
Phylogeny and Expression Atlas of the NITRATE TRANSPORTER 1/PEPTIDE TRANSPORTER FAMILY in Agave

 ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. Identification and Subcellular Localization of Agave NPF Genes
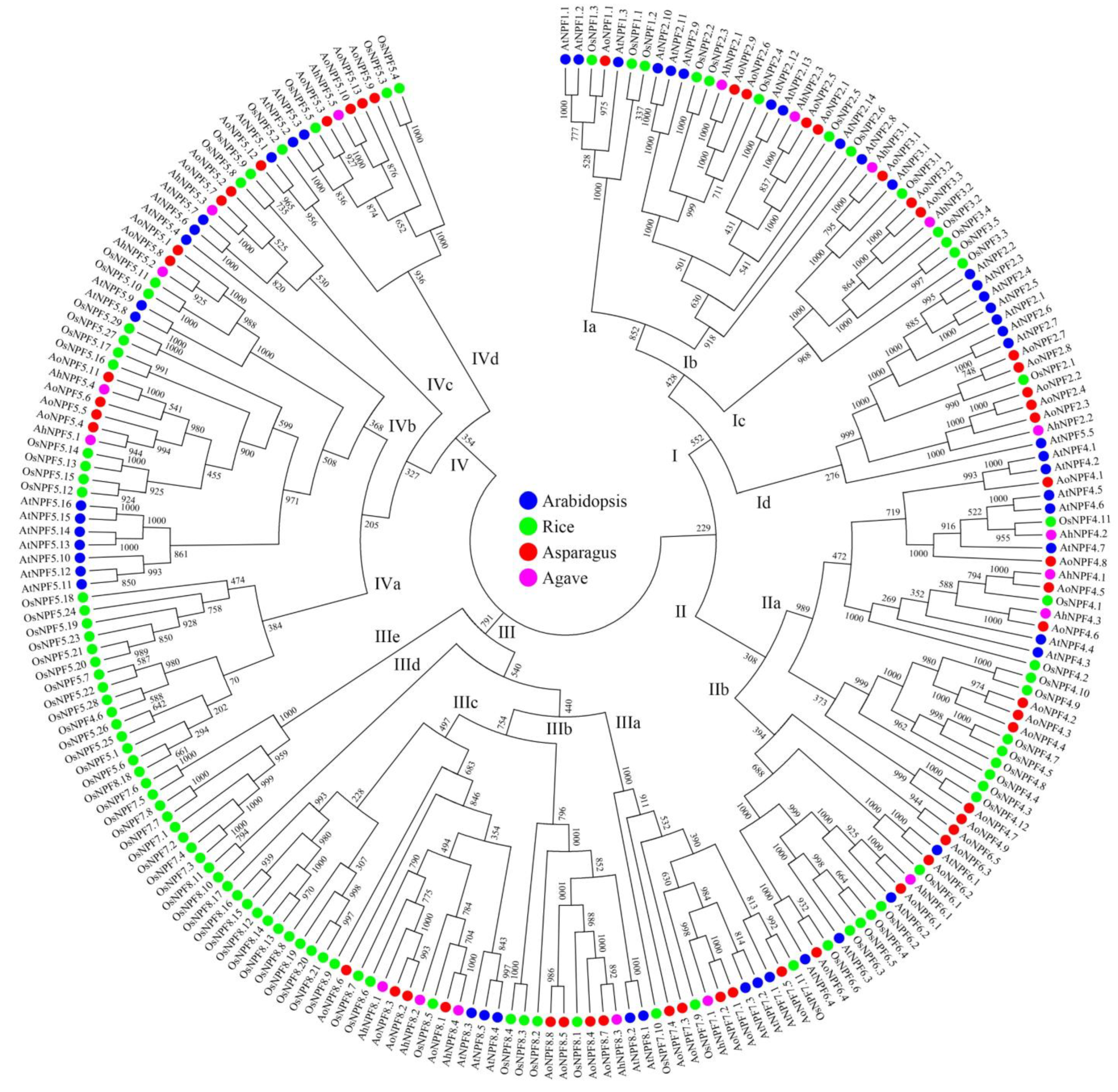
2.2. Phylogenetic Analysis of Agave NPF Genes
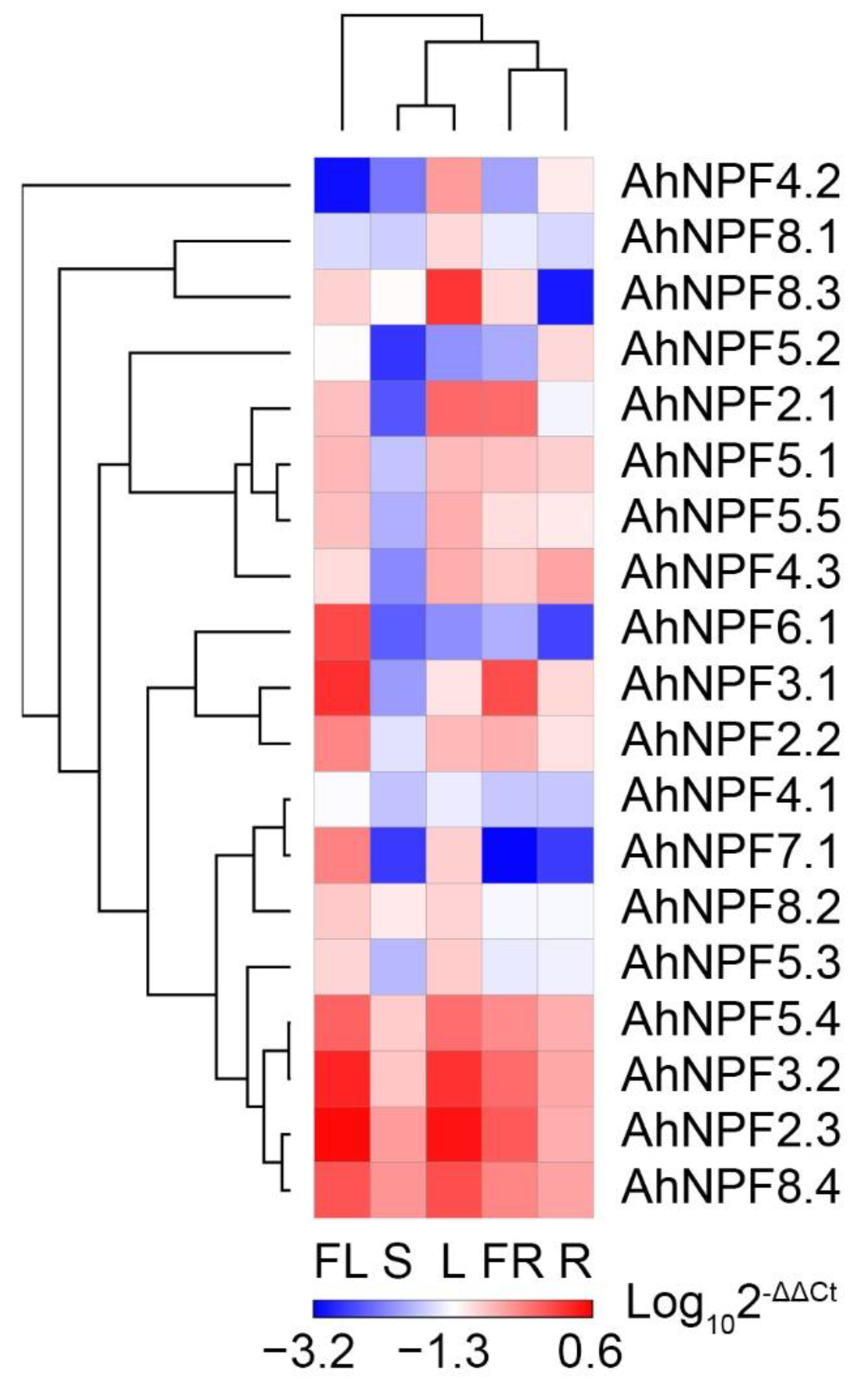
2.3. Expression Profiles of AhNPFs in Different Agave Tissues
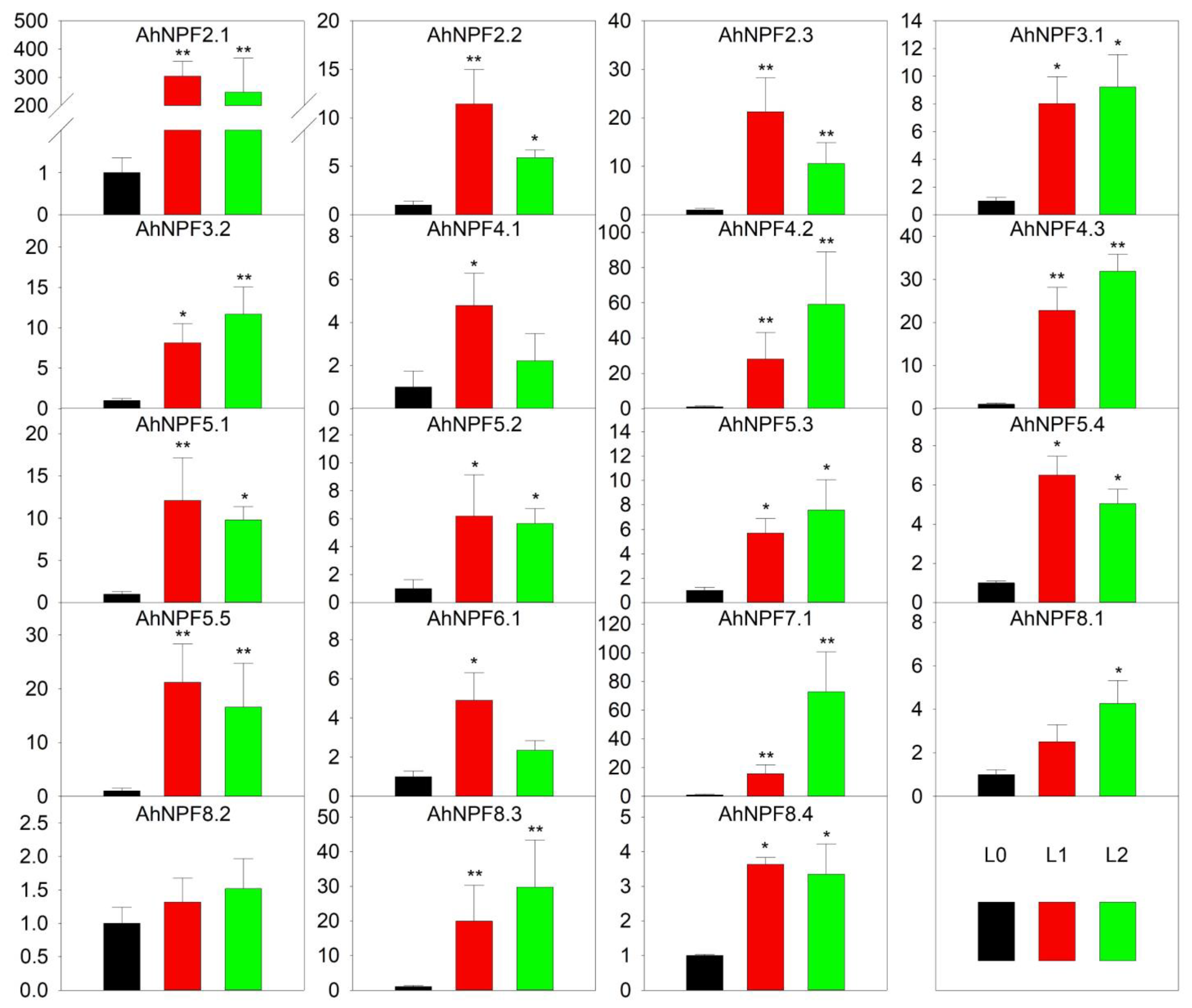
2.4. Expression Profiles of AhNPFs during Leaf Development
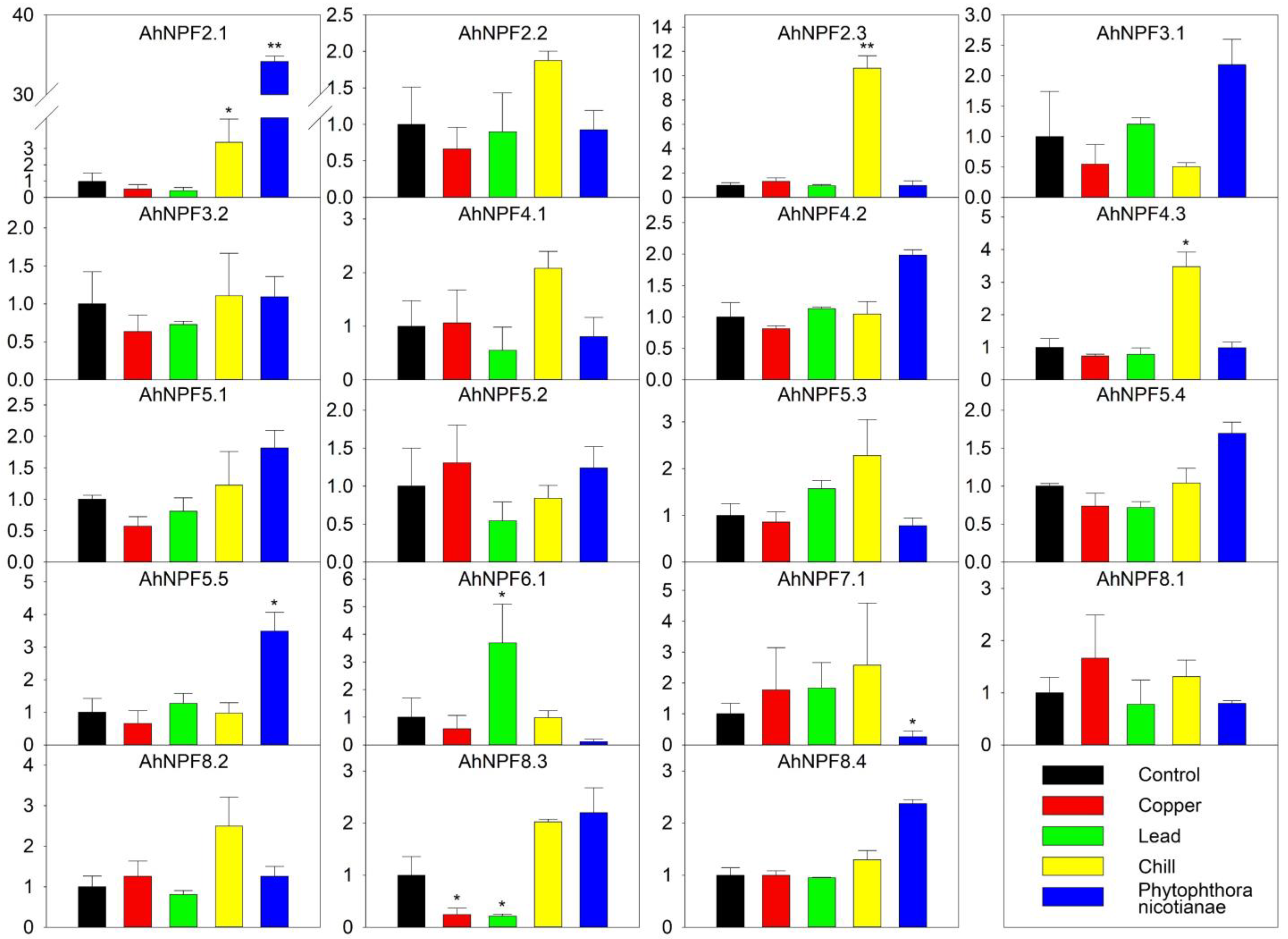
2.5. Expression Profiles of AhNPFs under Abiotic/Biotic Stresses
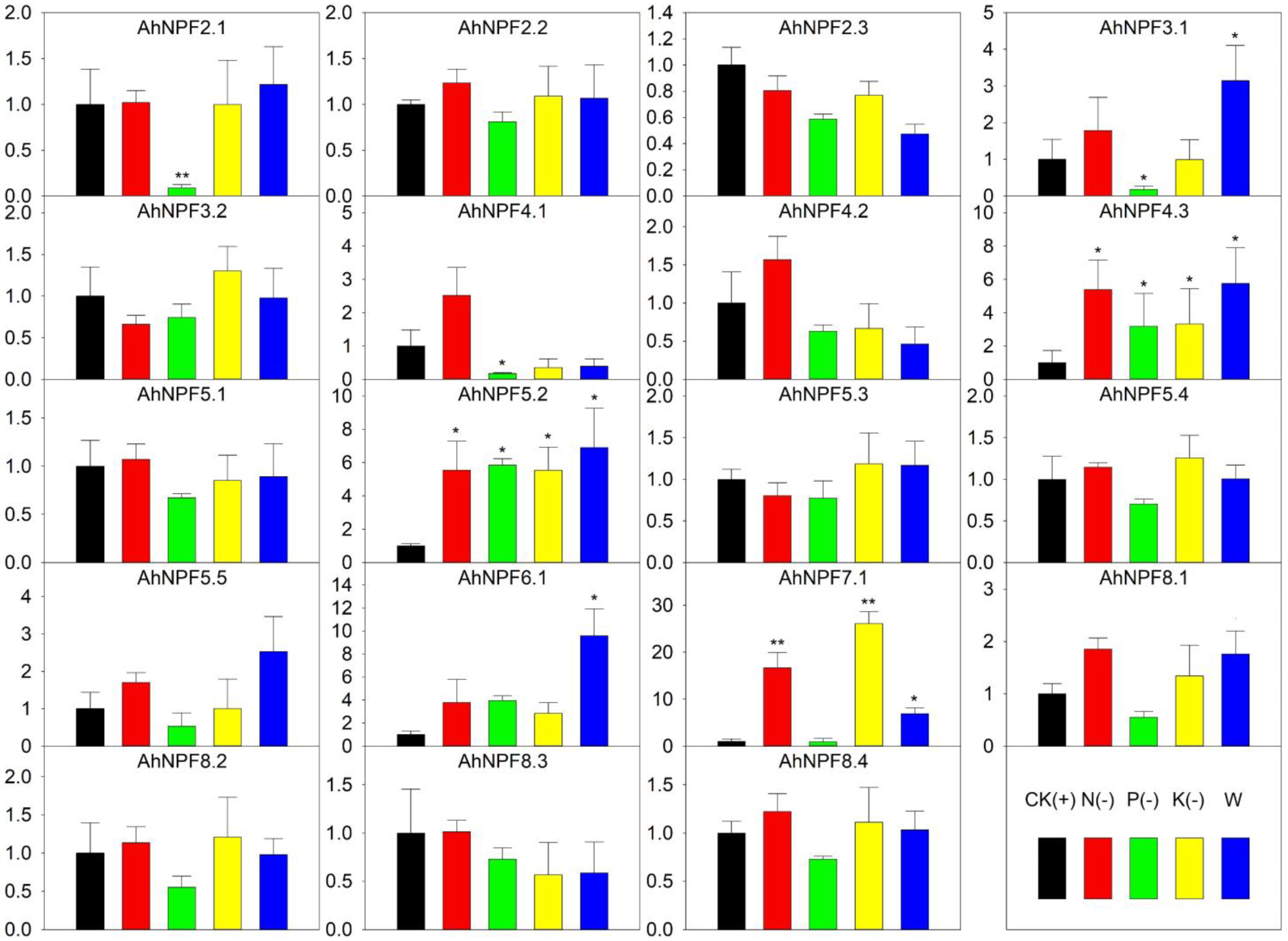
2.6. Expression Profiles of AhNPFs under Nutrient Deficiency
3. Discussion
3.1. Identification of Agave NPF Genes
3.2. Regulation of Agave NPF Genes Associated with Leaf Development, Abiotic/Biotic Stresses and Nutrient Deficiency
4. Materials and Methods
4.1. Sequence Retrieval and Phylogenetic Analysis
4.2. Plant Materials and RNA Extraction
4.3. Analysis of Gene Expression Using Quantitative Reverse Transcription PCR (qRT-PCR)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Crawford, N.M. Nitrate: Nutrient and signal for plant growth. Plant Cell 1995, 7, 859–868. [Google Scholar]
- Wang, Y.Y.; Cheng, Y.H.; Chen, K.E.; Tsay, Y.F. Nitrate Transport, Signaling, and Use Efficiency. Annu. Rev. Plant Biol. 2018, 69, 85–122. [Google Scholar] [CrossRef]
- Hsu, P.K.; Tsay, Y.F. Two phloem nitrate transporters, NRT1.11 and NRT1.12, are important for redistributing xylem-borne nitrate to enhance plant growth. Plant Physiol. 2013, 163, 844–856. [Google Scholar] [CrossRef] [Green Version]
- Taochy, C.; Gaillard, I.; Ipotesi, E.; Oomen, R.; Leonhardt, N.; Zimmermann, S.; Peltier, J.B.; Szponarski, W.; Simonneau, T.; Sentenac, H.; et al. The Arabidopsis root stele transporter NPF2.3 contributes to nitrate translocation to shoots under salt stress. Plant J. 2015, 83, 466–479. [Google Scholar] [CrossRef]
- Segonzac, C.; Boyer, J.C.; Ipotesi, E.; Szponarski, W.; Tillard, P.; Touraine, B.; Sommerer, N.; Rossignol, M.; Gibrat, R. Nitrate efflux at the root plasma membrane: Identification of an Arabidopsis excretion transporter. Plant Cell 2007, 19, 3760–3777. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.Y.; Tsay, Y.F. Arabidopsis nitrate transporter NRT1.9 is important in phloem nitrate transport. Plant Cell 2011, 23, 1945–1957. [Google Scholar] [CrossRef] [Green Version]
- Almagro, A.; Lin, S.H.; Tsay, Y.F. Characterization of the Arabidopsis nitrate transporter NRT1.6 reveals a role of nitrate in early embryo development. Plant Cell 2008, 20, 3289–3299. [Google Scholar] [CrossRef] [Green Version]
- Fan, S.; Lin, C.; Hsu, P.; Lin, S.; Tsay, Y. The Arabidopsis nitrate transporter NRT1.7, expressed in phloem, is responsible for source-to-sink remobilization of nitrate. Plant Cell 2009, 21, 2750–2761. [Google Scholar] [CrossRef] [Green Version]
- Pike, S.; Gao, F.; Kim, M.J.; Kim, S.H.; Schachtman, D.P.; Gassmann, W. Members of the NPF3 transporter subfamily encode pathogen-inducible nitrate/nitrite transporters in grapevine and Arabidopsis. Plant Cell Physiol. 2014, 55, 162–170. [Google Scholar] [CrossRef] [Green Version]
- Kanno, Y.; Kamiya, Y.; Seo, M. Nitrate does not compete with abscisic acid as a substrate of AtNPF4.6/NRT1.2/AIT1 in Arabidopsis. Plant Signal. Behav. 2013, 8, e26624. [Google Scholar] [CrossRef] [Green Version]
- L’eran, S.; Garg, B.; Boursiac, Y.; Corratg’e-Faillie, C.; Brachet, C.; Tillard, P.; Gojon, A.; Lacombe, B. AtNPF5.5, a nitrate transporter affecting nitrogen accumulation in Arabidopsis embryo. Sci. Rep. 2015, 5, 7962. [Google Scholar] [CrossRef] [Green Version]
- He, Y.N.; Peng, J.S.; Cai, Y.; Liu, D.F.; Guan, Y.; Yi, H.Y.; Gong, J.M. Tonoplast-localized nitrate uptake transporters involved in vacuolar nitrate efflux and reallocation in Arabidopsis. Sci. Rep. 2017, 7, 6417. [Google Scholar] [CrossRef]
- Chiu, C.C.; Lin, C.S.; Hsia, A.P.; Su, R.C.; Lin, H.L.; Tsay, Y.F. Mutation of a nitrate transporter, AtNRT1:4, results in a reduced petiole nitrate content and altered leaf development. Plant Cell Physiol. 2004, 45, 1139–1148. [Google Scholar] [CrossRef]
- Krouk, G.; Lacombe, B.; Bielach, A.; Perrine-Walker, F.; Malinska, K.; Mounier, E.; Hoyerova, K.; Tillard, P.; Leon, S.; Ljung, K.; et al. Nitrate-regulated auxin transport by NRT1.1 defines a mechanism for nutrient sensing in plants. Dev. Cell 2010, 18, 927–937. [Google Scholar] [CrossRef]
- Li, J.Y.; Fu, Y.L.; Pike, S.M.; Bao, J.; Tian, W.; Zhang, Y.; Chen, C.Z.; Zhang, Y.; Li, H.M.; Huang, J.; et al. The Arabidopsis nitrate transporter NRT1.8 functions in nitrate removal from the xylem sap and mediates cadmium tolerance. Plant Cell 2010, 22, 1633–1646. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.H.; Kuo, H.F.; Canivenc, G.; Lin, C.S.; Lepetit, M.; Hsu, P.K.; Tillard, P.; Lin, H.L.; Wang, Y.Y.; Tsai, C.B.; et al. Mutation of the Arabidopsis NRT1.5 nitrate transporter causes defective root-to-shoot nitrate transport. Plant Cell 2008, 20, 2514–2528. [Google Scholar] [CrossRef] [Green Version]
- Komarova, N.Y.; Thor, K.; Gubler, A.; Meier, S.; Dietrich, D.; Weichert, A.; Grotemeyer, M.S.; Tegeder, M.; Rentsch, D. AtPTR1 and AtPTR5 transport dipeptides in planta. Plant Physiol. 2008, 148, 856–869. [Google Scholar] [CrossRef]
- Chiba, Y.; Shimizu, T.; Miyakawa, S.; Kanno, Y.; Koshiba, T.; Kamiya, Y.; Seo, M. Identification of Arabidopsis thaliana NRT1/PTR FAMILY (NPF) proteins capable of transporting plant hormones. J. Plant Res. 2015, 128, 679–686. [Google Scholar] [CrossRef]
- Huang, X.; Wang, B.; Xi, J.; Zhang, Y.; He, C.; Zheng, J.; Gao, J.; Chen, H.; Zhang, S.; Wu, W.; et al. Transcriptome comparison reveals distinct selection patterns in domesticated and wild Agave species, the important CAM plants. Int. J. Genom. 2018, 2018, 5716518. [Google Scholar] [CrossRef] [Green Version]
- Beltran-Garcia, M.J.; White, J.F., Jr.; Prado, F.M.; Prieto, K.R.; Yamaguchi, L.F.; Torres, M.S.; Kato, M.J.; Medeiros, M.H.; Di Mascio, P. Nitrogen acquisition in Agave tequilana from degradation of endophytic bacteria. Sci. Rep. 2014, 4, 6938. [Google Scholar] [CrossRef]
- Yang, X.; Cushman, J.C.; Borland, A.M.; Edwards, E.J.; Wullschleger, S.D.; Tuskan, G.A.; Owen, N.A.; Griffiths, H.; Smith, J.A.; De Paoli, H.C.; et al. A roadmap for research on crassulacean acid metabolism (CAM) to enhance sustainable food and bioenergy production in a hotter, drier world. New Phytol. 2015, 207, 491–504. [Google Scholar] [CrossRef]
- Nava-Cruz, N.Y.; Medina-Morales, M.A.; Martinez, J.L.; Rodriguez, R.; Aguilar, C.N. Agave biotechnology: An overview. Crit. Rev. Biotechnol. 2015, 35, 546–559. [Google Scholar] [CrossRef]
- Deng, G.; Huang, X.; Xie, L.; Tan, S.; Gbokie, T., Jr.; Bao, Y.; Xie, Z.; Yi, K. Identification and Expression of SAUR Genes in the CAM Plant Agave. Genes 2019, 10, 555. [Google Scholar] [CrossRef] [Green Version]
- Robert, M.L.; Lim, K.Y.; Hanson, L.; Sanchez-Teyer, F.; Bennett, M.D.; Leitch, A.R.; Leitch, I.J. Wild and agronomically important Agave species (Asparagaceae) show proportional increases in chromosome number, genome size, and genetic markers with increasing ploidy. Bot. J. Linn. Soc. 2010, 158, 215–222. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Xiao, M.; Xi, J.; He, C.; Zheng, J.; Chen, H.; Gao, J.; Zhang, S.; Wu, W.; Liang, Y.; et al. De novo transcriptome assembly of Agave H11648 by Illumina sequencing and identification of cellulose synthase genes in Agave species. Genes 2019, 10, 103. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.B.; Meng, S.; Gong, J.M. The Expected and Unexpected Roles of Nitrate Transporters in Plant Abiotic Stress Resistance and Their Regulation. Int. J. Mol. Sci. 2018, 19, 3535. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Li, S.; Shi, M.; Wang, S.; Shi, L.; Xu, F.; Ding, G. Genome-Wide Systematic Characterization of the NPF Family Genes and Their Transcriptional Responses to Multiple Nutrient Stresses in Allotetraploid Rapeseed. Int. J. Mol. Sci. 2020, 21, 5947. [Google Scholar] [CrossRef]
- Harkess, A.; Zhou, J.; Xu, C.; Bowers, J.E.; Van der Hulst, R.; Ayyampalayam, S.; Mercati, F.; Riccardi, P.; McKain, M.R.; Kakrana, A.; et al. The asparagus genome sheds light on the origin and evolution of a young Y chromosome. Nat. Commun. 2017, 8, 1279. [Google Scholar] [CrossRef]
- Wen, J.; Li, P.F.; Ran, F.; Guo, P.C.; Zhu, J.T.; Yang, J.; Zhang, L.L.; Chen, P.; Li, J.N.; Du, H. Genome-wide characterization, expression analyses, and functional prediction of the NPF family in Brassica napus. BMC Genom. 2020, 21, 871. [Google Scholar] [CrossRef]
- Zolfaghari, F.; Khosravi, H.; Shahriyari, A.; Jabbari, M.; Abolhasani, A. Hierarchical cluster analysis to identify the homogeneous desertification management units. PLoS ONE 2019, 14, e0226355. [Google Scholar] [CrossRef]
- Schuster, S.C. Next-generation sequencing transforms today’s biology. Nat. Methods 2008, 5, 16–18. [Google Scholar] [CrossRef]
- Mahmoud, M.; Zywicki, M.; Twardowski, T.; Karlowski, W.M. Efficiency of PacBio long read correction by 2nd generation Illumina sequencing. Genom. 2019, 111, 43–49. [Google Scholar] [CrossRef]
- Wang, P.; Kelly, S.; Fouracre, J.P.; Langdale, J.A. Genome-wide transcript analysis of early maize leaf development reveals gene cohorts associated with the differentiation of C4 Kranz anatomy. Plant J. 2013, 75, 656–670. [Google Scholar] [CrossRef]
- Hunter, N. Meiotic Recombination: The Essence of Heredity. Cold Spring Harb. Perspect. Biol. 2015, 7, a016618. [Google Scholar] [CrossRef] [Green Version]
- Zhao, B.; Cao, J.F.; Hu, G.J.; Chen, Z.W.; Wang, L.Y.; Shangguan, X.X.; Wang, L.J.; Mao, Y.B.; Zhang, T.Z.; Wendel, J.F.; et al. Core cis-element variation confers subgenome-biased expression of a transcription factor that functions in cotton fiber elongation. New Phytol. 2018, 218, 1061–1075. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Hu, R.; Palla, K.J.; Tuskan, G.A.; Yang, X. Advances and perspectives on the use of CRISPR/Cas9 systems in plant genomics research. Curr. Opin. Plant Biol. 2016, 30, 70–77. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.; Takahashi, H. Improving nitrogen use efficiency: From cells to plant systems. J. Exp. Bot. 2020, 71, 4359–4364. [Google Scholar] [CrossRef]
- Zhu, J.K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [Green Version]
- Nath, M.; Tuteja, N. NPKS uptake, sensing, and signaling and miRNAs in plant nutrient stress. Protoplasma 2016, 253, 767–786. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Rombel, I.T.; Sykes, K.F.; Rayner, S.; Johnston, S.A. ORF-FINDER: A vector for high-throughput gene identification. Gene 2002, 282, 33–41. [Google Scholar] [CrossRef]
- Yu, C.S.; Chen, Y.C.; Lu, C.H.; Hwang, J.K. Prediction of protein subcellular localization. Proteins 2006, 64, 643–651. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Hoagland, D.R.; Arnon, D.S. The water culture method for growing plants without soil. Calif. Agric. Exp. Stn. Circ. 1950, 347, 32. [Google Scholar]
- Deng, G.; Liu, L.J.; Zhong, X.Y.; Lao, C.Y.; Wang, H.Y.; Wang, B.; Zhu, C.; Shah, F.; Peng, D.X. Comparative proteome analysis of the response of ramie under N, P and K deficiency. Planta 2014, 239, 1175–1186. [Google Scholar] [CrossRef]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Accession | Coding Sequence (bp) | Predicted Protein (aa) | Molecular Weight | pI | Subcellular Localization |
|---|---|---|---|---|---|---|
| AhNPF2.1 | DN41135_c0_g1_i1 | 1767 | 588 | 65,558.83 | 8.91 | PlasmaMembrane (4.946) |
| AhNPF2.2 | DN48816_c0_g1_i2 | 2400 | 799 | 89,795.73 | 9.29 | PlasmaMembrane (3.655) |
| AhNPF2.3 | DN50815_c0_g2_i3 | 1866 | 621 | 68,836.88 | 8.57 | PlasmaMembrane (4.952) |
| AhNPF3.1 | DN41985_c0_g1_i1 | 1797 | 598 | 66,083.81 | 8.58 | PlasmaMembrane (4.828) |
| AhNPF3.2 | DN49643_c0_g1_i2 | 1788 | 595 | 65,163.86 | 9.11 | PlasmaMembrane (4.727) |
| AhNPF4.1 | DN35785_c0_g1_i1 | 1761 | 586 | 64,956.66 | 8.90 | PlasmaMembrane (4.906) |
| AhNPF4.2 | DN44495_c0_g1_i3 | 1743 | 580 | 63,224.67 | 8.98 | PlasmaMembrane (4.860) |
| AhNPF4.3 | DN46864_c0_g1_i1 | 1782 | 593 | 66,186.76 | 8.21 | PlasmaMembrane (4.872) |
| AhNPF5.1 | DN41592_c0_g1_i2 | 1668 | 555 | 61,626.22 | 5.80 | PlasmaMembrane (4.911) |
| AhNPF5.2 | DN44315_c0_g1_i1 | 1611 | 536 | 59,762.91 | 6.47 | PlasmaMembrane (4.969) |
| AhNPF5.3 | DN46789_c0_g1_i1 | 1602 | 533 | 59,330.15 | 8.77 | PlasmaMembrane (4.937) |
| AhNPF5.4 | DN47034_c0_g2_i1 | 1734 | 577 | 63,963.40 | 8.70 | PlasmaMembrane (4.971) |
| AhNPF5.5 | DN48393_c3_g3_i2 | 1761 | 586 | 65,431.38 | 9.05 | PlasmaMembrane (4.836) |
| AhNPF6.1 | DN42911_c0_g2_i1 | 1737 | 578 | 62,801.28 | 9.00 | PlasmaMembrane (4.919) |
| AhNPF7.1 | DN45587_c0_g2_i1 | 1854 | 617 | 68,530.90 | 5.77 | PlasmaMembrane (4.917) |
| AhNPF8.1 | DN41583_c0_g1_i1 | 1377 | 458 | 50,193.55 | 6.82 | PlasmaMembrane (4.782) |
| AhNPF8.2 | DN43232_c0_g2_i2 | 1773 | 590 | 65,292.70 | 5.87 | PlasmaMembrane (4.876) |
| AhNPF8.3 | DN50692_c0_g1_i8 | 1701 | 566 | 62,878.70 | 7.09 | PlasmaMembrane (4.754) |
| AhNPF8.4 | DN51386_c0_g1_i1 | 1758 | 585 | 64,615.74 | 6.18 | PlasmaMembrane (4.838) |
| ID | Forward Primer | Reverse Primer | Product Size (bp) |
|---|---|---|---|
| AhNPF2.1 | GCGCAGACCAATTCAATCCT | CGTGCCGATGAAGAAGAAGG | 206 |
| AhNPF2.2 | TCTCCTTCTTCCAAGCCCTG | AGCTCACCTGATAGACACCG | 160 |
| AhNPF2.3 | GCTTCCAAATTCCTCCTGCC | CACAATCATGGACAGAGCCG | 182 |
| AhNPF3.1 | CCTCGTTGCTGGATTCATCG | CTCCCCGCTGATATCACCAT | 240 |
| AhNPF3.2 | TGCTCTAATCGCCGACTCAT | ACCAATGCAAGTCAGGAGGA | 208 |
| AhNPF4.1 | ATCTGCCCGTCAAACTCTGA | CTTGCTCAACCTGTGTCACC | 181 |
| AhNPF4.2 | TCATCCCTGAAGCCTCCATC | GTGTCTTCGTCGAACTGCTC | 173 |
| AhNPF4.3 | TGGTGGAGAGCAAGAGAAGG | ACCCGAATGAGTACGAGCAA | 201 |
| AhNPF5.1 | ATCCTCCATTTTGCCAAGCG | GACCAGAACCGAGACCATCA | 223 |
| AhNPF5.2 | TGGCTGGTGTGGAGAGATTT | CAAGTCCCGCAACATAGAGC | 222 |
| AhNPF5.3 | ACCAACACCTGCGACAAATC | CTCCACTACAGAGGCCACAA | 199 |
| AhNPF5.4 | ATTGCGAGCGAGACCAAAAG | TCTCAGCAACCTCAACACCT | 187 |
| AhNPF5.5 | AGATAACGTGGGGTGGACTG | AGCTCCTTGGGGTCATTAGG | 195 |
| AhNPF6.1 | CAATTCGACGACAAGGACCC | AGGACCATCGATATGCAGCA | 170 |
| AhNPF7.1 | ACCTGGGCTCTCTCTTTTCC | TTGACCGTCCATTTCCTCGA | 214 |
| AhNPF8.1 | ATCAAACGCCTCCAGATTGC | CGTCGTCACAAGAGCAAGAG | 204 |
| AhNPF8.2 | GCAAGATAACTGCGGATGGG | AGAGAACTGTCAAGAGGGGC | 195 |
| AhNPF8.3 | GCATAATCTTCGCCACGGTT | CTCATGTCCGGTGAACTTGC | 200 |
| AhNPF8.4 | CCGCAGTCTACAGCCAAATC | AAGCCTCTCTCTTTGCCAGT | 193 |
| PP2A | CCTCCTCCTCCTTCGGTTTG | GCCATGAATGTCACCGCAGA | 235 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, S.; Liang, Y.; Huang, Y.; Xi, J.; Huang, X.; Yang, X.; Yi, K. Phylogeny and Expression Atlas of the NITRATE TRANSPORTER 1/PEPTIDE TRANSPORTER FAMILY in Agave. Plants 2022, 11, 1434. https://doi.org/10.3390/plants11111434
Tan S, Liang Y, Huang Y, Xi J, Huang X, Yang X, Yi K. Phylogeny and Expression Atlas of the NITRATE TRANSPORTER 1/PEPTIDE TRANSPORTER FAMILY in Agave. Plants. 2022; 11(11):1434. https://doi.org/10.3390/plants11111434
Chicago/Turabian StyleTan, Shibei, Yanqiong Liang, Yanlei Huang, Jingen Xi, Xing Huang, Xiaohan Yang, and Kexian Yi. 2022. "Phylogeny and Expression Atlas of the NITRATE TRANSPORTER 1/PEPTIDE TRANSPORTER FAMILY in Agave" Plants 11, no. 11: 1434. https://doi.org/10.3390/plants11111434
APA StyleTan, S., Liang, Y., Huang, Y., Xi, J., Huang, X., Yang, X., & Yi, K. (2022). Phylogeny and Expression Atlas of the NITRATE TRANSPORTER 1/PEPTIDE TRANSPORTER FAMILY in Agave. Plants, 11(11), 1434. https://doi.org/10.3390/plants11111434