
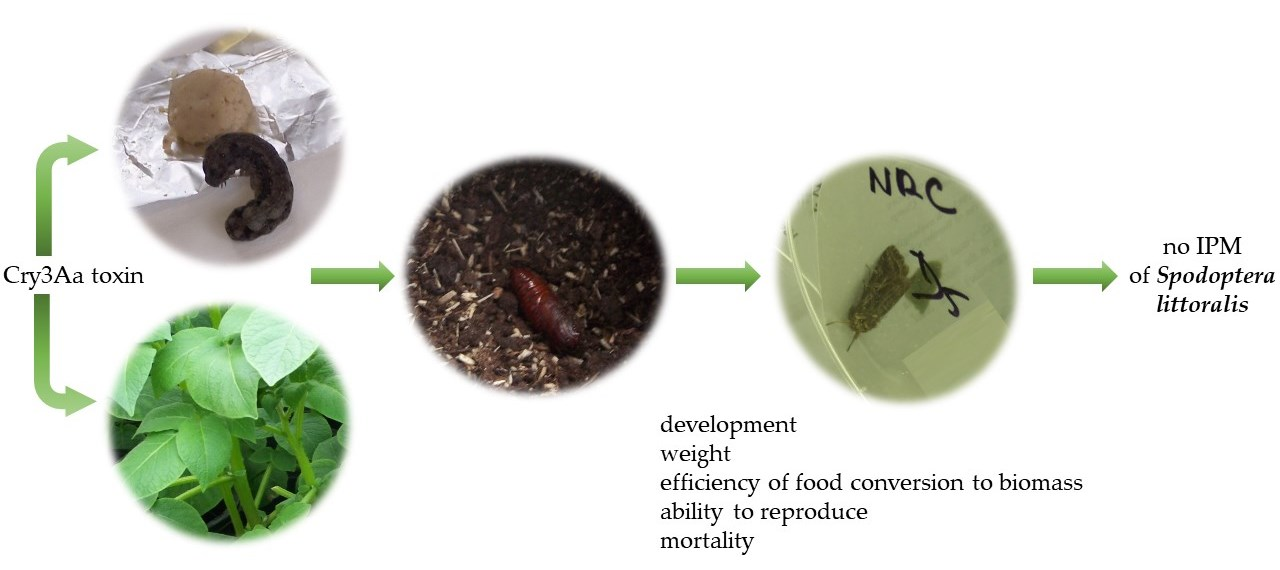
Cry3Aa Toxin Is Not Suitable to Control Lepidopteran Pest Spodoptera littoralis (Boisd.)


Abstract
:
1. Introduction
2. Results
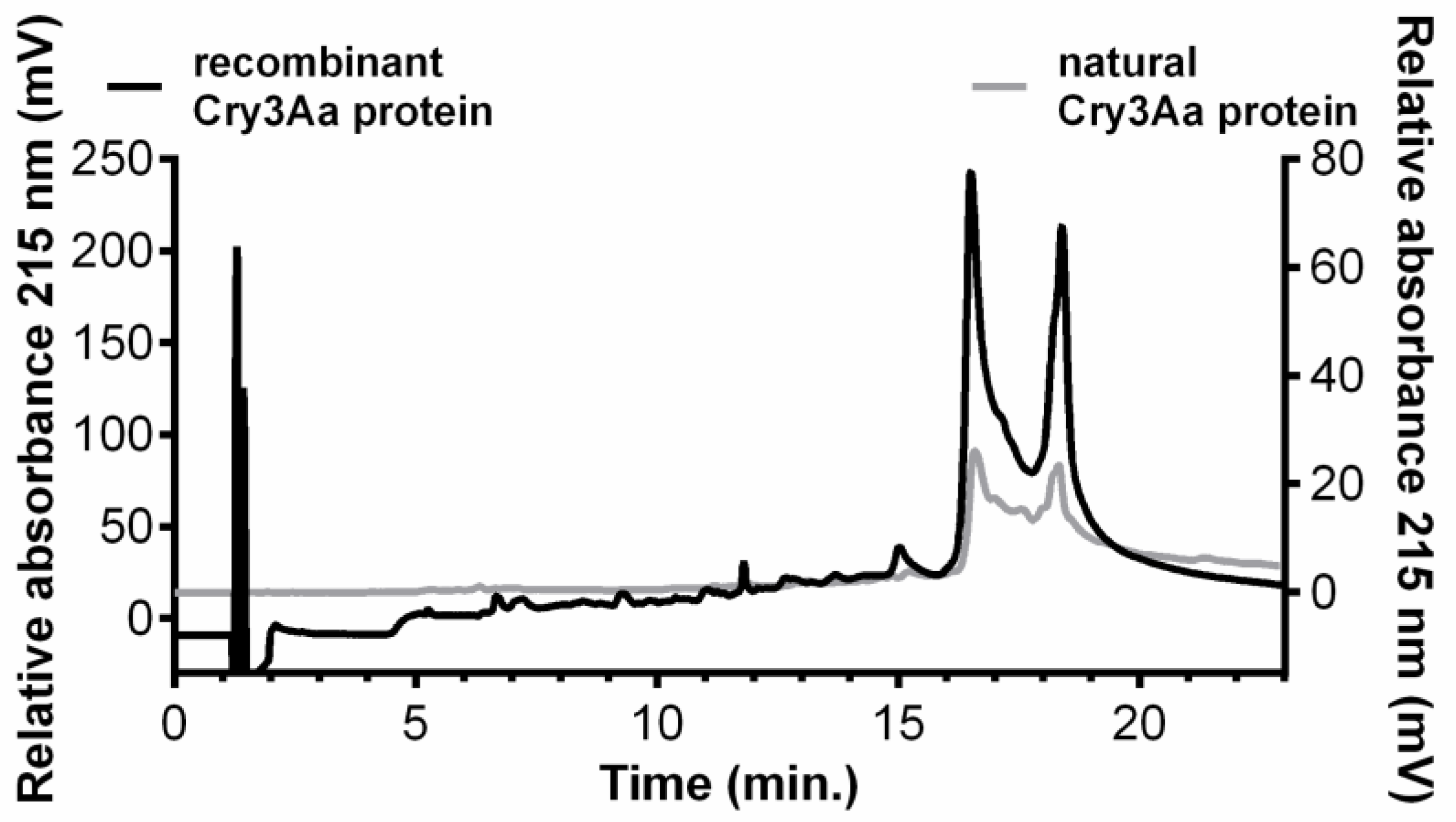
2.1. Cry3Aa Content
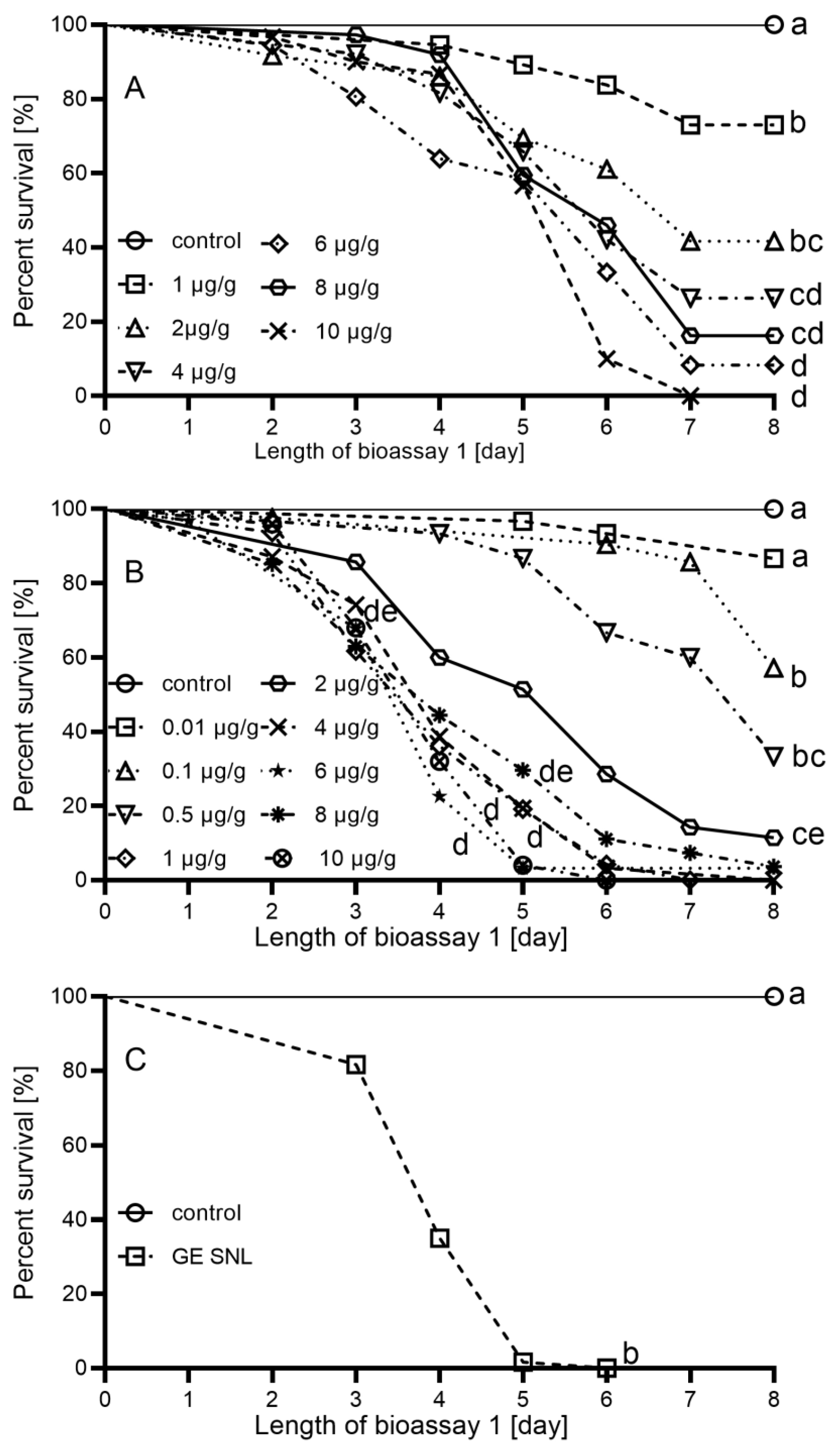
2.2. Effect of Cry3Aa on Survival of L. decemlineata
2.3. Effect of Cry3Aa on Larval and Pupal Mortality of S. littoralis
2.4. Sublethal Effects of Cry3Aa on S. littoralis
3. Discussion
3.1. Bioassay 1—Effect of Cry3Aa on L. decemlineata
3.2. Bioassay 2—Effect of Cry3Aa in Semi-Artificial Diet on S. littoralis
3.3. Bioassay 3—Effect of Cry3Aa Expressed in GE Potato SNL on S. littoralis
3.4. Larval and Pupal Mortality (Survival) of S. littoralis in Cry3Aa Treatments
3.5. Sublethal Effects of Cry3Aa on Different S. littoralis Populations
4. Conclusions
5. Materials and Methods
5.1. Culture of Leptinotarsa Decemlineata
5.2. Cultures of Spodoptera Littoralis
5.3. Semi-Artificial Diets
5.4. Origin of Cry3Aa Toxins
5.5. Quantification of Cry3Aa Toxins
5.6. Bioassays
5.7. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bravo, A.; Gomez, I.; Porta, H.; Garcia-Gomez, B.I.; Rodriguez-Almazan, C.; Pardo, L.; Soberon, M. Evolution of Bacillus thuringiensis Cry toxins insecticidal activity. Microb. Biotechnol. 2013, 6, 17–26. [Google Scholar] [CrossRef]
- Jurat-Fuentes, J.L.; Crickmore, N. Specificity determinants for Cry insecticidal proteins: Insights from their mode of action. J. Invertebr. Pathol. 2017, 142, 5–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, W.; Zhu, L.; Liu, Y.; Crickmore, N.; Peng, D.; Ruan, L.; Sun, M. Mining new crystal protein genes from Bacillus thuringiensis on the basis of mixed plasmid-enriched genome sequencing and a computational pipeline. Appl. Microbiol. Biotechnol. 2012, 78, 4795–4801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Frankenhuyzen, K. Cross-order and cross-phylum activity of Bacillus thuringiensis pesticidal proteins. J. Invertebr. Pathol. 2013, 114, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Vojtech, E.; Meissle, M.; Poppy, G.M. Effects of Bt maize on the herbivore Spodoptera littoralis (Lepidoptera: Noctuidae) and the parasitoid Cotesia marginiventris (Hymenoptera: Braconidae). Transgenic Res. 2005, 14, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Alfazairy, A.A.; El-Ahwany, A.M.; Mohamed, E.A.; Zaghloul, H.A.; El-Helow, E.R. Microbial control of the cotton leafworm Spodoptera littoralis (Boisd.) by Egyptian Bacillus thuringiensis isolates. Folia Microbiol. 2013, 58, 155–162. [Google Scholar] [CrossRef]
- BenFarhat-Touzri, D.; Saadaoui, M.; Abdelkefi-Mesrati, L.; Saadaoui, I.; Azzouz, H.; Tounsi, S. Histopathological effects and determination of the putative receptor of Bacillus thuringiensis Cry1Da toxin in Spodoptera littoralis midgut. J. Invertebr. Pathol. 2013, 112, 142–145. [Google Scholar] [CrossRef]
- Krieg, A. Bacillus Thuringiensis—Ein Mikrobielles Insekticid. Grundlagen und Anwendung (Acta Phytomedica, Heft 10); DM 58,00; Paul Parey: Berlin/Hamburg, Germany, 1986; 191p, ISBN 3-489-60826-7. [Google Scholar] [CrossRef]
- Keller, B.; Langerbruch, G.A. Control of coleopteran pests by Bacillus thuringiensis. In Bacillus Thuringiensis, an Environmental Biopesticide: Theory and Practice; Entwistle, P.F., Cory, J.S., Bailey, M.J., Higgs, S., Eds.; The Quarterly Review of Biology; Wiley: Chichester, UK, 1993; Volume 69, pp. 171–191. [Google Scholar] [CrossRef]
- Knowles, B.H. Mechanism of action of Bacillus thuringiensis insecticidal δ-endotoxins. Adv. Insect Physiol. 1994, 24, 275–308. [Google Scholar] [CrossRef]
- Deml, R.; Meise, T.; Dettner, K. Effects of Bacillus thuringiensis δ-endotoxins on food utilization, growth, and survival of selected phytophagous insects. J. Appl. Entomol. 1999, 123, 55–64. [Google Scholar] [CrossRef]
- Hussein, H.M.; Habuštová, O.; Sehnal, F. Beetle-specific Bacillus thuringiensis Cry3Aa toxin reduces larval growth and curbs reproduction in Spodoptera littoralis (Boisd.). Pest Manag. Sci. 2005, 61, 1186–1192. [Google Scholar] [CrossRef]
- Hussein, H.M.; Sehnal, F.; Habuštová, O. Bt-potatoes resistant to Colorado potato beetle affect the performance of Egyptian armyworm (Spodoptera littoralis). Acta Fytotech. Et Zootech. 2005, 8, 38–41. [Google Scholar]
- Hussein, H.M.; Habuštová, O.; Turanli, F.; Sehnal, F. Potato Expressing beetle-specific Bacillus thuringiensis Cry3Aa toxin reduces performance of a moth. J. Chem. Ecol. 2006, 32, 1–13. [Google Scholar] [CrossRef] [PubMed]
- European and Mediterranean Plant Protection Organization [OEPP/EPPO]. Diagnostic protocol for Spodoptera littoralis, Spodoptera litura, Spodoptera frugiperda, Spodoptera eridania. OEPP/EPPO Bull. 2015, 34, 257–270. [Google Scholar]
- Lopez-Vaamonde, C. Species accounts of 100 of the most invasive alien species in Europe. In Handbook of Alien Species in Europe; Springer: Dordrecht, The Netherlands, 2008; Volume 3, pp. 269–374. [Google Scholar] [CrossRef]
- Reed, G.L.; Jensen, A.S.; Riebe, J.; Head, G.; Duan, J.J. Transgenic Bt potato and conventional insecticides for Colorado potato beetle management: Comparative efficacy and non-target impacts. Entomol. Exp. Et Appl. 2001, 100, 89–100. [Google Scholar] [CrossRef]
- Duncan, D.; Hammond, D.; Zalewski, J.; Cudnohufsky, J.; Kaniewski, W.; Thornton, M.; Bookout, J.; Lavrik, P.; Rogan, G.; Feldman-Riebe, J. Field performance of transgenic potato, with resistance to Colorado potato beetle and viruses. HortScience 2002, 37, 275–276. [Google Scholar] [CrossRef] [Green Version]
- Ferry, N.; Mulligan, E.A.; Majerus, M.E.N.; Gatehouse, A.M.R. Bitrophic and tritrophic effects of Bt Cry3A transgenic potato on beneficial, non-target, beetles. Transgenic Res. 2007, 16, 795–812. [Google Scholar] [CrossRef]
- Latham, J.R.; Love, M.; Hilbeck, A. The distinct properties of natural and GM cry insecticidal proteins. Biotechnol. Genet. Eng. Rev. 2017, 33, 62–96. [Google Scholar] [CrossRef] [Green Version]
- Robertson, J.L.; Preisler, H.K.; Ng, S.S.; Hickle, L.A.; Gelernter, W.D. Natural variation: A complicating factor in bioassay with chemical and microbial pesticides. J. Econ. Entomol. 1995, 88, 1–10. [Google Scholar] [CrossRef]
- Robertson, J.L.; Russell, R.M.; Preisler, H.K.; Savin, N.E. Bioassays with Arthropods, 2nd ed.; CRC Press: Boca Raton, FL, USA; Taylor and Francis Group: Boca Raton, FL, USA, 2007; 199p. [Google Scholar]
- Perlak, F.J.; Stone, T.B.; Muskopf, Y.M.; Petersen, L.J.; Parker, G.B.; McPherson, S.A.; Wyman, J.; Love, S.; Reed, G.; Biever, D.; et al. Genetically improved potatoes: Protection from damage by Colorado potato beetles. Plant Mol. Biol. 1993, 22, 313–321. [Google Scholar] [CrossRef]
- Whalon, M.E.; Wierenga, J.M. Bacillus thuringiensis resistant Colorado potato beetle and transgenic plants: Some operational and ecological implications for deployment. Biocontrol Sci. Technol. 1994, 4, 555–561. [Google Scholar] [CrossRef]
- Grafius, E.J.; Douches, D.S. The present and future role of insect-resistant genetically modified potato cultivars in IPM. In Integration of Insect-Resistant Genetically Modified Crops within IPM Programs, 1st ed.; Springer: Dordrecht, The Netherlands, 2008; pp. 195–221. [Google Scholar]
- International Service for the Acquisition of Agri-Biotech Applications [ISAAA]. Global Status of Commercialized Biotech/GM Crops ISAAA Brief No. 55, ISAAA, Ithaca, USA. 2019. Available online: https://www.isaaa.org/resources/publications/briefs/55/ (accessed on 1 April 2022).
- Haider, M.Z.; Ellar, D.J. Mechanism of action of Bacillus thuringiensis insecticidal δ-endotoxin: Interaction with phospholipid vesicles. Biochim. Et Biophys. Acta (BBA) Biomenbrane 1989, 978, 216–222. [Google Scholar] [CrossRef]
- Spies, A.G.; Spence, K.D. Effect of sublethal Bacillus thuringiensis crystal endotoxin treatment on the larval midgut of a moth, Manduca sexta—SEM study. Tissue Cell 1985, 17, 379–394. [Google Scholar] [CrossRef]
- Peacock, J.W.; Schweitzer, D.F.; Carter, J.L.; Dubois, N.R. Laboratory assessment of the effects of Bacillus thuringiensis on native Lepidoptera. Environ. Entomol. 1998, 27, 450–457. [Google Scholar] [CrossRef]
- Moreau, G.; Bauce, E. Developmental polymorphism: A major factor for understanding sublethal effects of Bacillus thuringiensis. Entomol. Exp. Appl. 2001, 98, 133–140. [Google Scholar] [CrossRef] [Green Version]
- Sneh, B.; Schuster, S.; Broza, M. Insecticidal activity of Bacillus thuringiensis strains against the Egyptian cotton leafworm, Spodoptera littoralis Boisd. (Lep. Noctuidae). Entomophaga 1981, 26, 179–190. [Google Scholar] [CrossRef]
- Keller, M.; Sneh, B.; Strizhov, N.; Prudovsky, E.; Regev, A.; Koncz, C.; Schell, J.; Zilberstein, A. Digestion of δ-endotoxin by gut proteases may explain reduced sensitivity of advanced instar larvae of Spodoptera littoralis to CryIC. Insect Biochem. Mol. Biol. 1996, 26, 365–373. [Google Scholar] [CrossRef]
- Mohamed, S.A.; Badr, N.A.; El-Hafez, A.A. Efficacy of two formulations of pathogenic bacteria Bacillus thuringiensis against the first instar larvae of Spodoptera littoralis (Boisd.) and Agrotis ipsilon (Han.) (Lepidoptera: Noctuidae). Egypt. J. Agric. Res. 2000, 78, 1025–1040. [Google Scholar]
- Porcar, M.; Caballero, P. Molecular and insecticidal characterization of a Bacillus thuringiensis strain isolated during a natural epizootic. J. Appl. Microbiol. 2000, 89, 309–316. [Google Scholar] [CrossRef] [Green Version]
- Romeilah, M.; Abdel-Megeed, M.A. The role of certain bacterial preparations (Bacillus thuringiensis) in controlling the cotton leafworm Spodoptera littoralis (Boisd.). Egypt. J. Agric. Res. 2000, 78, 1877–1887. [Google Scholar]
- Dutton, A.; Klein, H.; Romeis, J.; Bigler, F. Prey-mediated effects of Bacillus thuringiensis spray on the predator Chrysoperla carnea in maize. Biol. Control 2003, 26, 209–215. [Google Scholar] [CrossRef]
- Svobodová, Z.; Shu, Y.; Skoková Habuštová, O.; Romeis, J.; Meissle, M. Stacked Bt maize and arthropod predators—Exposure to insecticidal Cry proteins and potential hazards. Proceeding R. Soc. B Biol. Sci. 2017, 284, 20170440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Romeis, J.; Meissle, M. Performance of Daphnia magna on flour, leaves, and pollen from different maize lines: Implications for risk assessment of genetically engineered crops. Ecotoxicol. Environ. Saf. 2021, 212, 111967. [Google Scholar] [CrossRef] [PubMed]
- Chestukhina, G.G.; Kostina, L.I.; Zalunin, I.A.; Kotova, T.S.; Katruka, S.P.; Kuznetsov, Y.S.; Stepanov, V.M. Proteins of Bacillus thuringiensis delta-endotoxin crystals. Biokhimiia 1977, 42, 1660–1667. [Google Scholar] [PubMed]
- Dunn, O.J. Multiple comparisons among means. J. Am. Stat. Assoc. 1961, 56, 52–64. Available online: http://www.jstor.org/stable/2282330 (accessed on 28 March 2022). [CrossRef]
- Robertson, J.L.; Preisler, H.K.; Russell, R.M. PoloPlus Probit and Logit Analysis USER’S GUIDE; CRC Press: Boca Raton, FL, USA, 2002–2003; p. 36. [Google Scholar]
- StatSoft Inc. Statistica Electronic Manual; StatSoft Inc.: Tulsa, OK, USA, 2015; Available online: http://documentation.statsoft.com/STATISTICAHelp.aspx?path=common/AboutSTATISTICA/ElectronicManualIndex (accessed on 8 March 2018).
- GraphPad Software Inc. GraphPad Prism 5.0 User´s Guide; GraphPad Software Inc.: San Diego, CA, USA, 2007; Available online: https://www.graphpad.com/guides/prism/7/user-guide/index.htm?citing_graphpad_prism.htm (accessed on 15 May 2020).





{kind=link}
{kind=link}
{kind=link}
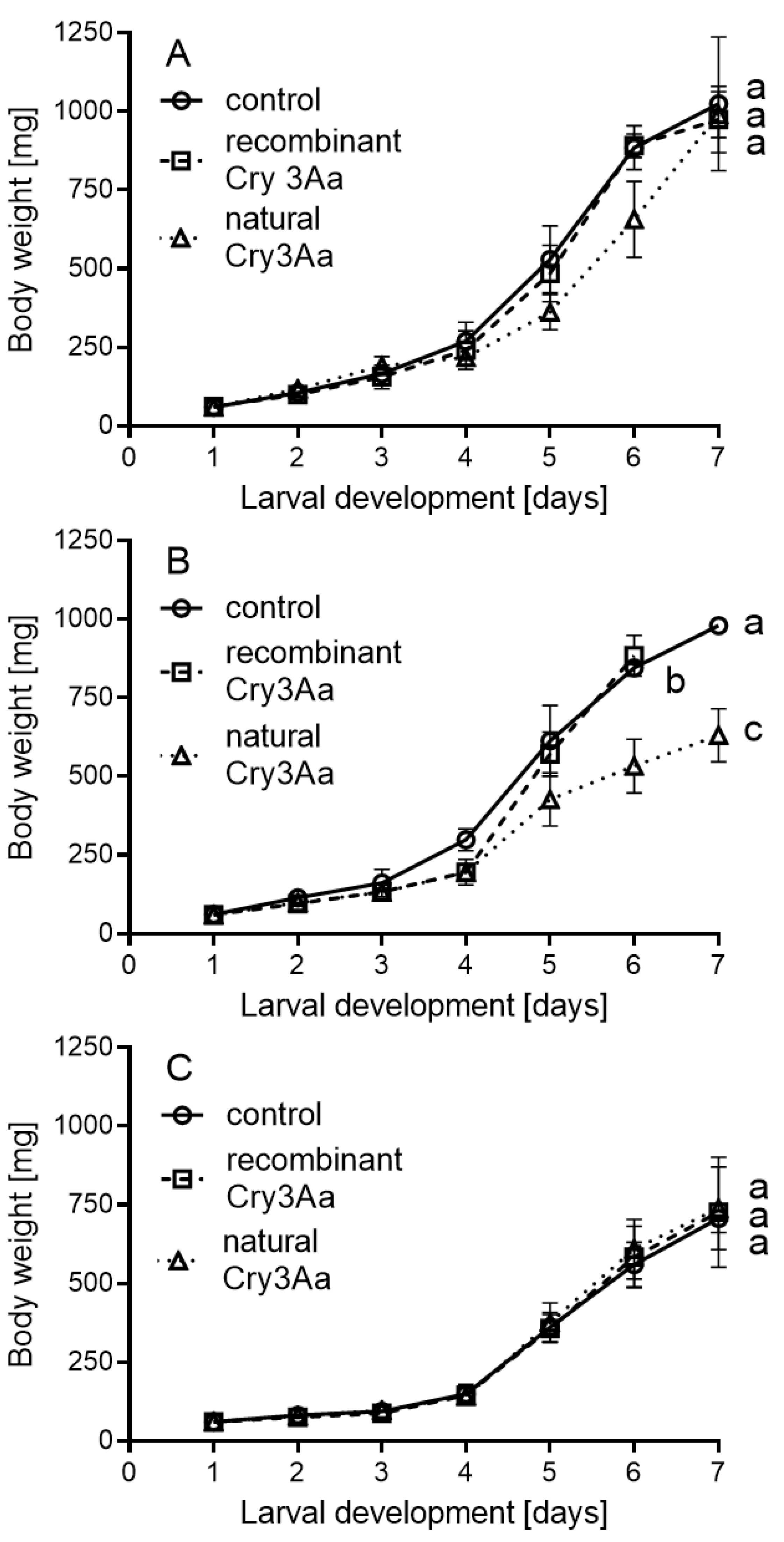{kind=link}
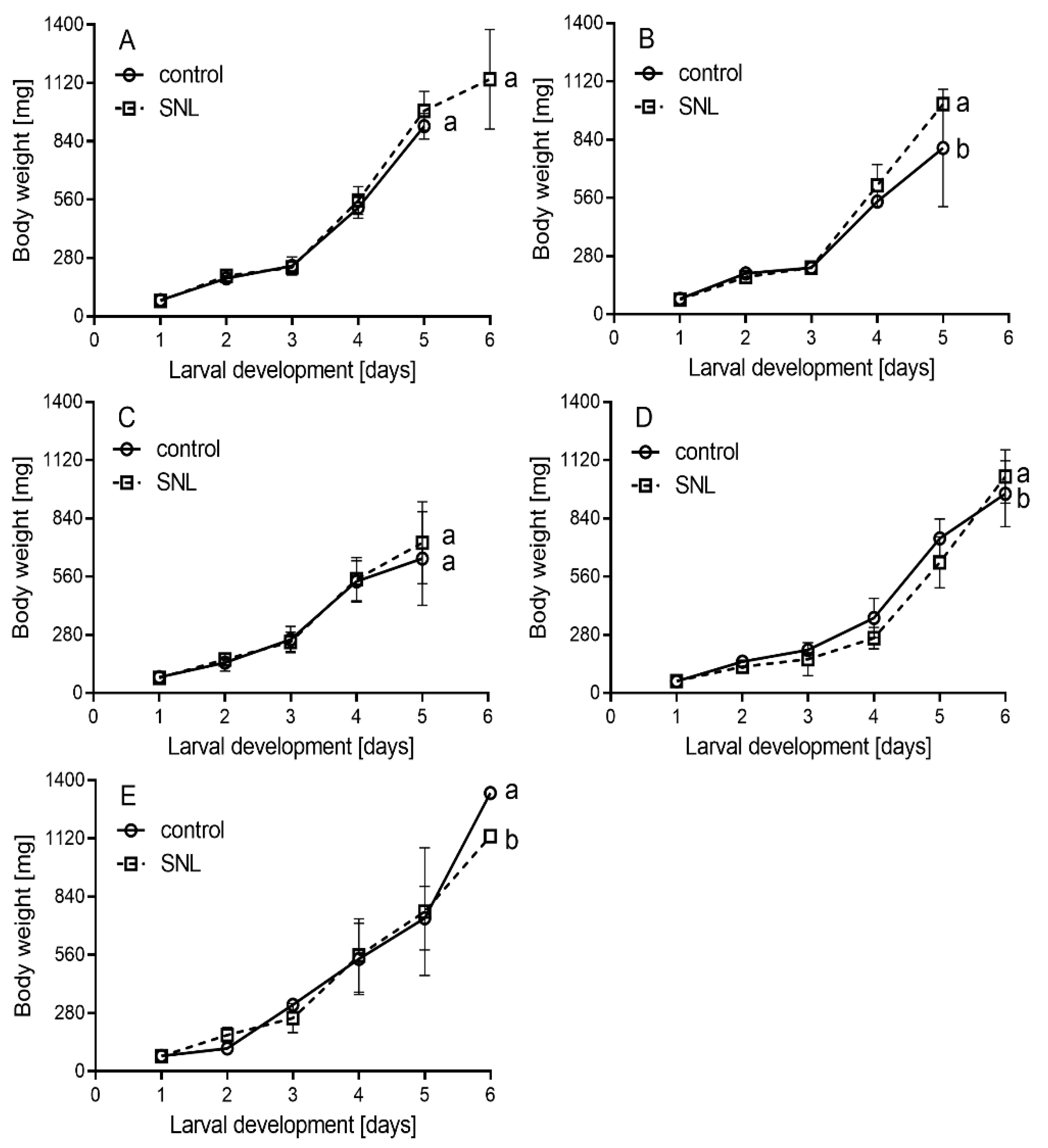{kind=link}
| Population | Examined Parameter | Control | Recombinant Cry3Aa | Natural Cry3Aa |
|---|---|---|---|---|
| NRC | Initial larval weight (mg) | 59.1 ± 3.3 | 60.3 ± 4.0 | 59.2 ± 3.5 |
| ECI (%) 1 | 24.2 ± 1.7 | 36.5 ± 1.9 | 31.8 ± 11.3 | |
| Weight increment (mg) | 950.0 ± 176.8 b | 1077.7 ± 149.5 a | 945.8 ± 153.1 b | |
| Maximal body weight (mg) | 1009.1 ± 176.4 b | 1137.9 ± 148.7 a | 1004.9 ± 152.9 b | |
| Length of fifth instar (days) | 2.9 ± 0.4 | 2.9 ± 0.3 | 2.9 ± 0.3 | |
| Length of sixth instar (days) | 2.9 ± 0.4 | 2.8 ± 0.4 | 2.9 ± 0.4 | |
| Length of prepupal stage (days) | 2.6 ± 0.6 | 2.7 ± 0.5 | 2.5 ± 0.5 | |
| Larval mortality (%) | 0 | 0 | 7.5 | |
| Pupal weight (mg) | 363.4 ± 52.5 b | 386.9 ± 45.9 a | 366.2 ± 40.4 b | |
| Length of pupal stage (days) | 8.7 ± 0.7 b | 10.2 ± 0.7 a | 8.5 ± 0.5 c | |
| Pupal mortality (%) | 0 | 0 | 8.1 | |
| No. of laid eggs per female per day | 281.2 ± 125.6 | 294.7 ± 121.3 | 274.6 ± 127.8 | |
| No. of hatched eggs per female per day | 136.1 ± 79.4 | 140.2 ± 127.9 | 147.1 ± 108.3 | |
| Hatching rate per female per day (%) | 43.8 ± 17.0 | 36.9 ± 30.4 | 50.2 ± 22.0 | |
| SE | Initial larval weight (mg) | 58.6 ± 3.6 | 59.7 ± 3.2 | 58.0 ± 2.8 |
| ECI (%) | 25.0 ± 7.5 | 28.7 ± 1.8 | 20.2 ± 3.8 | |
| Weight increment (mg) | 825.6 ± 104.8 a | 800.1 ± 114.6 a | 709.7 ± 177.0 b | |
| Maximal body weight (mg) | 884.3 ± 103.4 a | 859.8 ± 114.0 a | 767.7 ± 176.4 b | |
| Length of fifth instar (days) | 2.6 ± 0.5 a | 2.2 ±0.4 b | 2.6 ± 0.5 a | |
| Length of sixth instar (days) | 2.9 ± 0.4 | 3.1 ± 0.5 | 3.1 ± 0.5 | |
| Length of prepupal stage (days) | 2.5 ± 0.5 | 2.4 ± 0.6 | 2.7 ± 0.8 | |
| Larval mortality (%) | 20.0 | 32.5 | 32.5 | |
| Pupal weight (mg) | 327.2 ± 42.3 a | 303.9 ± 43.8 ab | 299.0 ± 52.9 b | |
| Length of pupal stage (days) | 8.5 ± 0.7 | 8.2 ± 0.9 | 8.3 ± 0.8 | |
| Pupal mortality (%) | 6.3 | 0 | 3.7 | |
| No. of laid eggs per female per day | 298.9 ± 132.5 | 328.8 ± 148 | 252.2 ± 123.9 | |
| No. of hatched eggs per female per day | 256.9 ± 150.3 | 292.2 ± 170.8 | 214.3 ± 135.5 | |
| Hatching rate per female per day (%) | 75.8 ± 29.5 | 81.8 ± 23 | 72.9 ± 34.9 | |
| SF | Initial larval weight (mg) | 59.9 ± 3.1 | 60.2 ± 3.5 | 59.3 ± 3.3 |
| ECI (%) | 21.4 ± 2.8 | 22.2 ± 3.4 | 23.2 ± 1.8 | |
| Weight increment (mg) | 861.8 ± 211.9 | 822.9 ± 131.3 | 811.1 ± 138.0 | |
| Maximal body weight (mg) | 921.7 ± 212.2 | 883.3 ± 131.2 | 870.4 ± 138.7 | |
| Length of fifth instar (days) | 2.3 ± 0.5 | 2.2 ± 0.4 | 2.2 ± 0.4 | |
| Length of sixth instar (days) | 3.1 ± 0.4 | 3.1 ± 0.5 | 3.0 ± 0.4 | |
| Length of prepupal stage (days) | 2.7 ± 0.5 | 2.9 ± 0.5 | 2.8 ± 0.5 | |
| Larval mortality (%) | 17.5 | 15.0 | 25.0 | |
| Pupal weight (mg) | 330.5 ± 46.1 | 326.6 ± 39.1 | 317.4 ± 41.7 | |
| Length of pupal stage (days) | 8.2 ± 0.8 ab | 8.1 ± 0.7 b | 8.6 ± 1.3 a | |
| Pupal mortality (%) | 15.2 | 11.8 | 3.3 | |
| No. of laid eggs per female per day | 245.8 ± 152.1 | 334.4 ± 180.6 | 259.0 ± 133.2 | |
| No. of hatched eggs per female per day | 157.1 ± 118.9 | 227.3 ± 179.6 | 165.9 ± 120.2 | |
| Hatching rate per female per day (%) | 48.5 ± 30.1 | 61.0 ± 29.8 | 52.4 ± 32.7 |
| Population | Examined Parameter | Control | GE Potato SNL |
|---|---|---|---|
| N | Initial larval weight (mg) | 75.7 ± 2.4 | 74.7 ± 2.5 |
| ECI (%) 1 | 24.1 ± 12.1 | 23.1 ± 5.1 | |
| Weight increment (mg) | 964.6 ± 141.5 | 933.3 ± 170.5 | |
| Maximal body weight (mg) | 1040.3 ± 141.6 | 1008.0 ± 170.5 | |
| Length of fifth instar (days) | 2.4 ± 0.5 | 2.2 ± 0.4 | |
| Length of sixth instar (days) | 3.5 ± 0.6 | 3.4 ± 0.6 | |
| Length of prepupal stage (days) | 1.7 ± 0.6 | 1.9 ± 0.6 | |
| Larval mortality (%) | 33.3 | 43.3 | |
| Pupal weight (mg) | 341.3 ± 30.2 | 326.1 ± 43.3 | |
| Length of pupal stage (days) | 10.4 ± 0.7 | 10.4 ± 0.6 | |
| Pupal mortality (%) | 02 | 0 | |
| No. of laid eggs per female per day | 217.3 ± 96.6 | 260.0 ± 142.5 | |
| No. of hatched eggs per female per day | 144.4 ± 130.3 | 150.6 ± 147.6 | |
| Hatching rate per female per day (%) | 60.8 ± 21.5 | 46.9 ± 29.6 | |
| NRC | Initial larval weight (mg) | 72.7 ± 2.4 | 71.7 ± 1.8 |
| ECI (%) | 24.0 ± 5.7 | 33.1 ± 16.1 | |
| Weight increment (mg) | 872.7 ± 125.6 | 913.6 ± 109.4 | |
| Maximal body weight (mg) | 945.4 ± 125.2 | 985.3 ± 110.0 | |
| Length of fifth instar (days) | 2.0 ± 02 | 2.0 ± 0 | |
| Length of sixth instar (days) | 3.4 ± 0.6 a | 3.0 ± 0 b | |
| Length of prepupal stage (days) | 2.6 ± 0.5 | 2.4 ± 0.5 | |
| Larval mortality (%) | 36.7 | 53.3 | |
| Pupal weight (mg) | 333.3 ± 17.8 | 340.1 ± 25.5 | |
| Length of pupal stage (days) | 8.2 ± 0.4 b | 9.4 ± 0.7 a | |
| Pupal mortality (%) | 02 | 0 | |
| No. of laid eggs per female per day | 297.5 ± 70.4 | 376.2 ± 152.7 | |
| No. of hatched eggs per female per day | 232.8 ± 94.3 | 262.7 ± 159.1 | |
| Hatching rate per female per day (%) | 74.5 ± 20.2 | 67.9 ± 31.2 | |
| CU | Initial larval weight (mg) | 74.5 ± 3.3 | 73.9 ± 3.3 |
| ECI (%) | 17.9 ± 4.2 | 19.8 ± 4.7 | |
| Weight increment (mg) | 703.5 ± 204.7 | 740.6 ± 138.9 | |
| Maximal body weight (mg) | 778.0 ± 203.5 | 814.5 ± 138.0 | |
| Length of fifth instar (days) | 2.0 ± 0.2 | 2.1 ± 0.3 | |
| Length of sixth instar (days) | 3.0 ± 0.2 | 2.9 ± 0.4 | |
| Length of prepupal stage (days) | 1.7 ± 0.5 | 1.9 ± 0.3 | |
| Larval mortality (%) | 40.0 | 46.7 | |
| Pupal weight (mg) | 287.4 ± 39.9 | 288.3 ± 42.6 | |
| Length of pupal stage (days) | 8.1 ± 0.9 | 7.6 ± 0.8 | |
| Pupal mortality (%) | 02 | 0 | |
| No. of laid eggs per female per day | 348.6 ± 108.3 | 300.4 ± 73.1 | |
| No. of hatched eggs per female per day | 321.9 ± 110.1 | 217.1 ± 78.0 | |
| Hatching rate per female per day (%) | 91.8 ± 4.0 | 71.5 ± 14.9 | |
| SE | Initial larval weight (mg) | 60.8 ± 6.8 | 61.6 ± 6.6 |
| ECI (%) | 24.2 ± 4.0 | 25.6 ± 4.1 | |
| Weight increment (mg) | 868.3 ± 145. | 912.2 ± 148.4 | |
| Maximal body weight (mg) | 929.1 ± 142.8 | 973.8 ± 149.9 | |
| Length of fifth instar (days) | 2.3 ± 0.7 | 2.3 ± 0.5 | |
| Length of sixth instar (days) | 3.8 ± 0.8 | 3.8 ± 0.5 | |
| Length of prepupal stage (days) | 1.4 ± 0.5 | 1.6 ± 0.5 | |
| Larval mortality (%) | 6.7 | 16.7 | |
| Pupal weight (mg) | 291.0 ± 52.2 | 298.8 ± 47.9 | |
| Length of pupal stage (days) | 8.8 ± 0.8 b | 9.3 ± 0.9 a | |
| Pupal mortality (%) | 3.6 | 8.3 | |
| No. of laid eggs per female per day | 322.9 ± 134.2 | 317.8 ± 76.9 | |
| No. of hatched eggs per female per day | 194.1 ± 94.6 | 161.7 ± 60.2 | |
| Hatching rate per female per day (%) | 60.2 ± 16.4 | 51.0 ± 17.7 | |
| SF | Initial larval weight (mg) | 73.6 ± 3.8 | 74.3 ± 3.9 |
| ECI (%) | 16.7 ± 3.6 | 21.0 ± 7.7 | |
| Weight increment (mg) | 769.9 ± 324.3 | 973.8 ± 294.2 | |
| Maximal body weight (mg) | 843.5 ± 325.1 | 1048.1 ± 293.0 | |
| Length of fifth instar (days) | 1.4 ± 0.5 a | 1.1 ± 0.3 b | |
| Length of sixth instar (days) | 3.8 ± 0.9 | 4.4 ± 1.0 | |
| Length of prepupal stage (days) | 1.3 ± 0.4 b | 1.4 ± 0.5 a | |
| Larval mortality (%) | 6.7 | 3.3 | |
| Pupal weight (mg) | 295.0 ± 81.1 | 351.1 ± 92.2 | |
| Length of pupal stage (days) | 10.7 ± 0.8 a | 10.1 ± 0.8 b | |
| Pupal mortality (%) | 57.1 a | 24.1 b | |
| No. of laid eggs per female per day | 332.9 ± 203.7 | 255.7 ± 146.7 | |
| No. of hatched eggs per female per day | 114.1 ± 145.4 | 150.9 ± 149.0 | |
| Hatching rate per female per day (%) | 32.2 ± 36.4 | 41.4 ± 39.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skoková Habuštová, O.; Svobodová, Z.; Kodrík, D.; Sehnal, F. Cry3Aa Toxin Is Not Suitable to Control Lepidopteran Pest Spodoptera littoralis (Boisd.). Plants 2022, 11, 1312. https://doi.org/10.3390/plants11101312
Skoková Habuštová O, Svobodová Z, Kodrík D, Sehnal F. Cry3Aa Toxin Is Not Suitable to Control Lepidopteran Pest Spodoptera littoralis (Boisd.). Plants. 2022; 11(10):1312. https://doi.org/10.3390/plants11101312
Chicago/Turabian StyleSkoková Habuštová, Oxana, Zdeňka Svobodová, Dalibor Kodrík, and František Sehnal. 2022. "Cry3Aa Toxin Is Not Suitable to Control Lepidopteran Pest Spodoptera littoralis (Boisd.)" Plants 11, no. 10: 1312. https://doi.org/10.3390/plants11101312
APA StyleSkoková Habuštová, O., Svobodová, Z., Kodrík, D., & Sehnal, F. (2022). Cry3Aa Toxin Is Not Suitable to Control Lepidopteran Pest Spodoptera littoralis (Boisd.). Plants, 11(10), 1312. https://doi.org/10.3390/plants11101312