
Physiological and Morphological Responses of Okra (Abelmoschus esculentus L.) to Rhizoglomus irregulare Inoculation under Ample Water and Drought Stress Conditions Are Cultivar Dependent

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Growth Conditions
2.2. Growth Substrate and AM Fungal Inoculation
2.3. Plant Material, Cultivation and Irrigation
2.4. Harvest, Quantification of AMF Colonized Root Length and Plant Growth Measurement
2.5. Nutrient Analysis and Statistics
3. Results
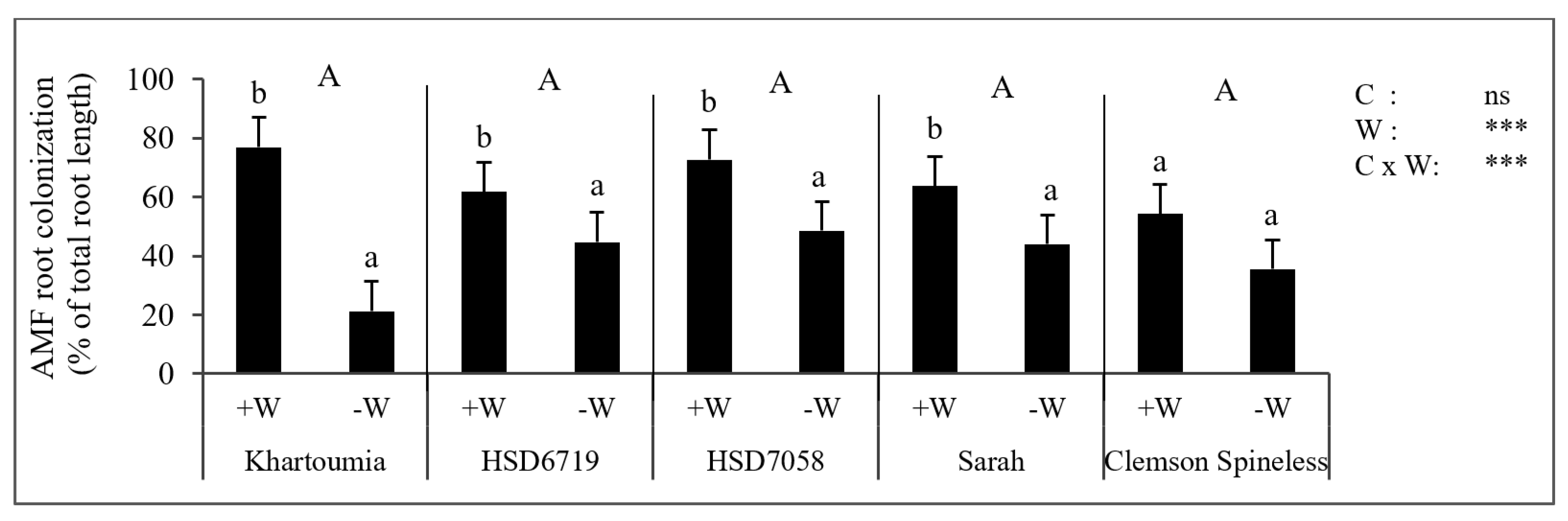
3.1. Mycorrhizal Status
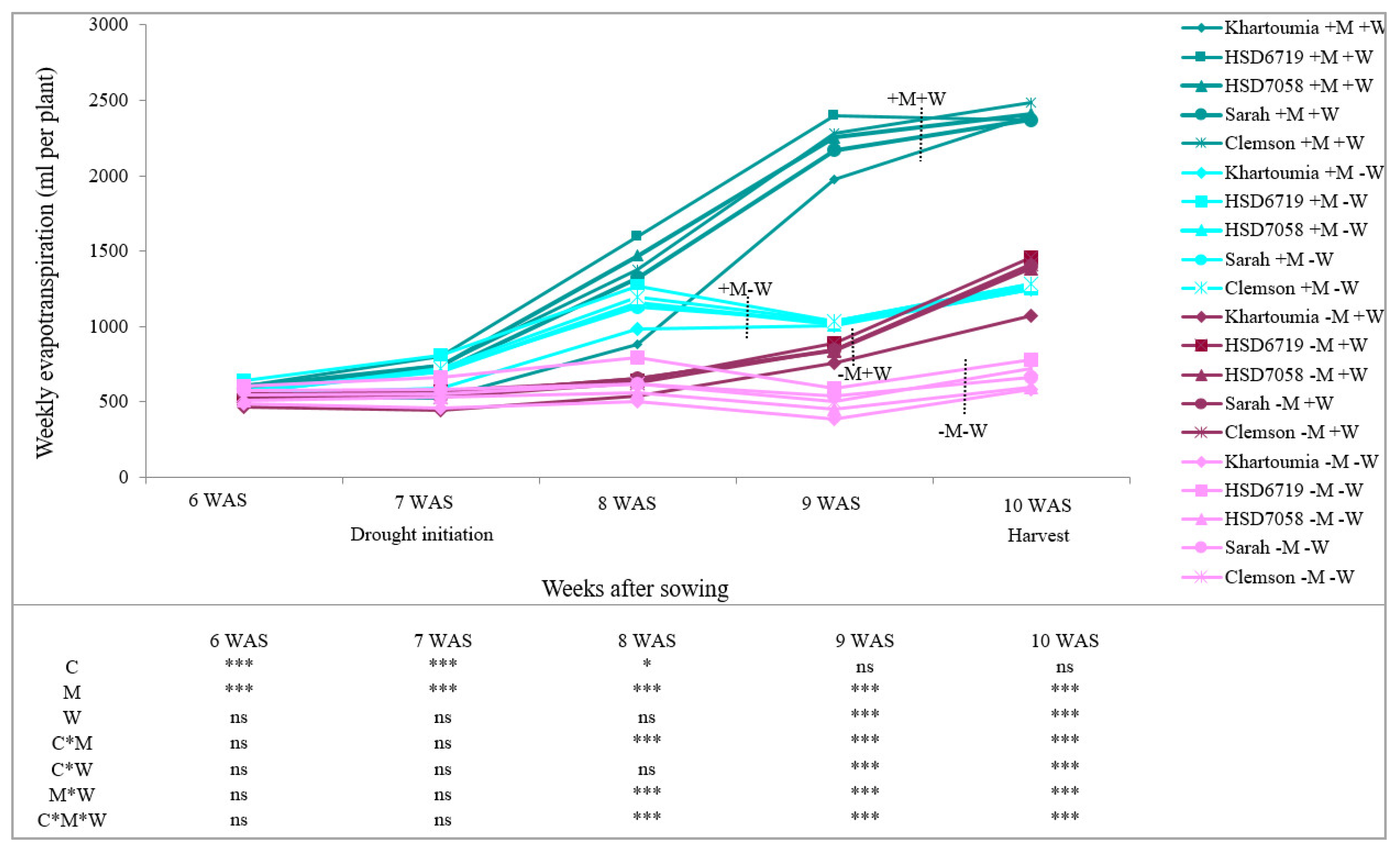
3.2. Evapotranspiration
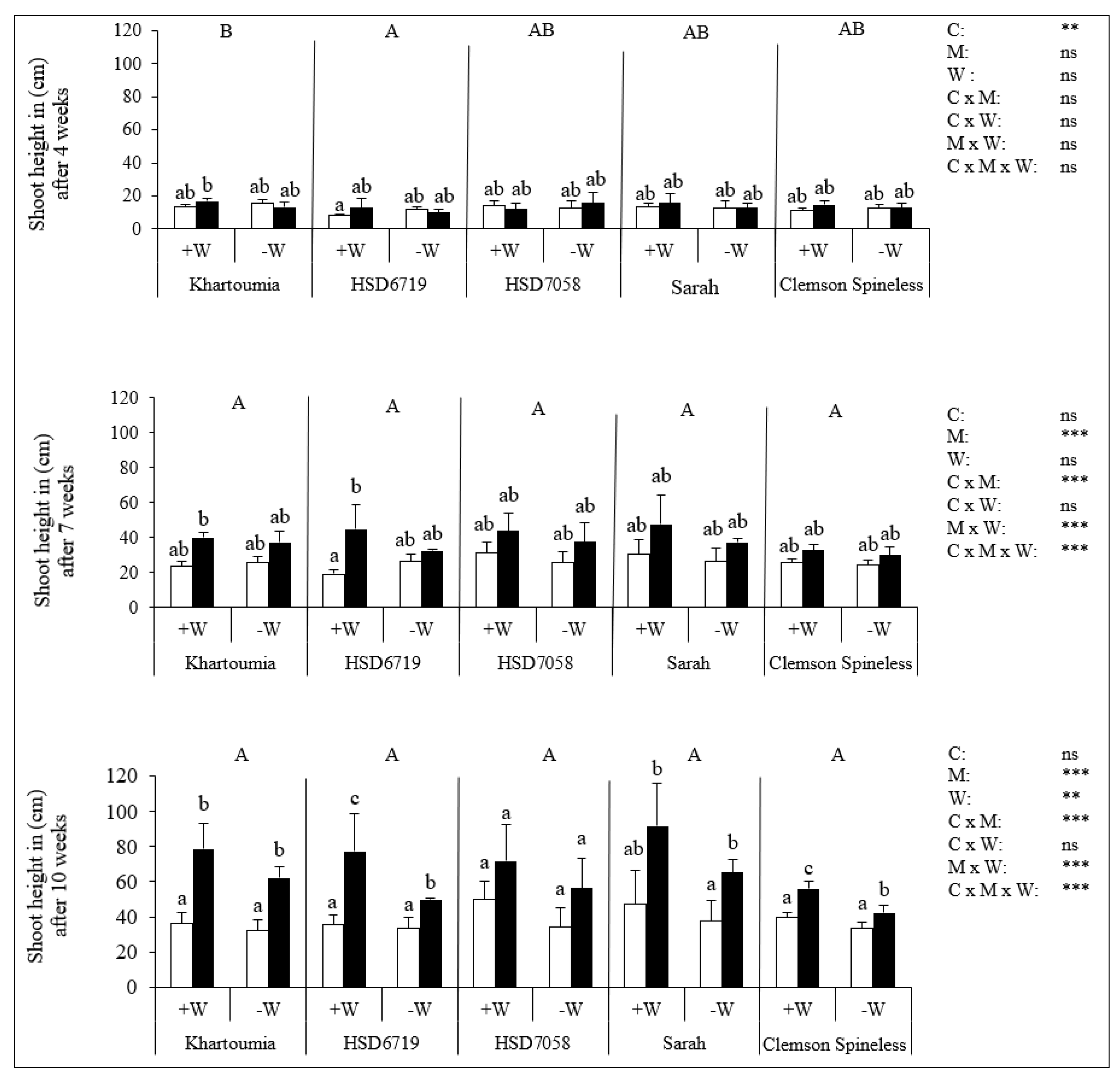
3.3. Plant Shoot Height
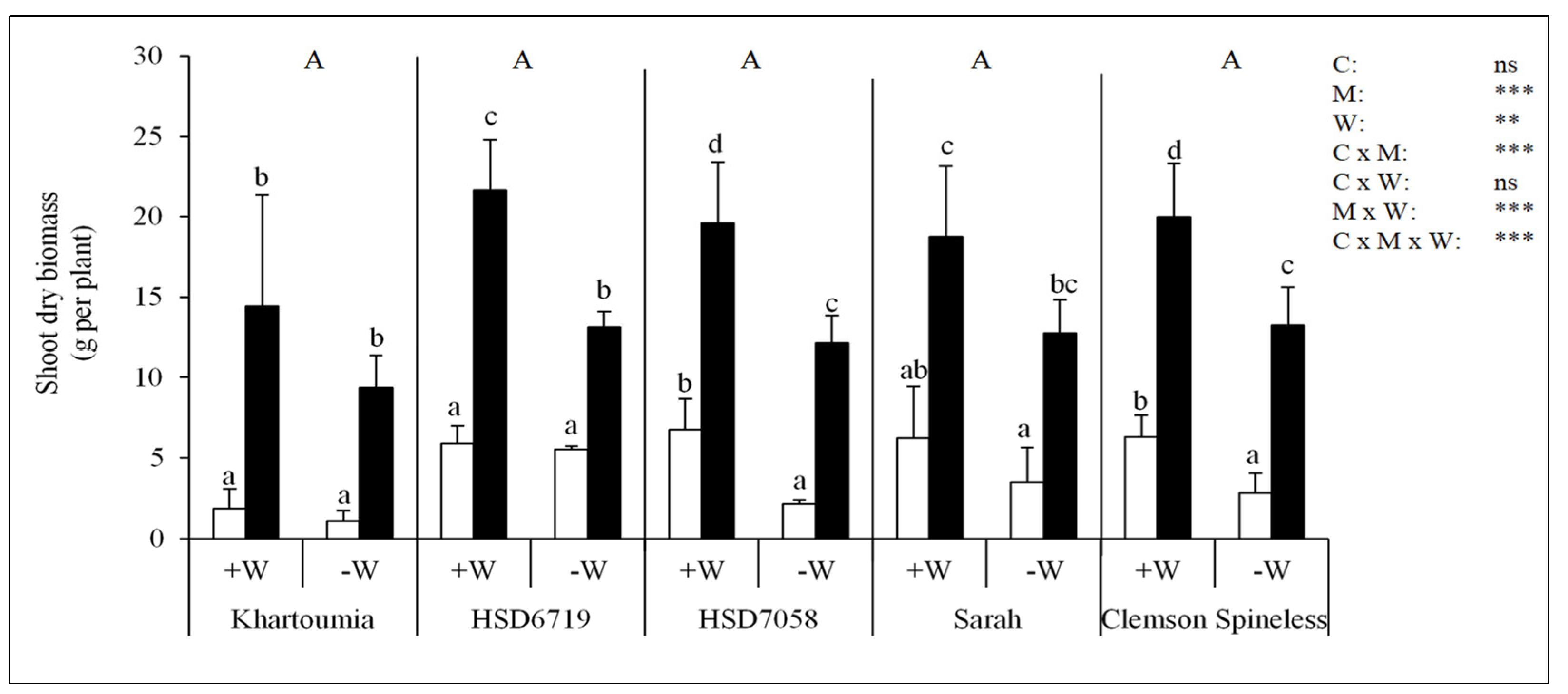
3.4. Shoot Dry Biomass
3.5. Leaf Area and Leaf-to-Stem Ratio
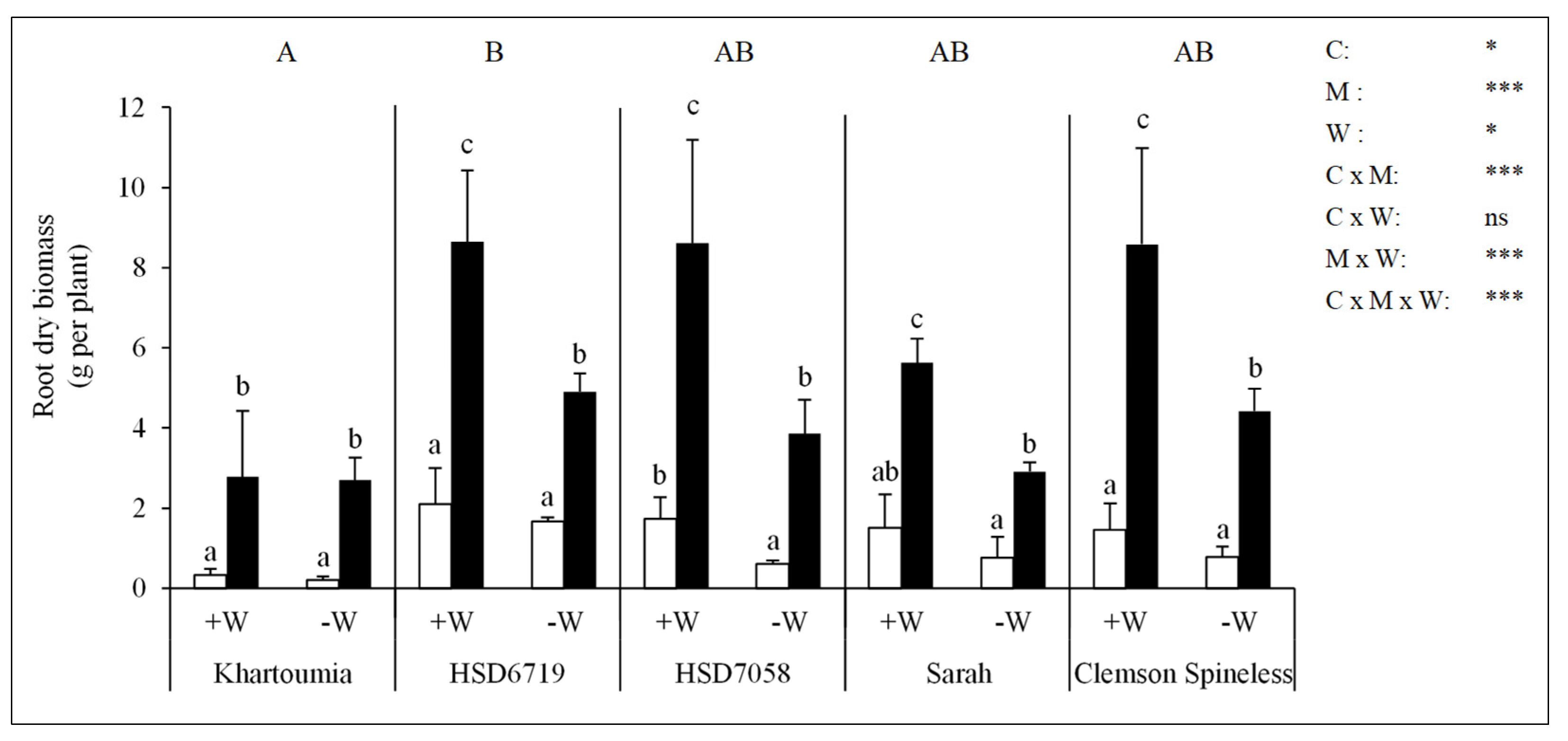
3.6. Root Dry Biomass
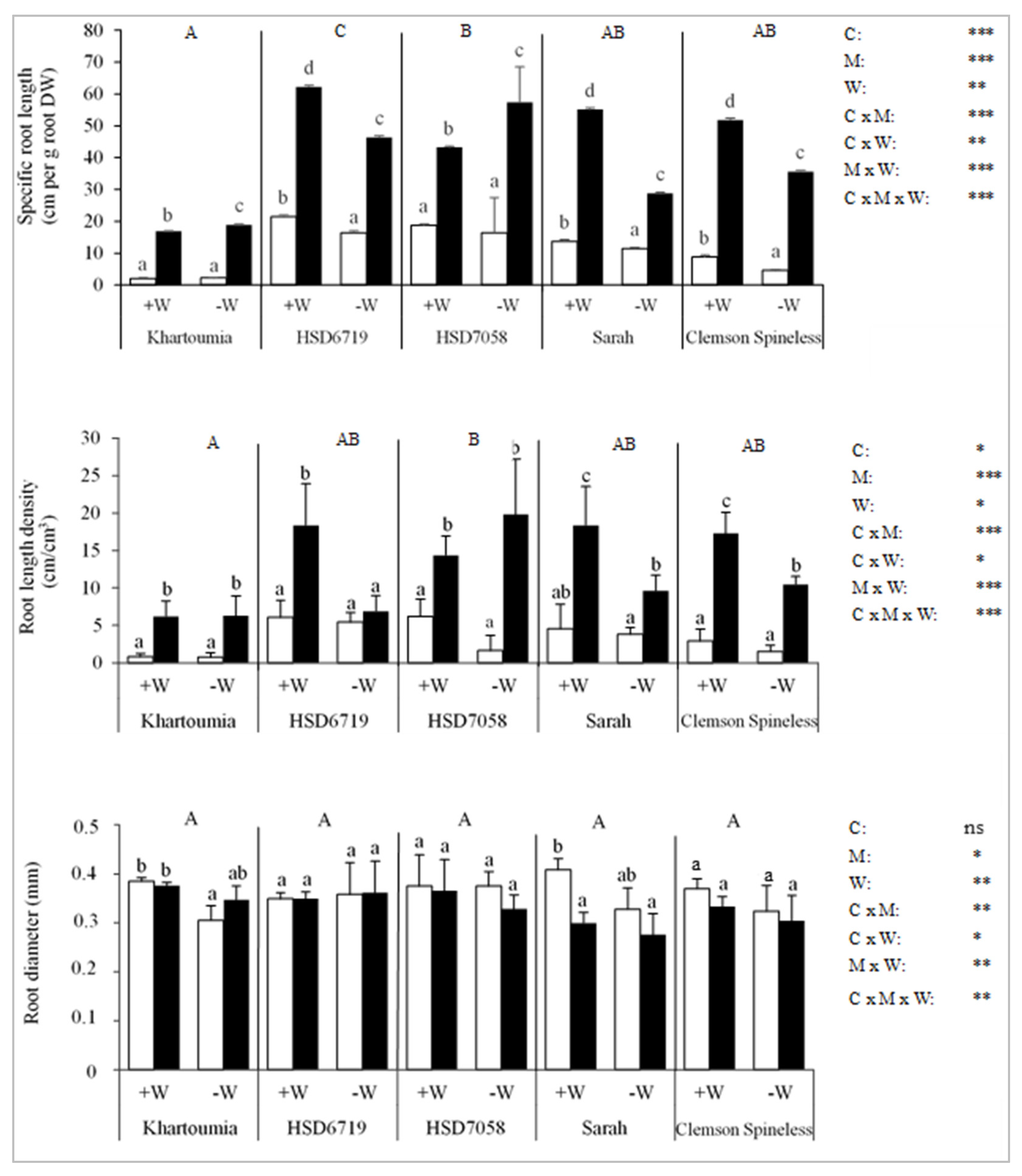
3.7. Root Development
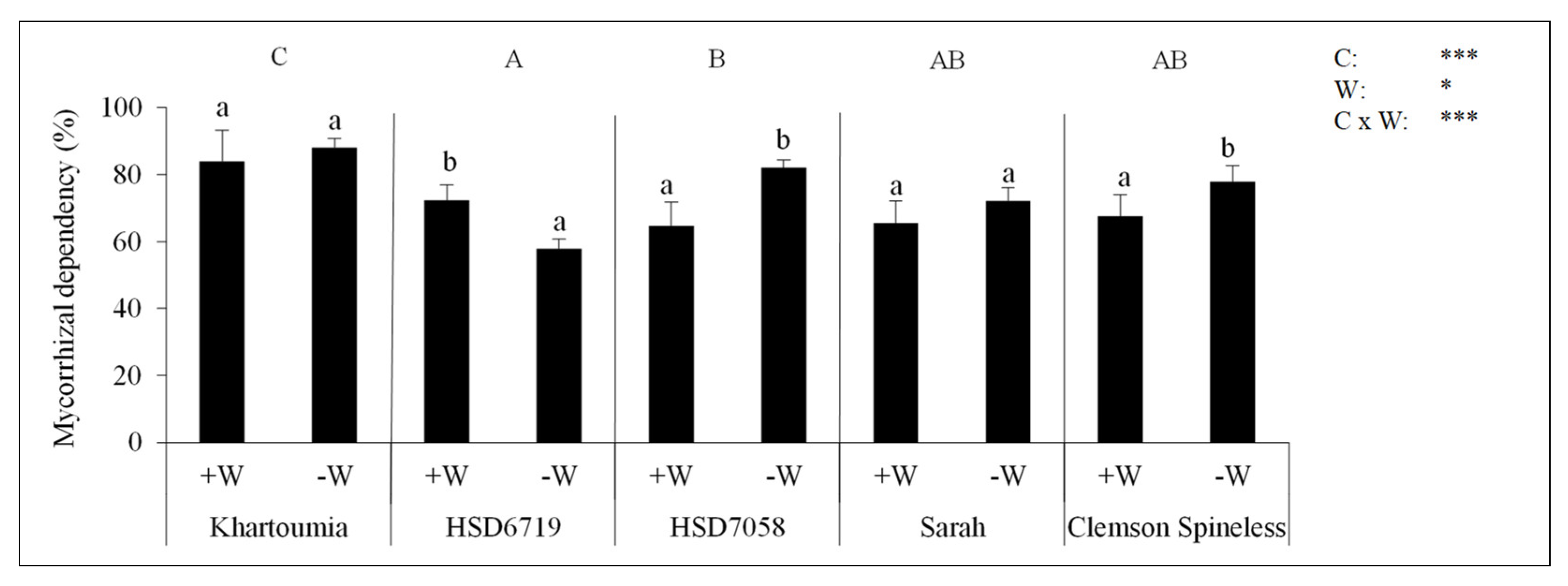
3.8. The Mycorrhizal Dependency (MD)
3.9. Plant Nutrient Status
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organic Matter Content (% of Substrate DW) | 1.8 |
| pH in (CaCl2) | 7.5 |
| Total cation exchange capacity (CEC) (µmol/g−1 DS) | 66 |
| CaCl2-extractable Mg (mg kg−1 DS) | 78 |
| CAL-extractable P (mg kg−1 DS) | 6.9 |
| CAL-extractable K (mg kg−1 DS) | 36 |
| Ca (mg kg−1 DS) | 3000 |
| CAT-extractable micronutrients: | |
| B (mg kg−1 DS) | 0.04 |
| Cu (mg kg−1 DS) | 0.5 |
| Mn (mg kg−1 DS) | 12 |
| Zn (mg kg−1 DS) | 0.2 |

| Khartoumia | HSD6719 | HSD7058 | Sarah | Clemson Spineless | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| +W | −W | +W | − W | +W | −W | +W | −W | +W | −W | ||||||||||
| −M | +M | −M | +M | −M | +M | −M | +M | −M | +M | −M | +M | −M | +M | −M | +M | −M | +M | −M | +M |
| Leaf area (cm2 per plant) | |||||||||||||||||||
| 35.8 a ± 10.74 | 88.1 b ± 33.97 | 19.4 a ± 8.66 | 82.1 b ± 9.89 | 81.4 a ± 25.91 | 160.4 b ± 15.41 | 72.1 a ± 12.19 | 130.9 b ± 13.47 | 109.5 b ± 13.43 | 177.0 c ± 29.29 | 38.50 a ± 4.50 | 118.5 b ± 31.27 | 69.2 a ± 23.07 | 111.8 b ± 14.79 | 46.6 a ± 26.59 | 101.0 b ± 14.40 | 93.2 ab ± 13.05 | 35.8 a ± 10.74 | 88.1 b ± 33.97 | 19.4 a ± 8.66 |
| A | C | C | B | C | |||||||||||||||
| Ratio leaf-to-stem | |||||||||||||||||||
| 1.11 a ± 0.19 | 0.69 a ± 0.22 | 0.88 a ± 0.25 | 0.94 a ± 0.13 | 1.91 b ± 0.15 | 0.84 a ± 0.24 | 1.02 a ± 0.27 | 1.14 a ± 0.42 | 1.15 a ± 0.46 | 1.43 a ± 0.14 | 0.71 a ± 0.75 | 0.85 a ± 0.24 | 1.31 b ± 0.65 | 0.78 ab ± 0.27 | 0.58 a ± 0.08 | 0.64 ab ± 0.08 | 1.40 b ± 0.37 | 1.11 a ± 0.19 | 0.69 a ± 0.22 | 0.88 a ± 0.25 |
| A | A | A | A | A | |||||||||||||||
| Leaf area | Ratio leaf-to-stem | ||||||||||||||||||
| C: | *** | ns | |||||||||||||||||
| M: | *** | ns | |||||||||||||||||
| W: | *** | ** | |||||||||||||||||
| C × M: | ns | ns | |||||||||||||||||
| C × W: | ** | * | |||||||||||||||||
| M × W: | ns | ** | |||||||||||||||||
| C × M × W: | ns | ** | |||||||||||||||||


References
- Lipiec, J.; Doussan, C.; Nosalewicz, A.; Kondracka, K. Effect of drought and heat stresses on plant growth and yield: A review. Int. Agrophys. 2013, 27, 463–477. [Google Scholar] [CrossRef]
- Sehgal, A.; Sita, K.; Siddique, K.H.; Kumar, R.; Bhogireddy, S.; Varshney, R.K.; Hanumantha Rao, B.; Nair, R.M.; Prasad, P.; Nayyar, H. Drought or/and heat-stress effects on seed filling in food crops: Impacts on functional biochemistry, seed yields, and nutritional quality. Front. Plant Sci. 2018, 9, 1705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubois, M.; Inzé, D. Plant growth under suboptimal water conditions: Early responses and methods to study them. J. Exp. Bot. 2020, 71, 1706–1722. [Google Scholar] [CrossRef]
- Rouphael, Y.; Cardarelli, M.; Schwarz, D.; Franken, P.; Colla, G. Effects of Drought on Nutrient Uptake and Assimilation in Vegetable Crops. In Plant Responses to Drought Stress; Aroca, R., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 171–195. [Google Scholar]
- Hu, Y.; Schmidhalter, U. Drought and salinity: A comparison of their effects on mineral nutrition of plants. J. Plant Nutr. Soil Sci. 2005, 168, 541–549. [Google Scholar] [CrossRef]
- He, M.; Dijkstra, F.A. Drought effect on plant nitrogen and phosphorus: A meta-analysis. New Phytol. 2014, 204, 924–931. [Google Scholar] [CrossRef]
- Liu, W.; Zhang, Y.; Jiang, S.; Deng, Y.; Christie, P.; Murray, P.J.; Li, X.; Zhang, J. Arbuscular mycorrhizal fungi in soil and roots respond differently to phosphorus inputs in an intensively managed calcareous agricultural soil. Sci. Rep. 2016, 6, 24902. [Google Scholar] [CrossRef] [Green Version]
- Picon-Cochard, C.; Pilon, R.; Tarroux, E.; Pagès, L.; Robertson, J.; Dawson, L. Effect of species, root branching order and season on the root traits of 13 perennial grass species. Plant Soil 2012, 353, 47–57. [Google Scholar] [CrossRef]
- Narayanan, S.; Mohan, A.; Gill, K.S.; Prasad, P.V. Variability of root traits in spring wheat germplasm. PLoS ONE 2014, 9, e100317. [Google Scholar] [CrossRef]
- Qiao, S.; Fang, Y.; Wu, A.; Xu, B.; Zhang, S.; Deng, X.; Djalovic, I.; Siddique, K.H.; Chen, Y. Dissecting root trait variability in maize genotypes using the semi-hydroponic phenotyping platform. Plant Soil 2019, 439, 75–90. [Google Scholar] [CrossRef]
- Fried, H.G.; Narayanan, S.; Fallen, B. Evaluation of soybean [Glycine max (L.) Merr.] genotypes for yield, water use efficiency, and root traits. PLoS ONE 2019, 14, e0212700. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.; Li, H.; Shen, Q.; Tang, X.; Xiong, C.; Li, H.; Pang, J.; Ryan, M.H.; Lambers, H.; Shen, J. Tradeoffs among root morphology, exudation and mycorrhizal symbioses for phosphorus-acquisition strategies of 16 crop species. New Phytol. 2019, 2, 882–895. [Google Scholar] [CrossRef]
- Fenta, B.; Beebe, S.; Kunert, K.; Burridge, J.; Barlow, K.; Lynch, J.; Foyer, C. Field phenotyping of soybean roots for drought stress tolerance. Agronomy 2014, 4, 418–435. [Google Scholar] [CrossRef] [Green Version]
- Rich, S.M.; Watt, M. Soil conditions and cereal root system architecture: Review and considerations for linking Darwin and Weaver. J. Exp. Bot. 2013, 64, 1193–1208. [Google Scholar] [CrossRef] [Green Version]
- Tawaraya, K. Arbuscular mycorrhizal dependency of different plant species and cultivars. Soil Sci. Plant Nutr. 2003, 49, 655–668. [Google Scholar] [CrossRef]
- Bowles, T.M.; Jackson, L.E.; Cavagnaro, T.R. Mycorrhizal fungi enhance plant nutrient acquisition and modulate nitrogen loss with variable water regimes. Glob. Chang. Biol. 2018, 24, 171–182. [Google Scholar] [CrossRef]
- Khalil, S.; Loynachan, T.E.; Tabatabai, M.A. Mycorrhizal dependency and nutrient uptake by improved and unimproved corn and soybean cultivars. Agron. J. 1994, 86, 949–958. [Google Scholar] [CrossRef]
- Gao, X.; Kuyper, T.W.; Zou, C.; Zhang, F.; Hoffland, E. Mycorrhizal responsiveness of aerobic rice genotypes is negatively correlated with their zinc uptake when nonmycorrhizal. Plant Soil 2007, 290, 283–291. [Google Scholar] [CrossRef]
- Chu, Q.; Wang, X.; Yang, Y.; Chen, F.; Zhang, F.; Feng, G. Mycorrhizal responsiveness of maize (Zea mays L.) genotypes as related to releasing date and available P content in soil. Mycorrhiza 2013, 23, 497–505. [Google Scholar] [CrossRef]
- Salloum, M.S.; Guzzo, M.C.; Velazquez, M.S.; Sagadin, M.B.; Luna, C.M. Variability in colonization of arbuscular mycorrhizal fungi and its effect on mycorrhizal dependency of improved and unimproved soybean cultivars. Can. J. Microbiol. 2016, 62, 1034–1040. [Google Scholar] [CrossRef]
- Bazghaleh, N.; Hamel, C.; Gan, Y.; Tar’an, B.; Knight, J.D. Genotypic variation in the response of chickpea to arbuscular mycorrhizal fungi and non-mycorrhizal fungal endophytes. Can. J. Microbiol. 2018, 64, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Oruru, M.B.; Njeru, E.M.; Pasquet, R.; Runo, S. Response of a wild-type and modern cowpea cultivars to arbuscular mycorrhizal inoculation in sterilized and non-sterilized soil. J. Plant Nutr. 2018, 41, 90–101. [Google Scholar] [CrossRef]
- Watts-Williams, S.J.; Emmett, B.D.; Levesque-Tremblay, V. Diverse Sorghum bicolor accessions show marked variation in growth and transcriptional responses to arbuscular mycorrhizal fungi. Plant Cell Environ. 2019, 42, 1758–1774. [Google Scholar] [CrossRef] [PubMed]
- Watts-Williams, S.J.; Cavagnaro, T.R.; Tyerman, S.D. Variable effects of arbuscular mycorrhizal fungal inoculation on physiological and molecular measures of root and stomatal conductance of diverse Medicago truncatula accessions. Plant Cell Environ. 2019, 42, 285–294. [Google Scholar] [CrossRef] [Green Version]
- Dreher, D.; Yadav, H.; Zander, S.; Hause, B. Is there genetic variation in mycorrhization of Medicago truncatula? PeerJ 2017, 5, e3713. [Google Scholar] [CrossRef] [Green Version]
- Hetrick, B.; Wilson, G.; Cox, T. Mycorrhizal dependence of modern wheat cultivars and ancestors: A synthesis. Can. J. Bot. 1993, 71, 512–518. [Google Scholar] [CrossRef]
- Abdelhalim, T.; Jannoura, R.; Joergensen, R.G. Arbuscular mycorrhizal dependency and phosphorus responsiveness of released, landrace and wild Sudanese sorghum genotypes. Arch. Agron. Soil Sci. 2020, 66, 706–716. [Google Scholar] [CrossRef]
- Rillig, M.C.; Aguilar-Trigueros, C.A.; Camenzind, T.; Cavagnaro, T.R.; Degrune, F.; Hohmann, P.; Lammel, D.R.; Roy, J.; van der Heijden, M.G.; Yang, G. Why farmers should manage the arbuscular mycorrhizal symbiosis. New Phytol. 2019, 222, 1171–1175. [Google Scholar] [CrossRef] [Green Version]
- Geneif, A.A. Tapping natural genetic variability of okra in the Sudan. Acta Hortic. 1984, 143, 175–182. [Google Scholar] [CrossRef]
- Koske, R.E.; Gemma, J.N. A modified procedure for staining roots to detect VA Mycorrhizas. Mycol. Res. 1989, 92, 486–488. [Google Scholar] [CrossRef]
- Tennant, D. A test of a modified line intersect method of estimating root length. J. Ecol. 1975, 63, 995–1001. [Google Scholar] [CrossRef]
- Kormanik, P.P.; McGraw, A.C. Quantification of Vesicular-Arbuscular Mycorrhizae in Plant Roots. In Methods and Principles of Mycorrhizal Research; Schenck, N.C., Ed.; The American Phytopathological Society: St. Paul, MN, USA, 1982; pp. 37–45. [Google Scholar]
- Chen, R.; Huang, J.W.; Chen, Z.K.; Xu, Y.; Liu, J.; Ge, Y.H. Effect of root density of wheat and okra on hydraulic properties of an unsaturated compacted loam. Eur. J. Soil Sci. 2019, 70, 493–506. [Google Scholar] [CrossRef]
- Abubaker, B.M.A.; Alhadi, M.; Shuang’en, Y.; Guangcheng, S. Different Irrigation Methods for Okra Crop Production under Semi-arid Conditions. Int. J. Eng. Res. Technol. 2014, 3, 787–794. [Google Scholar]
- Maduwanthi, A.K.M.R.B.; Karunarathna, B. Growth and Dry Matter Accumulation of Okra (Abelmoschus esculentus L.) as Influenced by Different Plating Pattern Under Okra—Cowpea (Vigna unguiculata L.) Intercropping. J. Hortic. Plant Res. 2019, 7, 81–96. [Google Scholar] [CrossRef] [Green Version]
- Nwangburuka, C.C.; Olawuyi, O.J.; Oyekale, O.; Ogunwwenmo, K.O.; Denton, O.; Daramola, D.S.; Awotade, A. Effect of Arburscular mycorrhizea [AM], poultry manure [PM], NPK fertilizer and the combination of AM-PM on the growth and yield of okra [Abelmoschus esculentus]. Int. J. Org. Agric. Res. Dev. 2010, 7, 27–42. [Google Scholar]
- Kusvuran, S. Influence of drought stress on growth, ion accumulation and antioxidative enzymes in okra genotypes. Int. J. Agric. Biol. 2012, 14, 401–406. [Google Scholar]
- Abbas, T.; Pervez, M.A.; Ayyub, C.M.; Ahmad, R. Assessment of morphological, antioxidant, biochemical and ionic responses of salt-tolerant and salt-sensitive okra (Abelmoschus esculentus) under saline regime. Pak. J. Life Soc. Sci. 2013, 11, 147–153. [Google Scholar]
- Kumar, A.; Kumar, M.; Sharma, V.R.; Singh, M.K.; Singh, B.; Chand, P. Genetic Variability, Heritability and Genetic Advance studies in Genotypes of Okra [(Abelmoschus esculentus (L.) Moench]. J. Pharmacogn. Phytochem. 2019, 8, 1285–1290. [Google Scholar]
- Hochmuth, G.; Maynard, D.; Vavrina, C.; Hanlon, E. Plant Tissue Analysis and Interpretations for Vegetable Crops in Florida; Ext. Bul. SS-VEC-42; University of Florida: Gainesville, FL, USA, 1991. [Google Scholar]
- Marschner, H.; Dell, B. Nutrient uptake in mycorrhizal symbiosis. Plant Soil 1994, 159, 89–102. [Google Scholar] [CrossRef]
- George, E. Nutrient Uptake. In Arbuscular Mycorrhizas: Physiology and Function; Kapulnik, Y., Douds, D.D., Eds.; Springer: Dordrecht, The Netherlands, 2000; pp. 307–343. [Google Scholar]
- Neumann, E.; George, E. Colonisation with the arbuscular mycorrhizal fungus Glomus mosseae (Nicol. & Gerd.) enhanced phosphorus uptake from dry soil in Sorghum bicolor (L.). Plant Soil 2004, 261, 245–255. [Google Scholar]
- Bowles, T.M.; Barrios-Masias, F.H.; Carlisle, E.A.; Cavagnaro, T.R.; Jackson, L.E. Effects of arbuscular mycorrhizae on tomato yield, nutrient uptake, water relations, and soil carbon dynamics under deficit irrigation in field conditions. Sci. Total Environ. 2016, 566, 1223–1234. [Google Scholar] [CrossRef] [Green Version]
- Asrar, A.A.; Abdel-Fattah, G.M.; Elhindi, K.M. Improving growth, flower yield, and water relations of snapdragon (Antirhinum majus L.) plants grown under well-watered and water-stress conditions using arbuscular mycorrhizal fungi. Photosynthetica 2012, 50, 305–316. [Google Scholar] [CrossRef]
- Ravnskov, S.; Jakobsen, I. Functional compatibility in arbuscular mycorrhizas measured as hyphal P transport to the plant. New Phytol. 1995, 129, 611–618. [Google Scholar] [CrossRef]
- UMA, T.; Aswim, R.; Kale, R.D.; Bagyaraj, D. Symbiotic response of Abelmoschus esculentus (Okra) to different arbuscular mycorrhizal fungi. J. Soil Biol. Ecol. 2017, 37, 19–25. [Google Scholar]
- Püschel, D.; Janoušková, M.; Voříšková, A.; Gryndlerová, H.; Vosátka, M.; Jansa, J. Arbuscular mycorrhiza stimulates biological nitrogen fixation in two Medicago spp. through improved phosphorus acquisition. Front. Plant Sci. 2017, 27, 390. [Google Scholar] [CrossRef] [Green Version]
- Beltrano, J.; Ruscitti, M.; Arango, M.C.; Ronco, M. Effects of arbuscular mycorrhiza inoculation on plant growth, biological and physiological parameters and mineral nutrition in pepper grown under different salinity and p levels. J. Soil Sci. Plant Nutr. 2013, 13, 123–141. [Google Scholar] [CrossRef] [Green Version]
- Symanczik, S.; Lehmann, M.F.; Wiemken, A. Effects of two contrasted arbuscular mycorrhizal fungal isolates on nutrient uptake by Sorghum bicolor under drought. Mycorrhiza 2018, 28, 779–785. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: London, UK, 2008. [Google Scholar]
- Rapparini, F.; Peñuelas, J. Mycorrhizal Fungi to Alleviate Drought Stress on Plant Growth. In Use of Microbes for the Alleviation of Soil Stresses; Miransari, M., Ed.; Springer: New York, NY, USA, 2014; Volume 1, pp. 21–42. [Google Scholar]
- Bryla, D.; Duniway, J. Growth, phosphorus uptake, and water relations of safflower and wheat infected with an arbuscular mycorrhizal fungus. New Phytol. 1997, 136, 581–590. [Google Scholar] [CrossRef] [PubMed]
- Bitterlich, M.; Sandmann, M.; Graefe, J. Arbuscular mycorrhiza alleviates restrictions to substrate water flow and delays transpiration limitation to stronger drought in tomato. Front. Plant Sci. 2018, 9, 154. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Sheng, M.; Wang, C.; Chen, H.; Li, Z.; Tang, M. Impact of arbuscular mycorrhizal fungi on the growth, water status, and photosynthesis of hybrid poplar under drought stress and recovery. Photosynthetica 2015, 53, 250–258. [Google Scholar] [CrossRef]
- Moradtalab, N.; Hajiboland, R.; Aliasgharzad, N.; Hartmann, T.E.; Neumann, G. Silicon and the association with an arbuscular-mycorrhizal fungus (Rhizophagus clarus) mitigate the adverse effects of drought stress on strawberry. Agronomy 2019, 9, 41. [Google Scholar] [CrossRef] [Green Version]
- Augé, R.M. Do VA mycorrhizae enhance transpiration by affecting host phosphorus content? J. Plant Nutr. 1989, 12, 743–753. [Google Scholar] [CrossRef]
- Collier, S.C.; Yarnes, C.T.; Herman, R.P. Mycorrhizal dependency of Chihuahuan Desert plants is influenced by life history strategy and root morphology. J. Arid Environ. 2003, 55, 223–229. [Google Scholar] [CrossRef]
- Marschner, H. Mineral Nutrition of Higher Plants, 2nd ed.; Institute of Plant Nutrition, University of Hohenheim: Hoehenheim, Germany, 1995. [Google Scholar]
- Sawers, R.J.; Ramírez-Flores, M.R.; Olalde-Portugal, V.; Paszkowski, U. The impact of domestication and crop improvement on arbuscular mycorrhizal symbiosis in cereals: Insights from genetics and genomics. New Phytol. 2018, 220, 1135–1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schüller, H. Die CAL-Methode, eine neue Methode zur Bestimmung des pflanzenverfügbaren Phosphates in Böden. J. Plant Nutr. Soil Sci. 1969, 123, 48–63. [Google Scholar] [CrossRef]
- Alt, D.; Peters, I. Analysis of macro- and trace elements in horticultural substrates by means of the CaCl2/DTPA method. Acta Hortic. 1993, 49, 287–292. [Google Scholar] [CrossRef]







| Khartoumia | HSD6719 | HSD7058 | Sarah | Clemson Spineless | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| +W | −W | +W | −W | +W | −W | +W | −W | +W | −W | ||||||||||
| −M | +M | −M | +M | −M | +M | −M | +M | −M | +M | −M | +M | −M | +M | −M | +M | −M | +M | −M | +M |
| Ratio root–shoot | |||||||||||||||||||
| 0.19 a ± 0.04 | 0.19 a ± 0.06 | 0.20 ab ± 0.04 | 0.29 b ± 0.03 | 0.34 a ± 0.11 | 0.39 a ± 0.04 | 0.30 a ± 0.01 | 0.37 a ± 0.03 | 0.27 a ± 0.10 | 0.43 b ± 0.08 | 0.28 ab ± 0.01 | 0.32 ab ± 0.09 | 0.24 ab ± 0.04 | 0.31 b ± 0.07 | 0.21 a ± 0.03 | 0.23 ab ± 0.03 | 0.22 a ± 0.06 | 0.43 b ± 0.09 | 0.29 ab ± 0.10 | 0.35 ab ± 0.12 |
| A | C | BC | AB | BC | |||||||||||||||
| C: | *** | ||||||||||||||||||
| M: | *** | ||||||||||||||||||
| W: | ns | ||||||||||||||||||
| C × M: | ns | ||||||||||||||||||
| C × W: | ns | ||||||||||||||||||
| M × W: | ns | ||||||||||||||||||
| C × M × W: | ns | ||||||||||||||||||
| Khartoumia | HSD6719 | HSD7058 | Sarah | Clemson Spineless | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| +W | −W | +W | − W | +W | −W | +W | −W | +W | −W | ||||||||||
| −M | +M | −M | +M | −M | +M | −M | +M | −M | +M | −M | +M | −M | +M | −M | +M | −M | +M | −M | +M |
| P concentration (mg g−1 DW) | |||||||||||||||||||
| 0.90 ab ± 0.17 | 1.43 b ± 0.42 | 0.65 a ± 0.06 | 1.03 ab ± 0.15 | 0.90 b ± 0.08 | 1.65 d ± 0.21 | 0.60 a ± 0.08 | 1.20 c ± 0.08 | 0.98 ab ± 0.05 | 1.75 c ± 0.19 | 0.63 a ± 0.06 | 1.08 b ± 0.21 | 0.88 ab ± 0.05 | 1.65 c ± 0.25 | 0.60 a ± 0.12 | 0.93 b ± 0.10 | 0.85 ab ± 0.06 | 1.85 b ± 0.19 | 0.60 a ± 0.20 | 1.20 ab ± 0.00 |
| A | A | A | A | A | |||||||||||||||
| N concentration (mg g−1 DW) | |||||||||||||||||||
| 33.15 bc ± 2.47 | 17.83 a ± 6.31 | 42.97 c ± 3.08 | 25.08 ab ± 4.28 | 33.88 b ± 4.51 | 12.89 a ± 1.39 | 34.64 b ± 2.53 | 17.52 a ± 1.02 | 30.46 c ± 1.60 | 13.64 a ± 1.78 | 34.20 c ± 2.30 | 18.64 b ± 1.74 | 27.38 b ± 4.56 | 13.74 a ± 2.65 | 33.13 b ± 3.43 | 18.60 a ± 2.03 | 30.45 b ± 3.50 | 12.96 a ± 0.87 | 27.27 ab ± 10.19 | 17.38 ab ± 1.74 |
| A | A | A | A | A | |||||||||||||||
| Fe concentration (mg kg−1 DW) | |||||||||||||||||||
| 84.79 ab ± 22.71 | 48.12 a ± 15.14 | 124.06 b ± 81.19 | 58.18 ab ± 14.29 | 125.10 b ± 53.20 | 56.18 a ± 9.68 | 74.66 ab ± 11.59 | 74.57 ab ± 21.48 | 102.57 b ± 14.28 | 58.51 a ± 24.93 | 76.80 ab ± 26.81 | 43.18 a ± 2.90 | 92.36 ab ± 31.19 | 46.35 a ± 15.38 | 128.01 b ± 107.00 | 65.28 ab ± 20.15 | 148.25 b ± 107.40 | 53.87 a ± 13.18 | 75.24 ab ± 41.34 | 61.59 ab ± 20.43 |
| A | A | A | A | A | |||||||||||||||
| Zn concentration (mg kg−1 DW) | |||||||||||||||||||
| 37.34 a ± 4.58 | 38.43 a ± 6.55 | 43.50 a ± 13.82 | 39.42 a ± 3.56 | 47.44 a ± 3.43 | 44.44 a ± 6.02 | 38.29 a ± 6.38 | 39.42 a ± 2.62 | 48.42 b ± 4.44 | 47.06 ab ± 6.81 | 43.77 ab ± 6.43 | 35.96 a ± 5.28 | 42.98 a ± 3.27 | 43.71 a ± 6.70 | 40.00 a ± 5.57 | 35.20 a ± 7.37 | 47.43 bc ± 6.66 | 52.66 c ± 2.96 | 35.87 a ± 3.77 | 39.49 ab ± 4.06 |
| A | A | A | A | A | |||||||||||||||
| Khartoumia | HSD6719 | HSD7058 | Sarah | Clemson Spineless | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| +W | −W | +W | − W | +W | −W | +W | −W | +W | −W | ||||||||||
| −M | +M | −M | +M | −M | +M | −M | +M | −M | +M | −M | +M | −M | +M | −M | +M | −M | +M | −M | +M |
| P total content (mg per plant) | |||||||||||||||||||
| 1.60 a ± 0.88 | 22.20 b ± 13.40 | 0.71 a ± 0.45 | 9.46 b ± 1.55 | 5.23 b ± 0.66 | 35.28 d ± 3.69 | 3.30 a ± 0.35 | 15.69 c ± 0.57 | 6.50 b ± 1.52 | 33.80 d ± 3.21 | 1.37 a ± 0.27 | 13.10 c ± 3.21 | 5.44 ab ± 2.82 | 30.82 c ± 7.77 | 2.22 a ± 1.66 | 11.86 b ± 2.79 | 5.33 b ± 1.00 | 36.79 d ± 6.46 | 1.58 a ± 0.46 | 15.88 c ± 2.84 |
| A | A | A | A | A | |||||||||||||||
| N total content (mg per plant) | |||||||||||||||||||
| 62.24 a ± 42.15 | 242.17 b ± 96.82 | 45.11 a ± 24.29 | 229.06 b ± 16.92 | 197.36 a ± 37.63 | 275.42 b ± 15.11 | 190.96 a ± 14.29 | 229.36 ab ± 12.12 | 203.46 b ± 48.74 | 263.11 b ± 27.32 | 73.73 a ± 12.78 | 224.34 b ± 12.24 | 162.53 ab ± 80.93 | 251.67 b ± 38.23 | 112.55 a ± 60.59 | 234.15 b ± 12.61 | 188.71 b ± 22.12 | 257.05 b ± 30.82 | 82.65 b ± 49.72 | 28.47 a ± 37.48 |
| A | A | A | A | A | |||||||||||||||
| Fe total content (mg per plant) | |||||||||||||||||||
| 175.6 ab ± 131.5 | 635.7 c ± 251.0 | 1074 a ± 41.09 | 532.2 bc ± 127.5 | 753.3 ab ± 392.34 | 1200 b ± 183.33 | 410.3 a ± 53.08 | 983.7 b ± 311.9 | 686.8 ab ± 185.70 | 1116.7 b ± 394.22 | 157.0 a ± 61.97 | 523.3 a ± 68.45 | 623.1 a ± 403.31 | 859.4 a ± 288.99 | 336.0 a ± 141.5 | 814.4 a ± 201.7 | 909.3 ab ± 594.44 | 1084.9 b ± 357.02 | 236.1 a ± 208.41 | 812.8 ab ± 307.23 |
| A | A | A | A | A | |||||||||||||||
| Zn total content (mg per plant) | |||||||||||||||||||
| 87.1 ab ± 67.00 | 676.2 b ± 338.02 | 51.1 a ± 26.81 | 470.5 ab ± 72.76 | 373.7 a ± 72.62 | 1324.3 c ± 105.03 | 126.8 a ± 37.99 | 710.3 b ± 62.81 | 415.2 b ± 131.49 | 1296.8 c ± 98.53 | 273.6 a ± 31.87 | 577.1 b ± 118.5 | 335.0 ab ± 177.48 | 1059.0 c ± 193.67 | 174.0 a ± 107.9 | 544.0 b ± 81.0 | 359.0 a ± 46.14 | 1502.0 c ± 271.26 | 132.9 a ± 64.42 | 698.1 b 114.34 |
| A | A | A | A | A | |||||||||||||||
| P Concentration | N Concentration | Fe Concentration | Zn Concentration | P Content | N Content | Fe Content | Zn Content | |
|---|---|---|---|---|---|---|---|---|
| C: | ns | ns | ns | ns | ns | ns | ns | ns |
| M: | *** | *** | *** | ns | *** | *** | *** | *** |
| W: | *** | * | ns | *** | *** | ** | ** | *** |
| C × M: | *** | *** | *** | ns | *** | *** | *** | *** |
| C × W: | * | ns | ns | ** | ns | ns | ** | ns |
| M × W: | *** | *** | *** | ns | *** | *** | *** | *** |
| C × M × W: | *** | *** | *** | ns | *** | *** | ns | *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eltigani, A.; Müller, A.; Ngwene, B.; George, E. Physiological and Morphological Responses of Okra (Abelmoschus esculentus L.) to Rhizoglomus irregulare Inoculation under Ample Water and Drought Stress Conditions Are Cultivar Dependent. Plants 2022, 11, 89. https://doi.org/10.3390/plants11010089
Eltigani A, Müller A, Ngwene B, George E. Physiological and Morphological Responses of Okra (Abelmoschus esculentus L.) to Rhizoglomus irregulare Inoculation under Ample Water and Drought Stress Conditions Are Cultivar Dependent. Plants. 2022; 11(1):89. https://doi.org/10.3390/plants11010089
Chicago/Turabian StyleEltigani, Amna, Anja Müller, Benard Ngwene, and Eckhard George. 2022. "Physiological and Morphological Responses of Okra (Abelmoschus esculentus L.) to Rhizoglomus irregulare Inoculation under Ample Water and Drought Stress Conditions Are Cultivar Dependent" Plants 11, no. 1: 89. https://doi.org/10.3390/plants11010089
APA StyleEltigani, A., Müller, A., Ngwene, B., & George, E. (2022). Physiological and Morphological Responses of Okra (Abelmoschus esculentus L.) to Rhizoglomus irregulare Inoculation under Ample Water and Drought Stress Conditions Are Cultivar Dependent. Plants, 11(1), 89. https://doi.org/10.3390/plants11010089