
Biomass Production and Predicted Ethanol Yield Are Linked with Optimum Photosynthesis in Phragmites karka under Salinity and Drought Conditions

 ,
, 
 , , , ,
, , , ,  ,
,  ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Plant Growth Conditions
2.2. Plant Harvest and Growth Parameters
2.3. Soil Water Potential
2.4. Leaf Gas Exchange and Chlorophyll Content
2.5. Leaf Chlorophyll Fluorescence
2.6. Lignocellulosic Analysis and Soluble Sugar Content
2.7. Statistical Analysis and Calculation
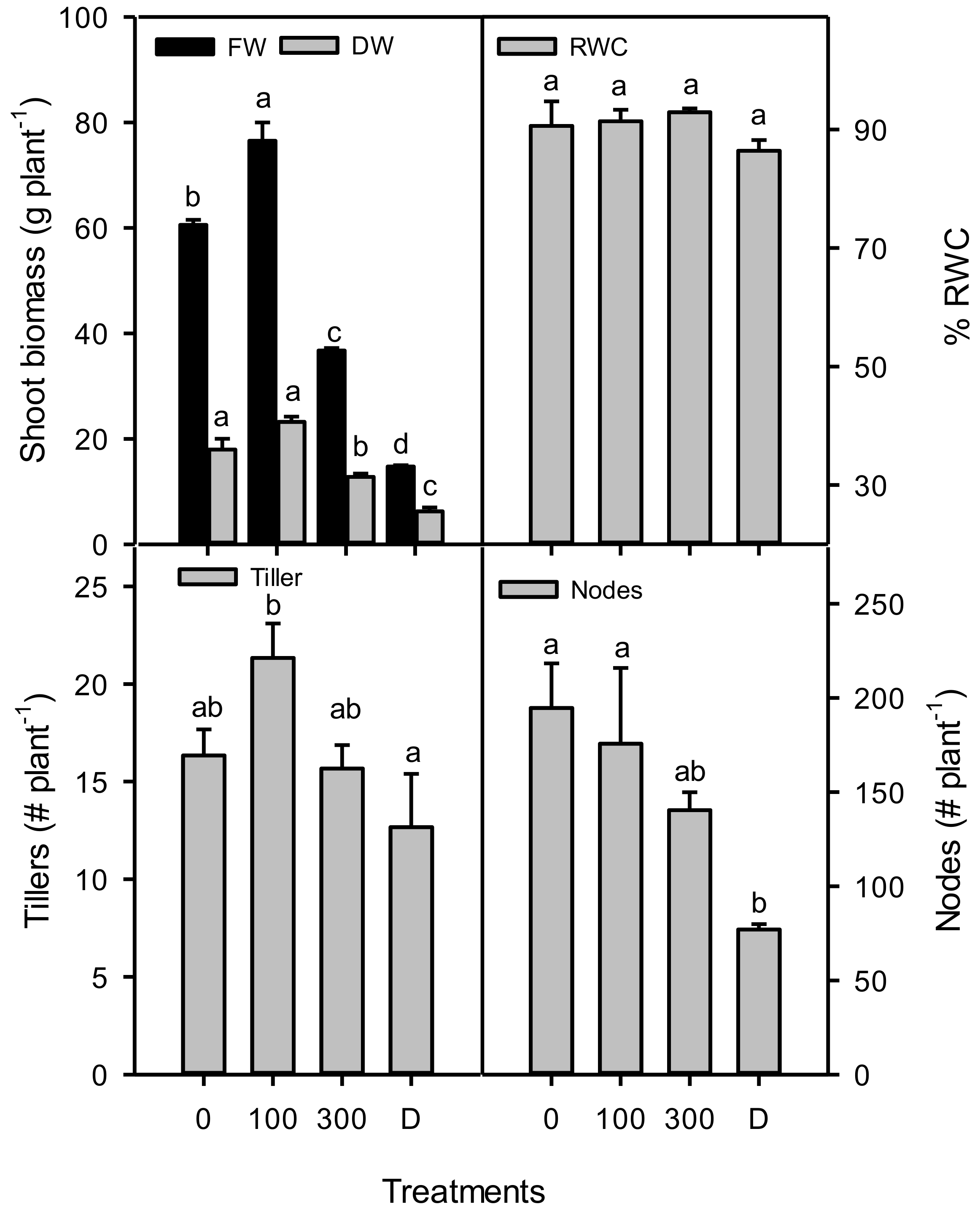
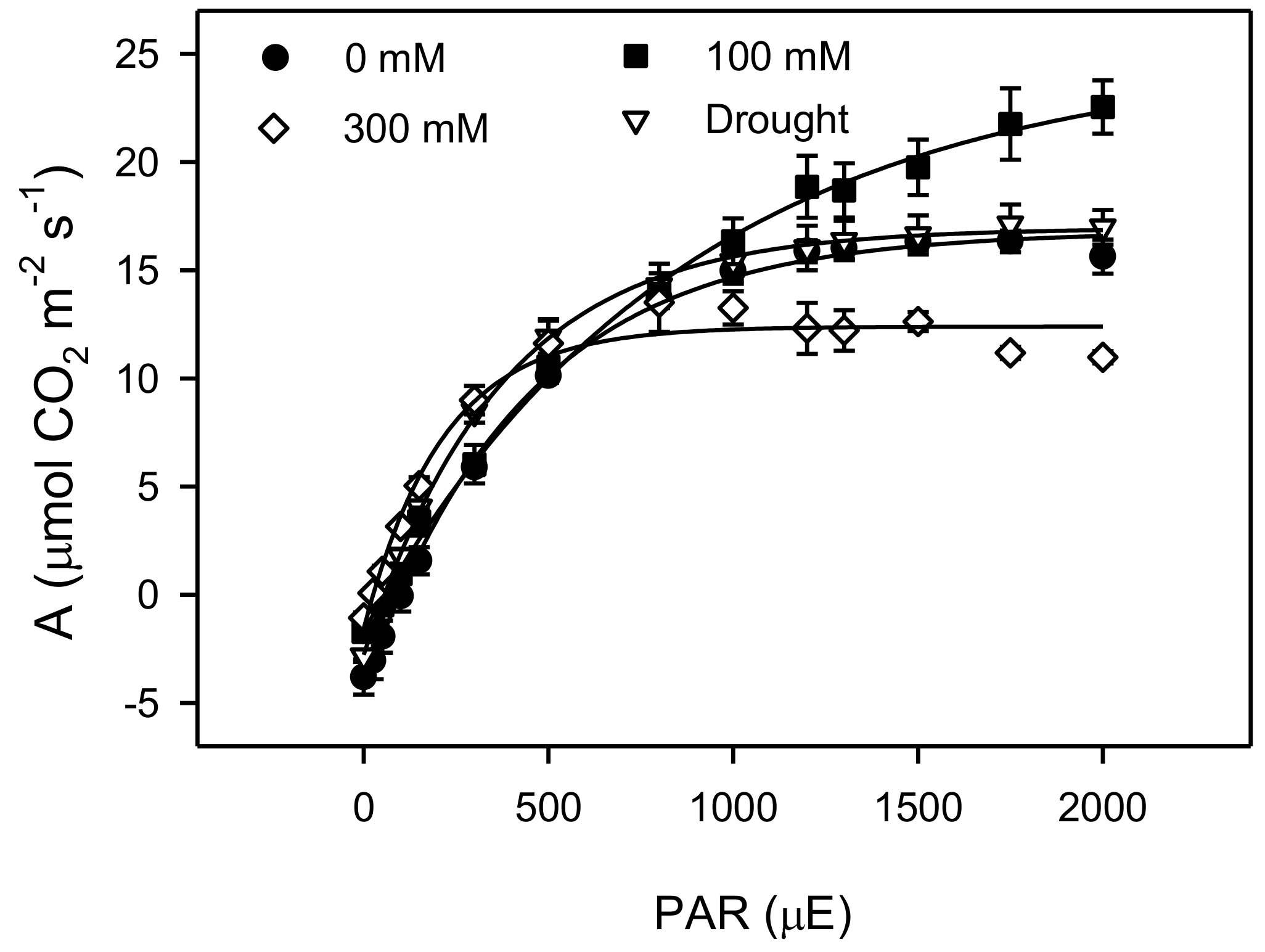
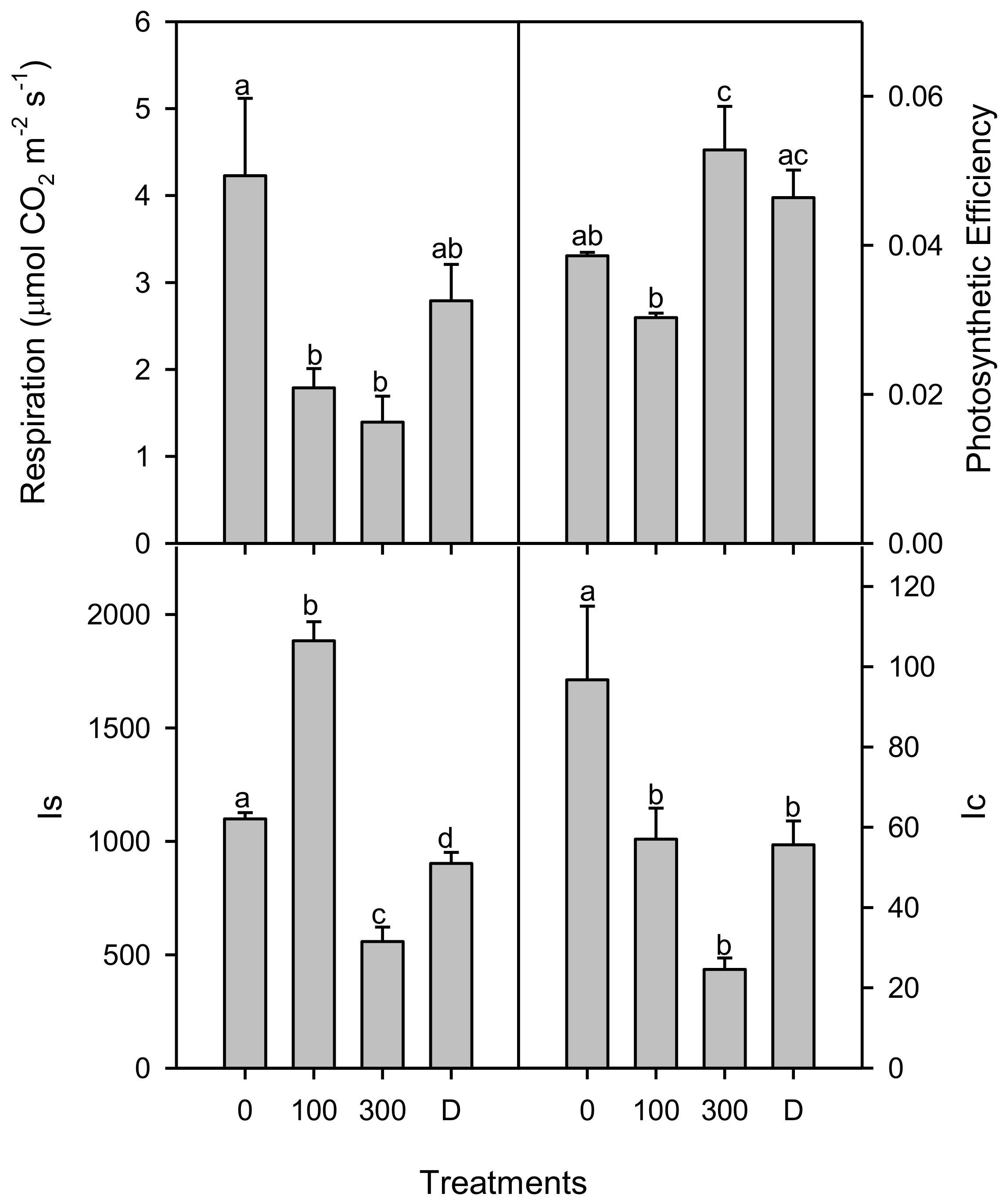
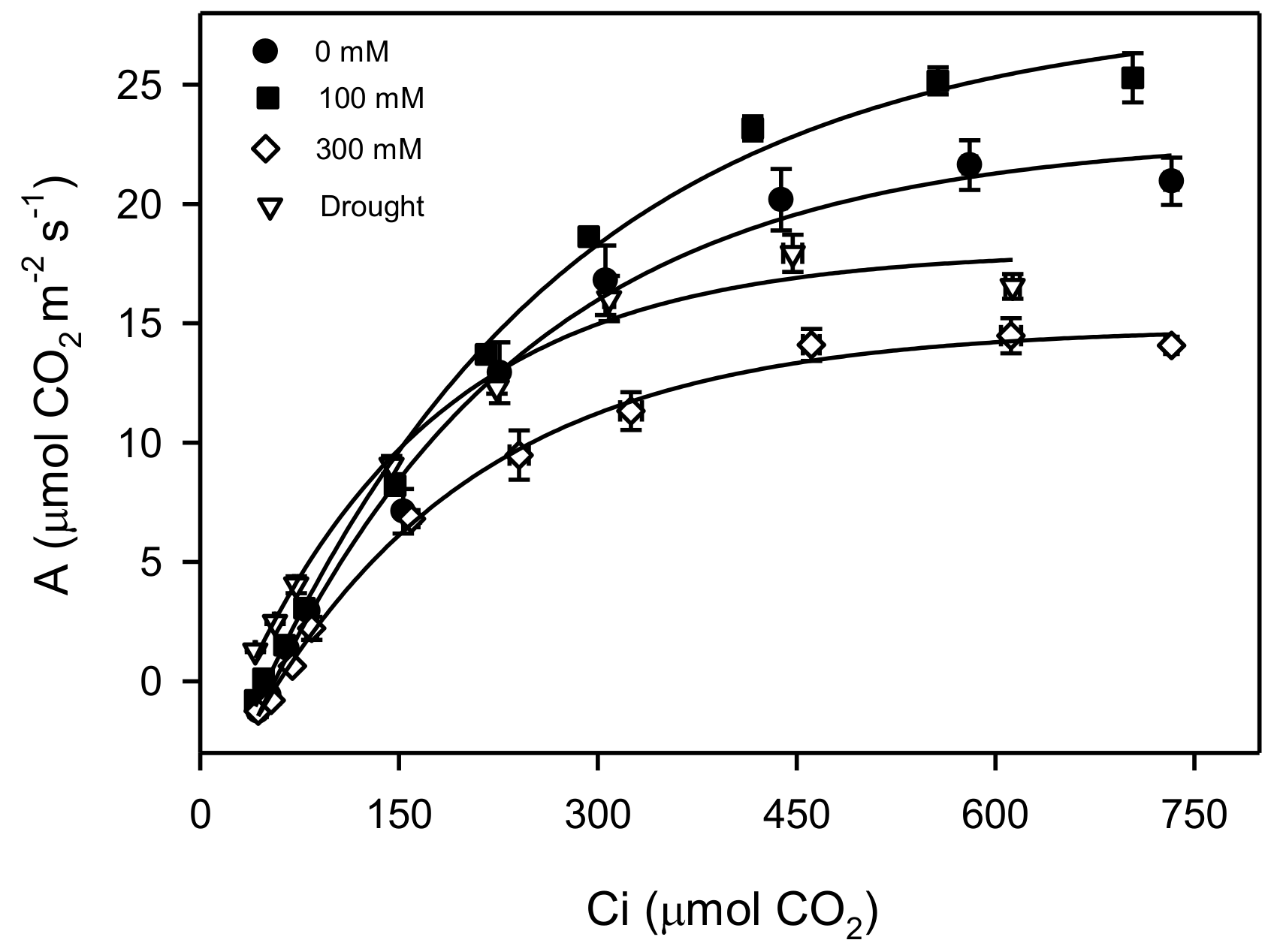
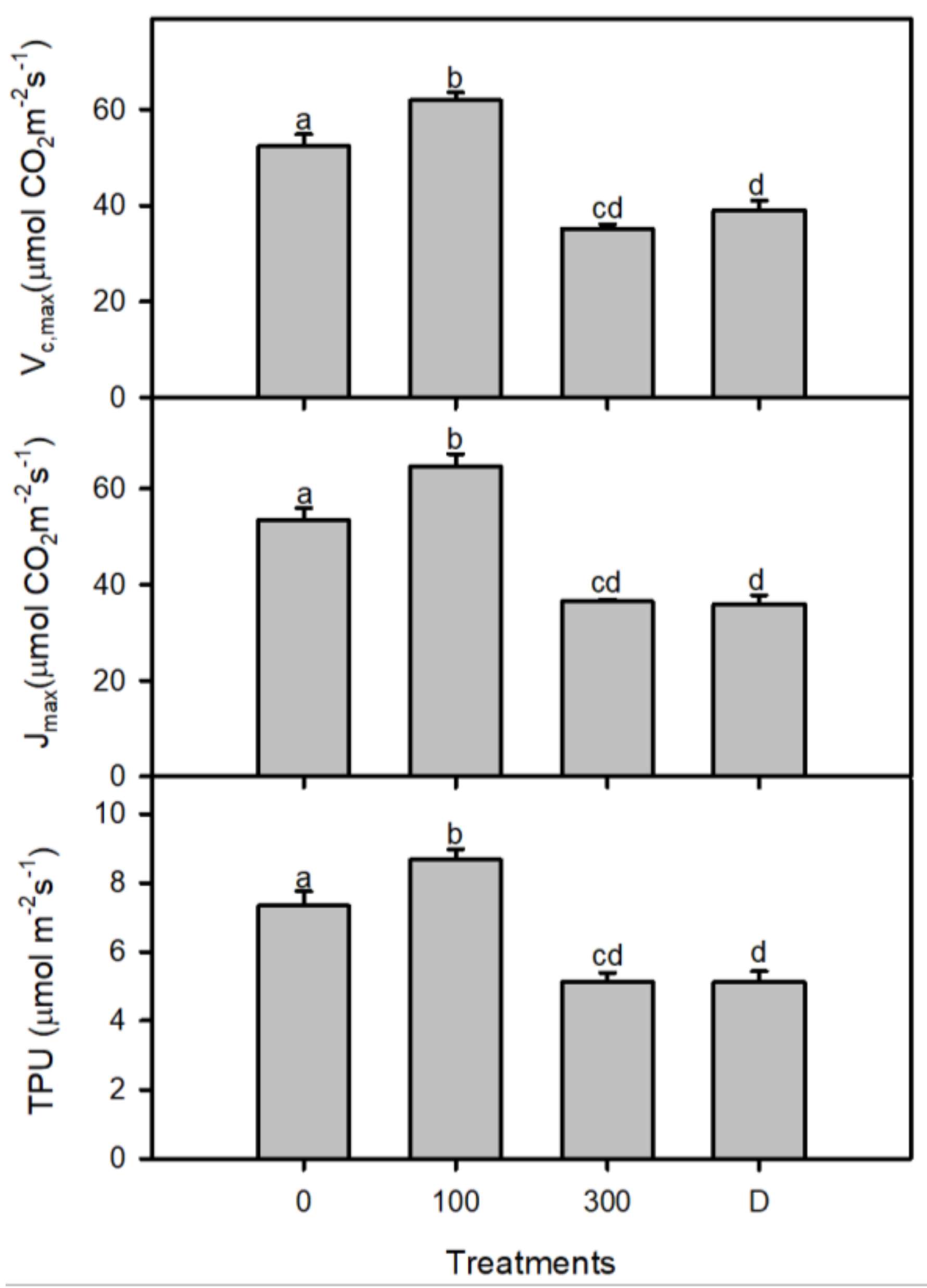
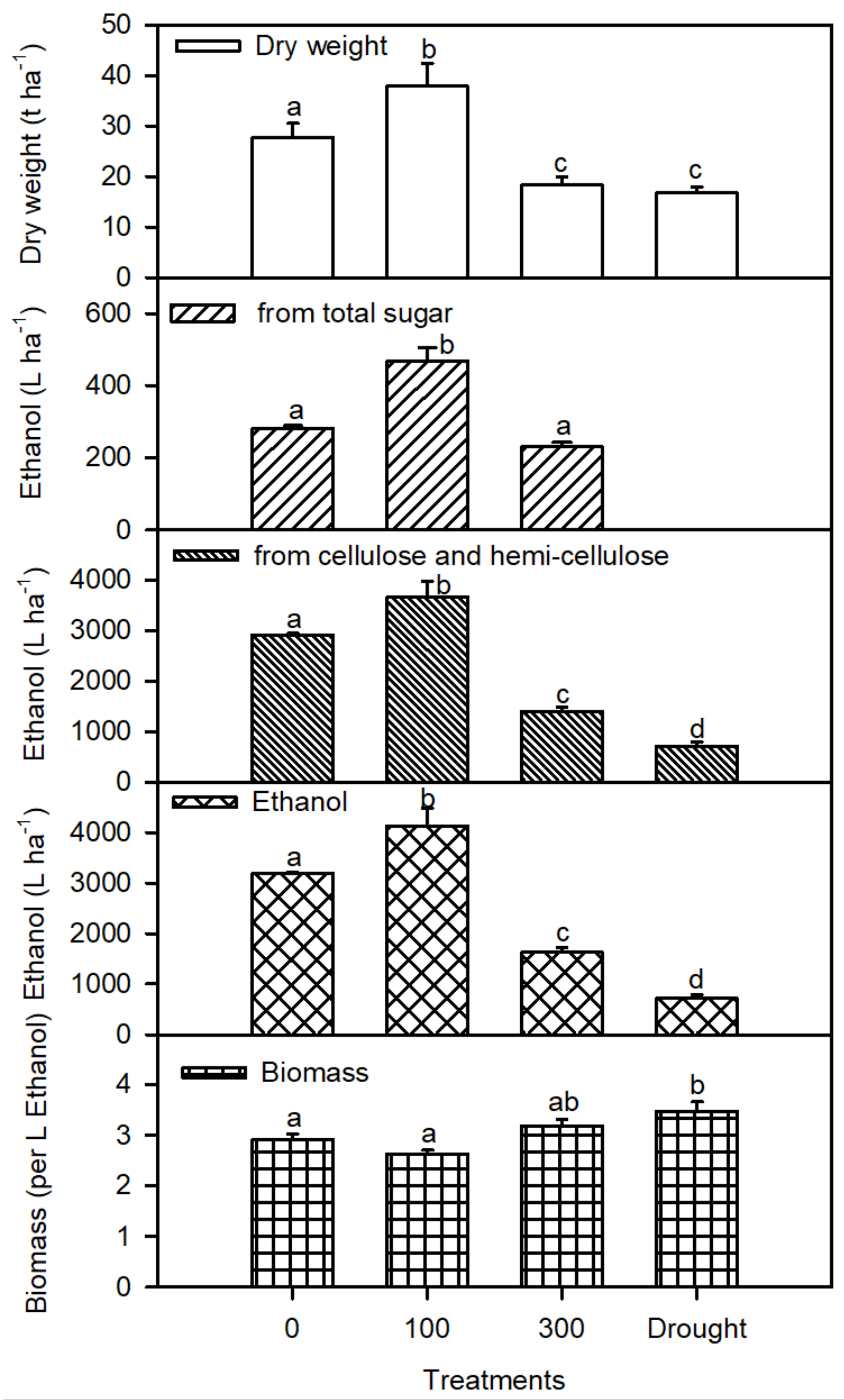
3. Results
4. Discussions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hossain, A.; Pamanick, B.; Venugopalan, V.K.; Ibrahimova, U.; Rahman, M.A.; Siyal, A.L.; Maitra, S.; Chatterjee, S.; Aftab, T. Emerging roles of plant growth regulators for plants adaptation to abiotic stress–induced oxidative stress. In Emerging Plant Growth Regulators in Agriculture; Academic Press: Cambridge, MA, USA, 2022; pp. 1–72. [Google Scholar]
- Otie, V.; Udo, I.; Shao, Y.; Itam, M.O.; Okamoto, H.; An, P.; Eneji, E.A. Salinity effects on morpho-physiological and yield traits of soybean (Glycine max L.) as mediated by foliar spray with brassinolide. Plants 2021, 10, 541. [Google Scholar] [CrossRef]
- Abideen, Z.; Koyro, H.W.; Huchzermeyer, B.; Ahmed, M.; Zulfiqar, F.; Egan, T.; Khan, M.A. Phragmites karka plants adopt different strategies to regulate photosynthesis and ion flux in saline and water deficit conditions. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2021, 155, 524–534. [Google Scholar] [CrossRef]
- Ziogas, V.; Tanou, G.; Morianou, G.; Kourgialas, N. Drought and salinity in citriculture: Optimal practices to alleviate salinity and water stress. Agronomy 2021, 11, 1283. [Google Scholar] [CrossRef]
- Kumar, A.; Sharma, S.K.; Lata, C.; Devi, R.; Kulshrestha, N.; Krishnamurthy, S.L.; Singh, K.; Yadav, R.K. Impact of water deficit (salt and drought) stress on physiological, biochemical and yield attributes on wheat (Triticum aestivum) varieties. Indian J. Agric. Sci. 2018, 88, 1624–1632. [Google Scholar]
- Álvarez, S.; Rodríguez, P.; Broetto, F.; Sánchez-Blanco, M.J. Long term responses and adaptive strategies of Pistacia lentiscus under moderate and severe deficit irrigation and salinity: Osmotic and elastic adjustment, growth, ion uptake and photosynthetic activity. Agric. Water Manag. 2018, 202, 253–262. [Google Scholar] [CrossRef] [Green Version]
- Shahzad, B.; Fahad, S.; Tanveer, M.; Saud, S.; Khan, I.A. Plant responses and tolerance to salt stress. In Approaches for Enhancing Abiotic Stress Tolerance in Plants; CRC Press: Boca Raton, FL, USA, 2019; pp. 61–78. [Google Scholar]
- Tang, L.; Zhou, Q.S.; Gao, Y.; Li, P. Biomass allocation in response to salinity and competition in native and invasive species. Ecosphere 2022, 13, e3900. [Google Scholar] [CrossRef]
- Ma, Y.; Wei, Z.; Liu, J.; Liu, X.; Liu, F. Growth and physiological responses of cotton plants to salt stress. J. Agron. Crop Sci. 2021, 207, 565–576. [Google Scholar] [CrossRef]
- Belghith, I.; Senkler, J.; Abdelly, C.; Braun, H.P.; Debez, A.; Turkan, I. Changes in leaf ecophysiological traits and proteome profile provide new insights into variability of salt response in the succulent halophyte Cakile maritima. Funct. Plant Biol. 2022, 49, 613–624. [Google Scholar] [CrossRef]
- Munir, N.; Hasnain, M.; Roessner, U.; Abideen, Z. Strategies in improving plant salinity resistance and use of salinity resistant plants for economic sustainability. Crit. Rev. Environ. Sci. Technol. 2021, 52, 2150–2196. [Google Scholar] [CrossRef]
- Ali, M.; Mustafa, A.; Abideen, Z.; Gul, B. Bioenergy Production from Halophytes Crops for Sustainable Development. In Energy and Environmental Security in Developing Countries; Springer: Cham, Switzerland, 2021; pp. 571–586. [Google Scholar]
- Derbali, W.; Manaa, A.; Spengler, B.; Goussi, R.; Abideen, Z.; Ghezellou, P.; Abdelly, C.; Forreiter, C.; Koyro, H.W. Comparative proteomic approach to study the salinity effect on the growth of two contrasting quinoa genotypes. Plant Physiol. Biochem. 2021, 163, 215–229. [Google Scholar] [CrossRef]
- Kronzucker, H.J.; Coskun, D.; Schulze, L.M.; Wong, J.R.; Britto, D.T. Sodium as nutrient and toxicant. Plant Soil 2013, 369, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Hussain, T.; Koyro, H.W.; Zhang, W.; Liu, X.; Gul, B.; Liu, X. Low salinity improves photosynthetic performance in Panicum antidotale under drought stress. Front. Plant Sci. 2020, 11, 481. [Google Scholar] [CrossRef]
- Marshner, W.H. Aquinas on the evaluation of human actions. Thomist A Specul. Q. Rev. 1995, 59, 347–370. [Google Scholar] [CrossRef]
- Subbarao, G.V.; Ito, O.; Berry, W.L.; Wheeler, R.M. Sodium—A functional plant nutrient. Crit. Rev. Plant Sci. 2003, 22, 391–416. [Google Scholar]
- Grof, C.P.; Johnston, M.; Brownell, P.F. Effect of sodium nutrition on the ultrastructure of chloroplasts of C4 plants. Plant Physiol. 1989, 89, 539–543. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Peñalver, A.; Reigosa, M.J.; Sánchez-Moreiras, A.M. Imaging chlorophyll a fluorescence reveals specific spatial distributions under different stress conditions. Flora-Morphol. Distrib. Funct. Ecol. Plants 2011, 206, 836–844. [Google Scholar] [CrossRef]
- Ghorbanzadeh, P.; Aliniaeifard, S.; Esmaeili, M.; Mashal, M.; Azadegan, B.; Seif, M. Dependency of growth, water use efficiency, chlorophyll fluorescence, and stomatal characteristics of lettuce plants to light intensity. J. Plant Growth Regul. 2021, 40, 2191–2207. [Google Scholar] [CrossRef]
- Abideen, Z.; Ansari, R.; Khan, M.A. Halophytes: Potential source of ligno-cellulosic biomass for ethanol production. Biomass Bioenergy 2011, 35, 1818–1822. [Google Scholar] [CrossRef]
- Abideen, Z.; Qasim, M.; Hussain, T.; Rasheed, A.; Gul, B.; Koyro, H.W.; Ansari, R.; Khan, M.A. Salinity improves growth, photosynthesis and bioenergy characteristics of Phragmites karka. Crop Pasture Sci. 2018, 69, 944–953. [Google Scholar] [CrossRef]
- Epstein, E. Mineral Nutrition of Plants: Principles and Perspectives; John Wiley and Sons, Inc.: New York, NY, USA, 1972. [Google Scholar]
- Koyro, H.W. Study of potential cash crop halophytes by a quick check system: Determination of the threshold of salinity tolerance and the ecophysiological demands. In Cash Crop Halophytes: Recent Studies; Springer: Dordrecht, The Netherlands, 2003; pp. 5–17. [Google Scholar]
- Veihmeyer, F.J.; Hendrickson, A.H. The moisture equivalent as a measure of the field capacity of soils. Soil Sci. 1931, 32, 181–194. [Google Scholar] [CrossRef]
- Schulte, M.; Offer, C.; Hansen, U. Induction of CO2-gas exchange and electron transport: Comparison of dynamic and steady-state responses in Fagus sylvatica leaves. Trees 2003, 17, 153–163. [Google Scholar] [CrossRef]
- Long, S.P.; Bernacchi, C.J. Gas exchange measurements, what can they tell us about the underlying limitations to photosynthesis? Procedures and sources of error. J. Exp. Bot. 2003, 54, 2393–2401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abideen, Z.; Koyro, H.W.; Huchzermeyer, B.; Ansari, R.; Zulfiqar, F.; Gul, B.J.P.B. Ameliorating effects of biochar on photosynthetic efficiency and antioxidant defence of Phragmites karka under drought stress. Plant Biol. 2020, 22, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Kitajima, M.B.W.L.; Butler, W.L. Quenching of chlorophyll fluorescence and primary photochemistry in chloroplasts by dibromothymoquinone. Biochim. Et Biophys. Acta (BBA)-Bioenerg. 1975, 376, 105–115. [Google Scholar] [CrossRef]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Et Biophys. Acta (BBA)-Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Bilger, W.; Björkman, O. Role of the xanthophyll cycle in photoprotection elucidated by measurements of light-induced absorbance changes, fluorescence and photosynthesis in leaves of Hedera canariensis. Photosynth. Res. 1990, 25, 173–185. [Google Scholar] [CrossRef]
- Van Kooten, O.; Snel, J.F. The use of chlorophyll fluorescence nomenclature in plant stress physiology. Photosynth. Res. 1990, 25, 147–150. [Google Scholar] [CrossRef]
- Krall, J.P.; Edwards, G.E. Relationship between photosystem II activity and CO2 fixation in leaves. Physiol. Plant. 1992, 86, 180–187. [Google Scholar] [CrossRef]
- Sun, Y.; Cheng, J. Hydrolysis of lignocellulosic materials for ethanol production: A review. Bioresour. Technol. 2002, 83, 1–11. [Google Scholar] [CrossRef]
- Ludwig, T.G.; Goldberg, H.J. The anthrone method for the determination of carbohydrates in foods and in oral rinsing. J. Dent. Res. 1956, 35, 90–94. [Google Scholar] [CrossRef]
- Zhao, Y.L.; Dolat, A.; Steinberger, Y.; Wang, X.; Osman, A.; Xie, G.H. Biomass yield and changes in chemical composition of sweet sorghum cultivars grown for biofuel. Field Crops Res. 2009, 111, 55–64. [Google Scholar] [CrossRef]
- Abideen, Z.; Ansari, R.; Gul, B.; Khan, M.A. The place of halophytes in Pakistan’s biofuel industry. Biofuels 2012, 3, 211–220. [Google Scholar] [CrossRef]
- Gul, B.; Abideen, Z.; Ansari, R.; Khan, M.A. Halophytic biofuels revisited. Biofuels 2013, 4, 575–577. [Google Scholar] [CrossRef]
- Khan, M.A.; Ansari, R.; Ali, H.; Gul, B.; Nielsen, B.L. Panicum turgidum a potentially sustainable cattle feed alternative to maize for saline areas. Agric. Ecosyst. Environ. 2009, 129, 542–546. [Google Scholar] [CrossRef]
- Abideen, Z.; Koyro, H.W.; Huchzermeyer, B.; Ahmed, M.Z.; Gul, B.; Khan, M.A. Moderate salinity stimulates growth and photosynthesis of Phragmites karka by water relations and tissue specific ion regulation. Environ. Exp. Bot. 2014, 105, 70–76. [Google Scholar] [CrossRef]
- Gorai, M.; Vadel, A.M.; Neffati, M.; Khemira, H. The effect of sodium chloride salinity on the growth, water status, and ion content of Phragmites communis. Pak. J. Biol. Sci. 2007, 13, 2225–2230. [Google Scholar] [CrossRef] [Green Version]
- Gorai, M.; Ennajeh, M.; Khemira, H.; Neffati, M. Combined effect of NaCl-salinity and hypoxia on growth, photosynthesis, water relations and solute accumulation in Phragmites australis plants. Flora 2010, 205, 462–470. [Google Scholar] [CrossRef]
- Naidoo, G.; Naidoo, Y.; Achar, P. Ecophysiological responses of the salt marsh grass Spartina maritima to salinity. Afr. J. Aquat. Sci. 2012, 37, 81–88. [Google Scholar] [CrossRef]
- Gulzar, S.; Khan, M.A.; Liu, X. Seed germination strategies of Desmostachya bipinnata: A fodder crop for saline soils. Rangel. Ecol. Manag. 2007, 60, 401–407. [Google Scholar] [CrossRef]
- Geissler, N.; Hussin, S.; Koyro, H.-W. Elevated atmospheric CO2 concentration ameliorates effects of NaCl salinity on photosynthesis and leaf structure of Aster tripolium L. J. Exp. Bot. 2009, 60, 137–151. [Google Scholar] [CrossRef] [Green Version]
- Redondo-Gómez, S.; Mateos-Naranjo, E.; Figueroa, M.E.; Davy, A.J. Salt stimulation of growth and photosynthesis in an extreme halophyte Arthrocnemum macrostachyum. Plant Biol. 2010, 12, 79–87. [Google Scholar] [CrossRef]
- Asrar, H.; Hussain, T.; Hadi, S.M.S.; Gul, B.; Nielsen, B.L.; Khan, M.A. Salinity induced changes in light harvesting and carbon assimilating complexes of Desmostachya bipinnata (L.) Staph. Environ. Exp. Bot. 2017, 135, 86–95. [Google Scholar] [CrossRef]
- Moinuddin, M.; Gulzar, S.; Hameed, A.; Gul, B.; Ajmal Khan, M.; Edwards, G.E. Differences in photosynthetic syndromes of four halophytic marsh grasses in Pakistan. Photosynth. Res. 2017, 131, 51–64. [Google Scholar] [CrossRef]
- Pagter, M.; Bragato, C.; Malagol, M.; Brix, H. Osmotic and ionic effects of NaCl and Na2SO4 salinity on Phragmites australis. Aquat. Bot. 2009, 90, 43–51. [Google Scholar] [CrossRef]
- Koyro, H.W. Effect of salinity on growth, photosynthesis, water relations and solute composition of the potential cash crop halophyte Plantago coronopus (L.). Environ. Exp. Bot. 2006, 56, 136–146. [Google Scholar] [CrossRef]
- Pieters, A.J.; Paul, M.J.; Lawlor, D.W. Low sink demand limits photosynthesis under Pi deficiency. J. Exp. Bot. 2001, 52, 1083–1091. [Google Scholar] [CrossRef] [Green Version]
- Lin, Z.H.; Chen, L.S.; Chen, R.B.; Zhang, F.Z.; Jiang, H.X.; Tang, N. CO2assimilation, ribulose-1, 5-bisphosphate carboxylase/oxygenase, carbohydrates and photosynthetic electron transport probed by the JIP-test, of tea leaves in response to phosphorus supply. BMC Plant Biol. 2009, 9, 43. [Google Scholar] [CrossRef] [Green Version]
- Athar, H.u.R.; Ashraf, M. Photosynthesis under Drought Stress. In Handbook of Photosynthesis, 2nd ed.; Pessarakli, M., Ed.; CRC Press: Boca Raton, FL, USA, 2005; pp. 793–809. [Google Scholar]
- Bano, S.; Ahmed, M.Z.; Abideen, Z.; Qasim, M.; Gul, B.; Khan, N.U. Humic acid overcomes salinity barriers and stimulates growth of Urochondra setulosa by altering ion-flux and photochemistry. Acta Physiol. Plant. 2022, 44, 39. [Google Scholar] [CrossRef]
- Ehsen, S.; Abideen, Z.; Rizvi, R.F.; Gulzar, S.; Aziz, I.; Gul, B.; Khan, M.A.; Ansari, R. Ecophysiological adaptations and anti-nutritive status of sustainable cattle feed Haloxylon stocksii under saline conditions. Flora 2019, 257, 151425. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Wei, Y.; Xu, X.; Tao, H.; Wang, P. Growth performance and physiological response in the halophyte Lycium barbarum grown at salt-affected soil. Ann. Appl. Biol. 2006, 149, 263–269. [Google Scholar] [CrossRef]
- Qiu, N.; Lu, Q.; Lu, C. Photosynthesis, photosystem II efficiency and the xanthophyll cycle in the salt-adapted halophyte Atriplex centralasiatica. New Phytol. 2003, 159, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Pompeiano, A.; Landi, M.; Meloni, G.; Vita, F.; Guglielminetti, L.; Guidi, L. Allocation pattern, ion partitioning, and chlorophyll a fluorescence in Arundo donax L. in responses to salinity stress. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2017, 151, 613–622. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, J.; Wang, X.; Durachko, D.M.; Zhang, S.; Cosgrove, D.J. Molecular insights into the complex mechanics of plant epidermal cell walls. Science 2021, 372, 706–711. [Google Scholar] [CrossRef] [PubMed]
- Cosgrove, D.J. Growth of the plant cell wall. Nat. Rev. Mol. Cell Biol. 2005, 6, 850–861. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, X.; Yang, S.; Wang, C.; Lu, G.; Wang, R.; Yang, Y.; Li, D. Heterogenous expression of Pyrus pyrifolia PpCAD2 and PpEXP2 in tobacco impacts lignin accumulation in transgenic plants. Gene 2017, 637, 181–189. [Google Scholar] [CrossRef]
- Maksup, S.; Sengsai, S.; Laosuntisuk, K.; Asayot, J.; Pongprayoon, W. Physiological responses and the expression of cellulose and lignin associated genes in Napier grass hybrids exposed to salt stress. Acta Physiol. Plant. 2020, 42, 109. [Google Scholar] [CrossRef]
- Berndes, G.; Hoogwijk, M.; Broek, R. The contribution of biomass in the future global energy supply: A review of 17 studies. Biomass Bioenergy 2003, 25, 1–28. [Google Scholar] [CrossRef]
- Dolciotti, I.; Mambelli, S.; Grandi, S.; Venturi, G. Comparison of two sorghum genotypes for sugar and fiber production. Ind. Crops Prod. 1998, 7, 265–272. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Fv/Fm | Y(II) | qP | NPQ | Y(NO) | Y(NPQ) | ETR |
|---|---|---|---|---|---|---|---|
| 0 | 0.81 ± 0.005a | 0.51 ± 0.012a | 0.70 ± 0.015abc | 0.59 ± 0.026b | 0.30 ± 0.006a | 0.18 ± 0.008abc | 40.77 ± 0.91a |
| 100 | 0.82 ± 0.006a | 0.50 ± 0.020a | 0.68 ± 0.033b | 0.66 ± 0.093b | 0.29 ± 0.018a | 0.19 ± 0.020b | 40.22 ± 1.63a |
| 300 | 0.82 ± 0.005a | 0.51 ± 0.030a | 0.68 ± 0.032b | 0.63 ± 0.057b | 0.29 ± 0.013a | 0.18 ± 0.020b | 41.14 ± 2.45a |
| Drought | 0.81 ± 0.004a | 0.59 ± 0.014b | 0.78 ± 0.017ac | 0.47 ± 0.092a | 0.27 ± 0.015a | 0.13 ± 0.019ac | 47.12 ± 1.12b |
| Treatments | Cellulose | Hemicellulose | Soluble Sugar | Chlorophyll |
|---|---|---|---|---|
| Control | 29.17 ± 1.14b | 22.31 ± 1.11b | 51.91 ± 4.21a | 40.51 ± 0.56a |
| 100 mM NaCl | 34.56 ± 1.20c | 20.77 ± 2.00a | 78.61 ± 2.73c | 43.91 ± 0.36a |
| 300 mM NaCl | 26.67 ± 1.49b | 17.72 ± 1.56a | 79.40 ± 2.97c | 47.47 ± 0.45b |
| Drought | 20.06 ± 0.63a | 14.82 ± 0.44a | 69.50 ± 8.27b | 39.69 ± 1.99a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abideen, Z.; Koyro, H.W.; Hussain, T.; Rasheed, A.; Alwahibi, M.S.; Elshikh, M.S.; Hussain, M.I.; Zulfiqar, F.; Mansoor, S.; Abbas, Z. Biomass Production and Predicted Ethanol Yield Are Linked with Optimum Photosynthesis in Phragmites karka under Salinity and Drought Conditions. Plants 2022, 11, 1657. https://doi.org/10.3390/plants11131657
Abideen Z, Koyro HW, Hussain T, Rasheed A, Alwahibi MS, Elshikh MS, Hussain MI, Zulfiqar F, Mansoor S, Abbas Z. Biomass Production and Predicted Ethanol Yield Are Linked with Optimum Photosynthesis in Phragmites karka under Salinity and Drought Conditions. Plants. 2022; 11(13):1657. https://doi.org/10.3390/plants11131657
Chicago/Turabian StyleAbideen, Zainul, Hans Werner Koyro, Tabassum Hussain, Aysha Rasheed, Mona S. Alwahibi, Mohamed S. Elshikh, Muhammad Iftikhar Hussain, Faisal Zulfiqar, Simeen Mansoor, and Zaheer Abbas. 2022. "Biomass Production and Predicted Ethanol Yield Are Linked with Optimum Photosynthesis in Phragmites karka under Salinity and Drought Conditions" Plants 11, no. 13: 1657. https://doi.org/10.3390/plants11131657
APA StyleAbideen, Z., Koyro, H. W., Hussain, T., Rasheed, A., Alwahibi, M. S., Elshikh, M. S., Hussain, M. I., Zulfiqar, F., Mansoor, S., & Abbas, Z. (2022). Biomass Production and Predicted Ethanol Yield Are Linked with Optimum Photosynthesis in Phragmites karka under Salinity and Drought Conditions. Plants, 11(13), 1657. https://doi.org/10.3390/plants11131657