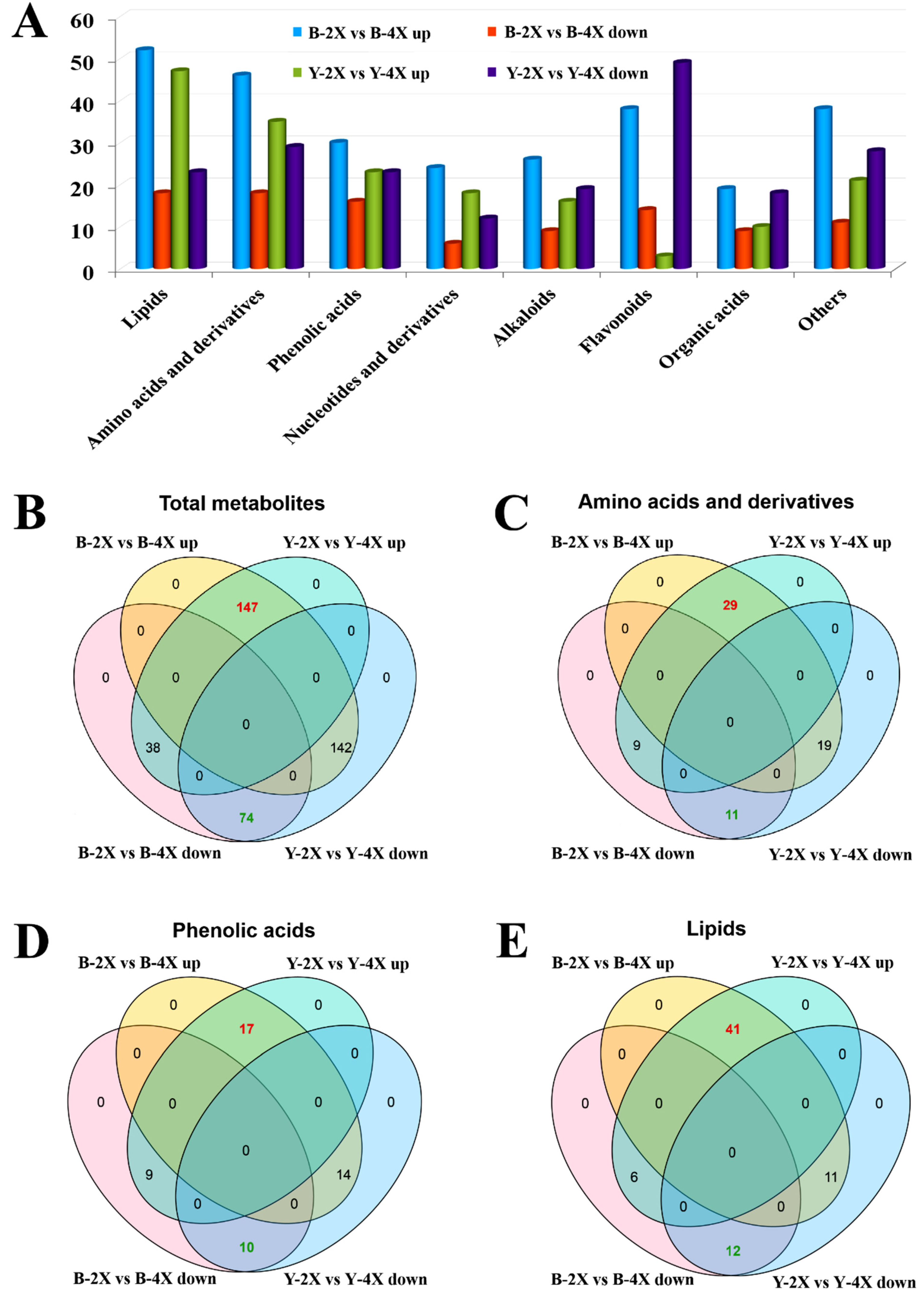
3.5. HCA and Volcano Plot of Differential Metabolites for Diploid vs. Tetraploid Groups
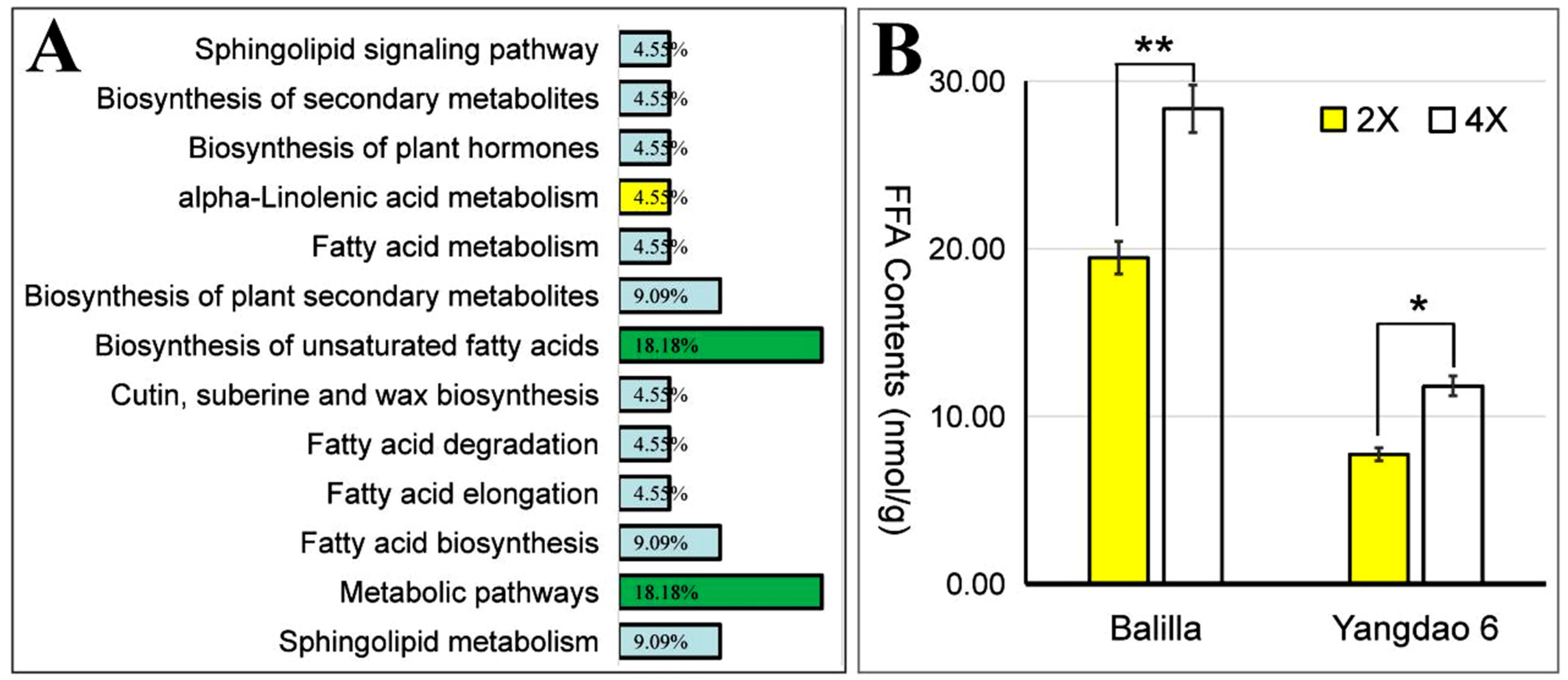
3.6. Clustering, Pathway, and Enrichment Analyses of Lipid Metabolites for Diploid vs. Tetraploid Groups
3.7. Statistical Analysis of Significant Differential Lipid Metabolites for Diploid vs. Tetraploid Groups
4. Discussion
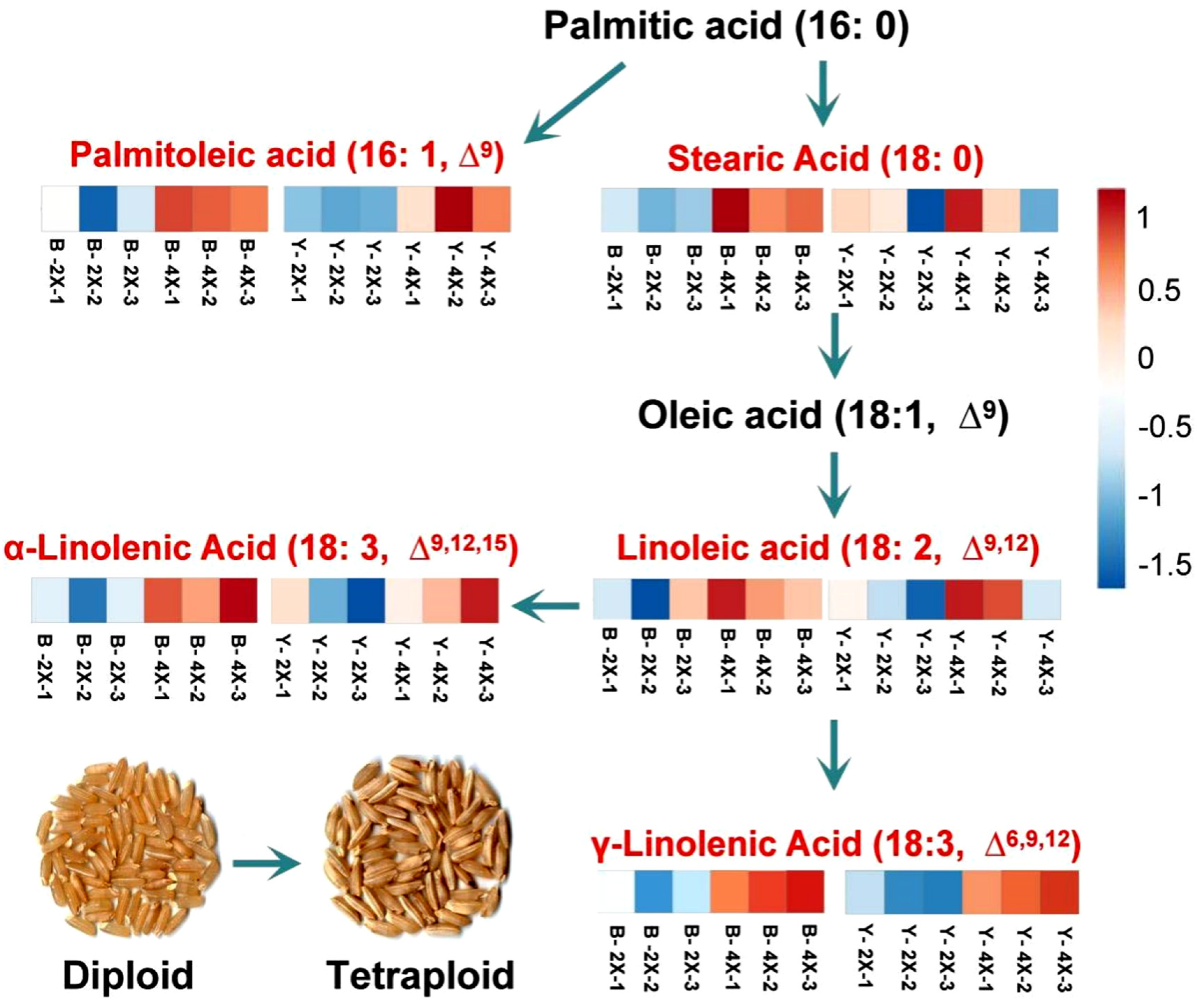
4.1. Differential Expression of Metabolites in Diploid and Tetraploid Rice
4.2. Application Potential of Polyploid Rice in Functional Rice Breeding
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Q.; Li, J.; Xue, Y.; Han, B.; Deng, X.W. Rice 2020: A call for an international coordinated effort in rice functional genomics. Mol. Plant 2008, 5, 715–719. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, H.; Tomiyama, Y.; Mizushina, Y. Lipid components, fatty acids and triacylglycerol molecular species of black and red rices. Food Chem. 2010, 123, 210–215. [Google Scholar] [CrossRef]
- Kim, N.; Kwak, J.; Baik, J.; Yoon, M.; Lee, J.; Yoon, S.; Kim, I. Changes in lipid substances in rice during grain development. Phytochemistry 2015, 116, 170–179. [Google Scholar] [CrossRef]
- Liu, L.; Waters, D.; Rose, T.; Bao, J.; King, G. Phospholipids in rice: Significance in grain quality and health benefits: A review. Food Chem. 2013, 139, 1133–1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juliano, B. Lipids in rice and rice processing. In Lipids in Cereal Technology; Academic Press: Cambridge, MA, USA, 1983; pp. 305–330. [Google Scholar] [CrossRef]
- Juliano, B. Production and Utilization of Rice; American Association of Cereal Chemists: St. Paul, MN, USA, 1985; pp. 1–16. [Google Scholar]
- Morrison, W. Starch lipids and how they relate to starch granule structure and functionality. Cereal Foods World 1995, 40, 437–446. [Google Scholar]
- Friedman, M. Rice brans, rice bran oils, and rice hulls: Composition, food and industrial uses, and bioactivities in humans, animals, and cells. J. Agric. Food Chem. 2013, 61, 10626–10641. [Google Scholar] [CrossRef]
- Yu, L.; Li, G.; Li, M.; Xu, F.; Trust, B. Genotypic variation in phenolic acids vitamin E and fatty acids in whole grain rice. Food Chem. 2016, 197, 776–782. [Google Scholar] [CrossRef]
- Zhou, Z.; Blanchard, C.; Helliwell, S. Fatty acid composition of three rice varieties following storage. J. Cereal Sci. 2003, 37, 327–335. [Google Scholar] [CrossRef]
- Kitta, K.; Ebihara, M.; lizuka, T.; Yoshikawa, R.; Isshiki, K.; Kawamoto, S. Variations in lipid content and fatty acid composition of major non-glutinous rice cultivars in Japan. J. Food Compos. Anal. 2005, 18, 269–278. [Google Scholar] [CrossRef]
- Rusydi, M.; Noraliza, C.; Azrina, A.; Zulkhaini, A. Nutritional changes in germinated legumes and rice varieties. Int. Food Res. J. 2011, 18, 688–696. [Google Scholar]
- Yoon, M.; Lee, S.; Kang, M. The lipid composition of rice cultivars with different eating qualities. J. Korean Soc. Appl. Biol. Chem. 2012, 55, 291–295. [Google Scholar] [CrossRef]
- Sharif, M.; Butt, M.; Anjum, F.; Khan, S. Rice bran: A novel functional ingredient. Crit. Rev. Food Sci. Nutr. 2014, 54, 807–816. [Google Scholar] [CrossRef] [PubMed]
- Concepcion, J.; Ouk, S.; Riede, A.; Calingacion, M.; Zhao, D.; Ouk, M.; Garson, J.; Fitzgerald, M. Quality evaluation, fatty acid analysis and untargeted profiling of volatiles in Cambodian rice. Food Chem. 2018, 240, 1014–1021. [Google Scholar] [CrossRef] [PubMed]
- Yoon, M.; Koh, H.; Kang, M. Variation of properties of lipid components in rice endosperm affected on palatability. J. Korean Soc. Appl. Biol. Chem. 2008, 51, 207–211. [Google Scholar]
- Ryan, D.; Kendall, M.; Robards, K. Bioactivity of oats as it relates to cardiovascular disease. Nutr. Res. Rev. 2007, 20, 147–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolfe, K. Yesterday’s polyploids and the mystery of diploidization. Nat. Rev. Genet. 2001, 2, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Soltis, P. Ancient and recent polyploidy in angiosperms. New Phytol. 2005, 166, 5–8. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Wickett, N.; Ayyampalayam, S.; Chanderbali, A.; Landherr, L.; Ralph, P.; Tomsho, L.; Hu, Y.; Liang, H.; Soltis, P.; et al. Ancestral polyploidy in seed plants and angiosperms. Nature 2011, 473, 97–113. [Google Scholar] [CrossRef] [PubMed]
- Peer, Y.; Mizrachi, E.; Marchal, K. The evolutionary significance of polyploidy. Nat. Rev. Genet. 2017, 18, 411–424. [Google Scholar] [CrossRef]
- Paterson, A.; Bowers, J.; Chapman, B. Ancient polyploidization predating divergence of the cereals, and its consequences for comparative genomics. Proc. Natl. Acad. Sci. USA 2004, 101, 9903–9908. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Wang, J.; Lin, W.; Li, S.; Li, H.; Zhou, J.; Ni, P.; Dong, W.; Hu, S.; Zeng, C.; et al. The genomes of Oryza sativa: A history of duplications. PLoS Biol. 2005, 3, e38. [Google Scholar] [CrossRef] [Green Version]
- Rice, A.; Smarda, P.; Novosolov, M.; Drori, M.; Glick, L.; Sabath, N.; Meiri, S.; Belmaker, J.; Mayrose, I. The global biogeography of polyploid plants. Nat. Ecol. Evol. 2019, 3, 265–273. [Google Scholar] [CrossRef]
- Song, W.; Zhang, Y. Rice tetraploid and its effects on agronomic traits and nutritional components. Acta Agron. Sin. 1992, 18, 137–144. [Google Scholar] [CrossRef]
- Tu, S.; Kong, F.; Xu, Q.; He, T. Study on new system of heterosis utilization in autotetraploid rice. Bull. Chin. Acad. Sci. 2003, 6, 426–428. [Google Scholar]
- Lu, Z.; Guo, X.; Huang, X.; Xia, J.; Li, X.; Wu, J.; Yu, H.; Shahid, M.; Liu, X. Transcriptome and gene editing analyses reveal MOF1a defect alters the expression of genes associated with tapetum development and chromosome behavior at meiosis stage resulting in low pollen fertility of tetraploid rice. Int. J. Mol. Sci. 2020, 21, 7489. [Google Scholar] [CrossRef] [PubMed]
- Cai, D.; Chen, J.; Chen, D.; Dai, B.; Zhang, W.; Song, Z.; Yang, Z.; Du, C.; Tang, Z.; He, Y.; et al. The breeding of two polyploid rice lines with the characteristic of polyploid meiosis stability. Sci. China Ser. C 2007, 50, 356–366. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Liu, X. The Research on Autotetraploid Rice; South China University of Technology Press: Guangzhou, China, 2014; pp. 90–92. [Google Scholar]
- Guo, H.; Mendrikahy, J.; Xie, L.; Deng, J.; Lu, Z.; Wu, J.; Li, X.; Shahid, M.; Liu, X. Transcriptome analysis of neo-tetraploid rice reveals specific differential gene expressions associated with fertility and heterosis. Sci. Rep. 2017, 7, 40139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, H.; Shahid, M.; Zhao, J.; Li, Y.; Wang, L.; Liu, X. Agronomic traits and cytogenetic evaluation of newly developed autotetraploid rice line. Pak. J. Agric. Sci. 2016, 53, 291–301. [Google Scholar]
- Yu, H.; Shahid, M.; Li, Q.; Li, Y.; Li, C.; Lu, Z.; Wu, J.; Zhang, Z.; Liu, X. Production assessment and genome comparison revealed high yield potential and novel specific alleles associated with fertility and yield in neotetraploid rice. Rice 2020, 13, 32. [Google Scholar] [CrossRef]
- Ghaleb, M.; Li, C.; Shahid, M.; Yu, H.; Liang, J.; Chen, R.; Wu, J.; Liu, X. Heterosis analysis and underlying molecular regulatory mechanism in a wide-compatible neo-tetraploid rice line with long panicles. BMC Plant Biol. 2020, 20, 83. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Li, Q.; Li, Y.; Yang, H.; Lu, Z.; Wu, J.; Zhang, Z.; Shahid, M.; Liu, X. Genomics analyses reveal unique classification, population structure and novel allele of neo-tetraploid rice. Rice 2021, 14, 16. [Google Scholar] [CrossRef]
- Chen, R.; Feng, Z.; Zhang, X.; Song, Z.; Cai, D. A new way of rice breeding: Polyploid rice breeding. Plants 2021, 10, 422. [Google Scholar] [CrossRef]
- Dhawan, O.; Lavania, U. Enhancing the productivity of secondary metabolites via induced polyploidy: A review. Euphytica 1996, 87, 81–89. [Google Scholar] [CrossRef]
- Xie, Z.; Mou, C.; Wang, B.; Wu, X.; Wang, X. Plant polyploidization and its appliance in breeding. Chin. Agric. Sci. Bull. 2002, 18, 70–76. [Google Scholar] [CrossRef]
- Luo, X.; He, H. Progress on polyploid rice breeding. Chin. Agric. Sci. Bull. 2001, 17, 53–56. [Google Scholar]
- Gan, L.; Huang, B.; Song, Z.; Zhang, Y.; Zhang, Y.; Chen, S.; Tong, L.; Wei, Z.; Yu, L.; Luo, X.; et al. Unique glutelin expression patterns and seed endosperm structure facilitate glutelin accumulation in polyploid rice seed. Rice 2021, 14, 61. [Google Scholar] [CrossRef] [PubMed]
- Ulaszewska, M.; Weinert, C.; Trimigno, A.; Portmann, P.; Lacueva, C.; Badertscher, R.; Brennan, L.; Brunius, C.; Bub, A.; Capozzi, F.; et al. Nutrimetabolomics: An integrative action for metabolomic analyses in human nutritional studies. Mol. Nutr. Food Res. 2018, 63, e1800384. [Google Scholar] [CrossRef] [PubMed]
- Weckwerth, W. Metabolomics in systems biology. Annu. Rev. Plant. Biol. 2003, 54, 669–689. [Google Scholar] [CrossRef] [PubMed]
- Dettmer, K.; Aronov, P.; Hammock, B. Mass spectrometry-based metabolomics. Mass Spectrom. Rev. 2007, 26, 51–78. [Google Scholar] [CrossRef]
- Sawada, Y.; Akiyama, K.; Sakata, A.; Kuwahara, A.; Otsuki, H.; Sakurai, T.; Saito, K.; Hirai, M. Widely targeted metabolomics based on large-scale MS/MS data for elucidating metabolite accumulation patterns in plants. Plant Cell Physiol. 2009, 50, 37–47. [Google Scholar] [CrossRef]
- Chen, W.; Gong, L.; Guo, Z.; Wang, W.; Zhang, H.; Liu, X.; Yu, S.; Xiong, L.; Luo, J. A novel integrated method for large-scale detection, identification, and quantification of widely targeted metabolites: Application in the study of rice metabolomics. Mol. Plant. 2013, 6, 1769–1780. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Zhang, L.; Huang, X.; Wang, X.; Yang, R.; Mao, J.; Wang, X.; Wang, X.; Zhang, Q.; Li, P. Identification of nutritional components in black Sesame determined by widely targeted metabolomics and traditional Chinese medicines. Molecules 2018, 23, 1180. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Song, J. Analysis of widely targeted metabolites of the euhalophyte Suaeda salsa under saline conditions provides new insights into salt tolerance and nutritional value in halophytic species. BMC Plant Biol. 2019, 19, 388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, C.; Shi, J.; Quan, S.; Cui, B.; Kleessen, S.; Nikoloski, Z.; Tohge, T.; Alexander, D.; Guo, L.; Lin, H.; et al. Metabolic variation between japonica and indica rice cultivars as revealed by non-targeted metabolomics. Sci. Rep. 2014, 4, 5067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, C.; Tohge, T.; Chan, S.; Song, Y.; Rao, J.; Cui, B.; Lin, H.; Wang, L.; Fernie, A.; Zhang, D.; et al. Identification of conserved and diverse metabolic shifts during rice grain development. Sci. Rep. 2016, 6, 20942. [Google Scholar] [CrossRef]
- Calingacion, M.; Boualaphanh, C.; Daygon, V.; Anacleto, R.; Hamilton, R.; Biais, B.; Deborde, C.; Maucourt, M.; Moing, A.; Mumm, R.; et al. A genomics and multi-platform metabolomics approach to identify new traits of rice quality in traditional and improved varieties. Metabolomics 2012, 8, 771–783. [Google Scholar] [CrossRef]
- Mumm, R.; Hageman, J.; Calingacion, M.; de Vos, R.C.H.; Jonker, H.; Erban, A.; Kopka, J.; Hansen, T.; Laursen, K.; Schjoerring, J.; et al. Multi-platform metabolomics analyses of a broad collection of fragrant and non-fragrant rice varieties reveals the high complexity of grain quality characteristics. Metabolomics 2016, 12, 38. [Google Scholar] [CrossRef] [Green Version]
- Yan, N.; Du, Y.; Liu, X.; Chu, M.; Shi, J.; Zhang, H.; Liu, Y.; Zhang, Z. A comparative UHPLC-QqQ-MS-based metabolomics approach for evaluating Chinese and north American wild rice. Food Chem. 2019, 275, 618–627. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, A.; Du, C.; Song, Z.; Wang, W.; He, Y.; Cai, D. An efficient method of developing synthetic allopolyploid rice (Oryza spp.). Genet. Resour. Crop Evol. 2014, 61, 809–816. [Google Scholar] [CrossRef]
- Gu, Y.; Dai, X.; Li, J. Rice quality analysis of different ploidy rice. Zhengzhou Univ. 2015, 47, 80–85. [Google Scholar] [CrossRef]
- Liu, Y.; Luan, L.; Long, W.; Wang, X.; Kong, F.; He, T.; Tu, S. Genetic relationships between Wx gene and starch quality in autotetraploid and diploid. Chin. J. Rice Sci. 2007, 21, 143–149. [Google Scholar] [CrossRef] [Green Version]
- Xie, H.; Huang, Q.; Li, G.; Wang, X.; Ye, C.; Qin, G. Differential expression of the proteins in endosperms of rice with different chromosome sets. Hereditas 2007, 29, 360–364. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.; Zhao, Q.; Zhou, L.; Zhao, C.; Zhang, Y.; Wang, C. Research progress on the relationship between rice protein content and eating quality and the influence factors. Jiangsu J. Agric. Sci. 2020, 36, 1305–1311. [Google Scholar] [CrossRef]
- Friedman, M. Nutritional value of proteins from different food sources. A Review. J. Agric. Food Chem. 1996, 44, 6–29. [Google Scholar] [CrossRef]
- Ghosh, M. Review on recent trends in rice bran oil processing. J. Am. Oil Chem. Soc. 2007, 84, 315–324. [Google Scholar] [CrossRef]
- Velioglu, Y.; Mazza, G.; Gao, L.; Oomah, B. Antioxidant activity and total phenolics in selected fruits, vegetables, and grain products. J. Agric. Food Chem. 1998, 46, 4113–4117. [Google Scholar] [CrossRef]
- Cai, Y.; Luo, Q.; Sun, M.; Corke, H. Antioxidant activity and phenolic compounds of 112 traditional Chinese medicinal plants associated with anticancer. Life Sci. 2004, 74, 2157–2184. [Google Scholar] [CrossRef] [PubMed]
- Shen, P.; Luo, Q.; Jin, Z. Relationship between protein content and the cooking and eating quality properties of rice grain. J. Northeast Agric. Univ. 2003, 4, 378–381. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, F. Relation Between protein, lipids of rice and rice quality: A Review. J. Chin. Cereals Oils Assoc. 2006, 4, 6–10. [Google Scholar] [CrossRef]
- Mochizuki, T.; Hara, S. Usefulness of the low protein rice on the diet therapy in patients with chronic renal failure. Jpn. J. Nephrol. 2000, 42, 24–29. [Google Scholar] [CrossRef]
- Zhang, X.; Zhou, R.; Feng, L. Chang of rice lipids during storage and it’s effects on rice gelatinization. J. Chin. Cereals Oils Assoc. 1998, 13, 16–20. Available online: http://qikan.cqvip.com/Qikan/Article/Detail?id=3214213&from=Qikan_Search_Index (accessed on 1 September 2021).
- Liu, G.; Zhou, X.; Chen, D.; Guo, J.; Chen, P.; Chen, K.; Li, Y.; Chen, Y. Progress and prospect of functional rice research. Guangdong Agric. Sci. 2021, 48, 87–99. [Google Scholar] [CrossRef]
- Hu, S.; Hu, P. Research progress and prospect of functional rice. Chin. J. Rice Sci. 2021, 35, 311–325. [Google Scholar] [CrossRef]
- Shao, Y. Nutritional and functional characteristics of rice grain. China Rice 2020, 26, 1–11. [Google Scholar] [CrossRef]
- Nakamori, E. On the occurrence of the tetraploid plant of rice (Oryza sativa L.). Proc. Imp. Acad. 1993, 9, 340–341. [Google Scholar] [CrossRef] [Green Version]
- Koide, Y.; Kuniyoshi, D.; Kishima, Y. Fertile Tetraploids: New resources for future rice breeding? Front. Plant Sci. 2020, 11, 1231. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lipid Compounds | B-2x vs. B-4x | Y-2x vs. Y-4x | ||||
|---|---|---|---|---|---|---|
| VIP | p | Trend | VIP | p | Trend | |
| γ-Linolenic acid | 2.13 | 1.85 × 10−3 | up | 1.11 | 8.38 × 10−4 | up |
| Punicic acid | 2.22 | 4.29 × 10−3 | down | 1.55 | 4.28 × 10−4 | down |
| LysoPC (15:0) | 1.28 | 3.73 × 10−4 | up | 1.25 | 3.94 × 10−4 | up |
| LysoPC (16:1) | 1.67 | 8.71 × 10−4 | up | 1.76 | 3.73 × 10−4 | up |
| LysoPC (18:1) | 2.50 | 9.17 × 10−4 | up | 2.31 | 1.15 × 10−3 | up |
| LysoPC (18:3) | 5.62 | 4.16 × 10−5 | up | 2.82 | 1.97 × 10−3 | up |
| LysoPC (18:3) (2n isomer) | 5.48 | 2.69 × 10−4 | up | 3.02 | 2.54 × 10−3 | up |
| 4-Hydroxysphinganine | 1.07 | 6.91 × 10−4 | down | 1.82 | 1.74 × 10−3 | up |
| Choline alfoscerate | 1.34 | 6.37 × 10−3 | up | 1.39 | 7.18 × 10−4 | up |
| MAG (18:2) | 5.34 | 2.33 × 10−4 | up | 4.83 | 9.12 × 10−5 | up |
| MAG (18:3) isomer1 | 2.26 | 1.89 × 10−4 | up | 1.08 | 1.13 × 10−4 | up |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, W.; Tu, Q.; Chen, R.; Lv, P.; Xu, Y.; Xie, Q.; Song, Z.; He, Y.; Cai, D.; Zhang, X. Polyploidization Increases the Lipid Content and Improves the Nutritional Quality of Rice. Plants 2022, 11, 132. https://doi.org/10.3390/plants11010132
Wang W, Tu Q, Chen R, Lv P, Xu Y, Xie Q, Song Z, He Y, Cai D, Zhang X. Polyploidization Increases the Lipid Content and Improves the Nutritional Quality of Rice. Plants. 2022; 11(1):132. https://doi.org/10.3390/plants11010132
Chicago/Turabian StyleWang, Wei, Qiang Tu, Rongrong Chen, Pincang Lv, Yanqing Xu, Qian Xie, Zhaojian Song, Yuchi He, Detian Cai, and Xianhua Zhang. 2022. "Polyploidization Increases the Lipid Content and Improves the Nutritional Quality of Rice" Plants 11, no. 1: 132. https://doi.org/10.3390/plants11010132
APA StyleWang, W., Tu, Q., Chen, R., Lv, P., Xu, Y., Xie, Q., Song, Z., He, Y., Cai, D., & Zhang, X. (2022). Polyploidization Increases the Lipid Content and Improves the Nutritional Quality of Rice. Plants, 11(1), 132. https://doi.org/10.3390/plants11010132