
Synergistic Interaction between Symbiotic N2 Fixing Bacteria and Bacillus strains to Improve Growth, Physiological Parameters, Antioxidant Enzymes and Ni Accumulation in Faba Bean Plants (Vicia faba) under Nickel Stress



Abstract
:1. Introduction
2. Results
2.1. Assessment of Different Rhizobium Isolates and Bacillus Strains for Ni Tolerance
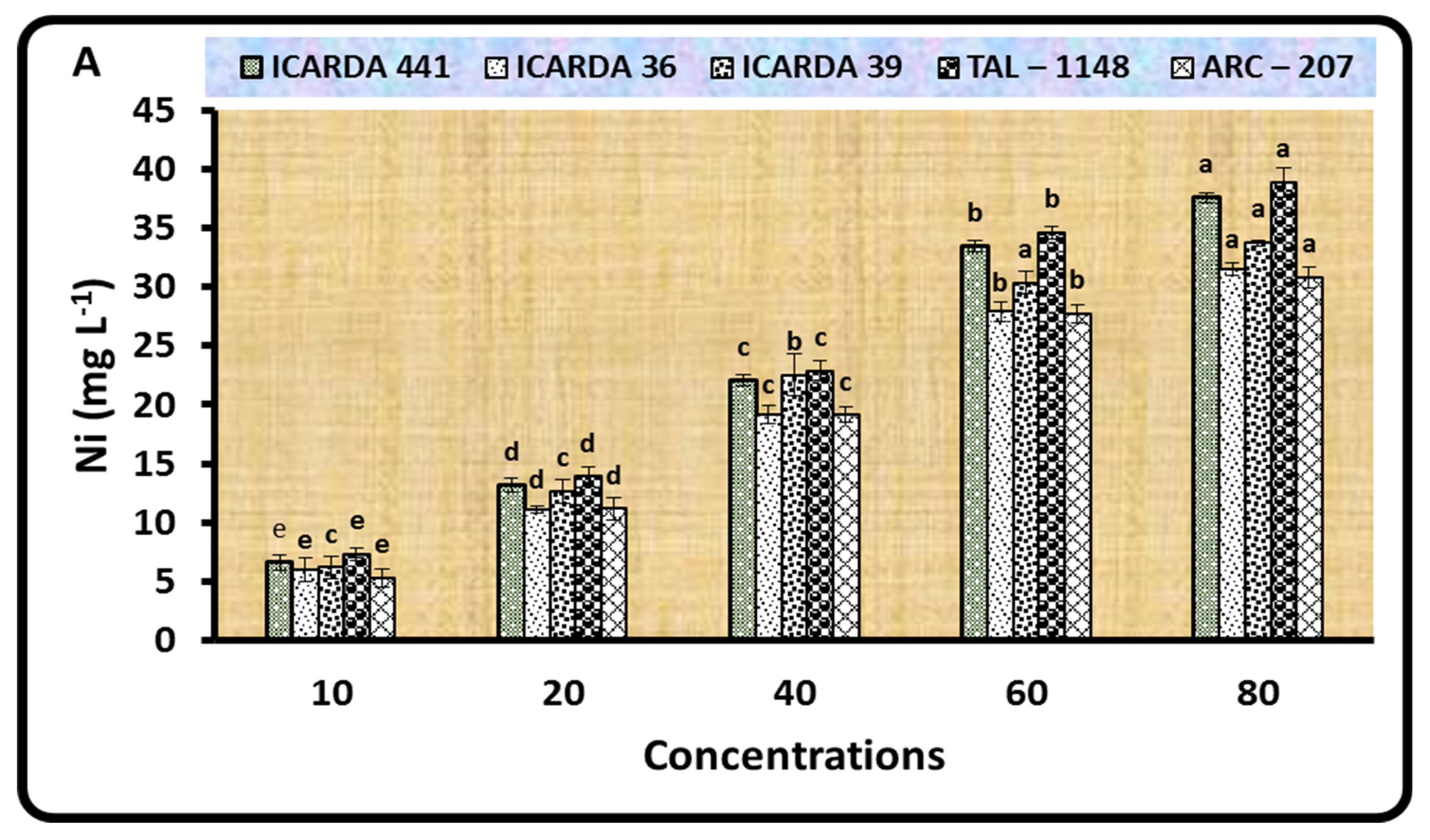
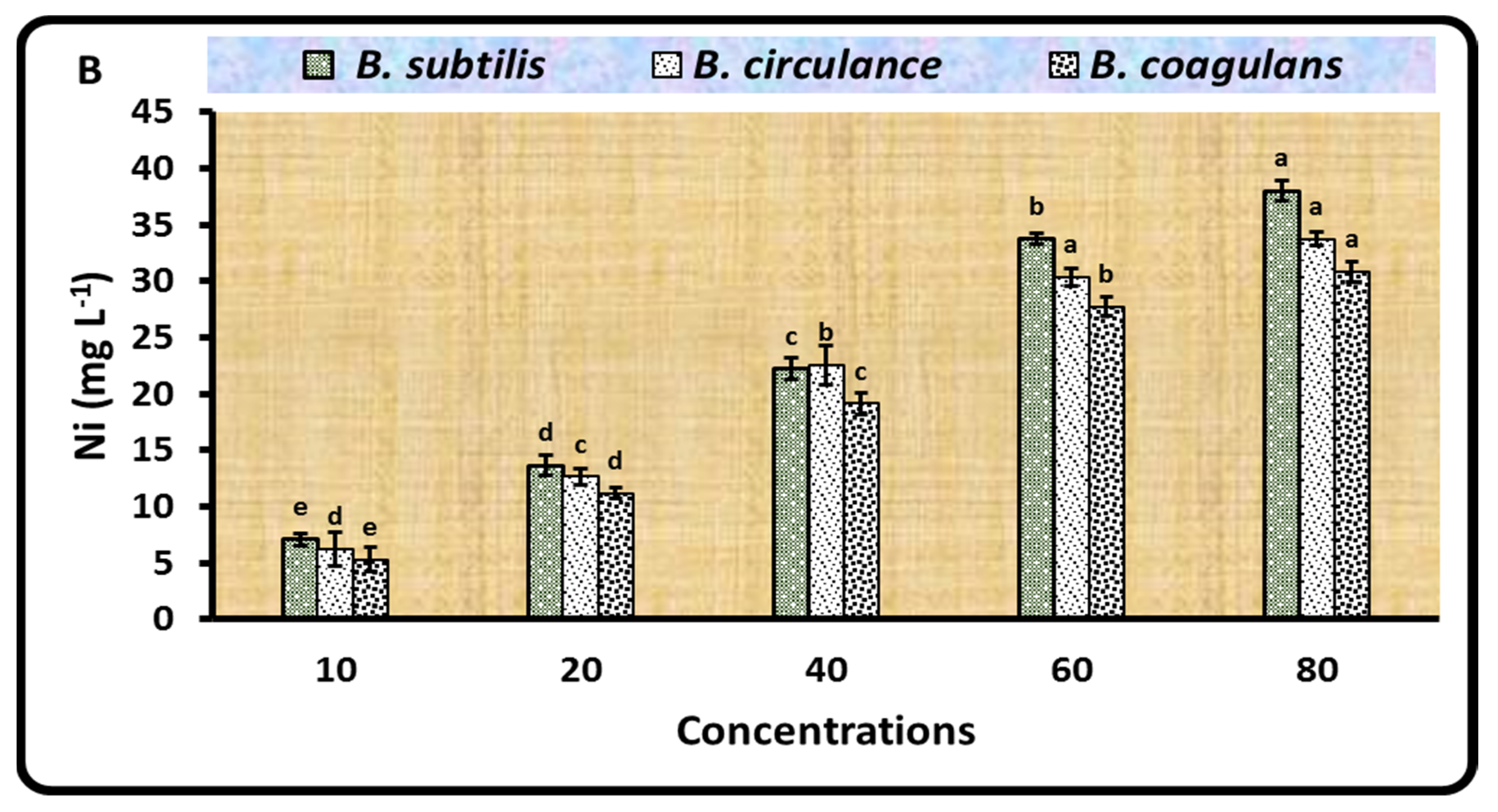
2.2. Biosorption of Ni by Different Rhizobium Isolates and Bacillus Strains
2.3. Pot Trial
2.3.1. Parameters of the Growth
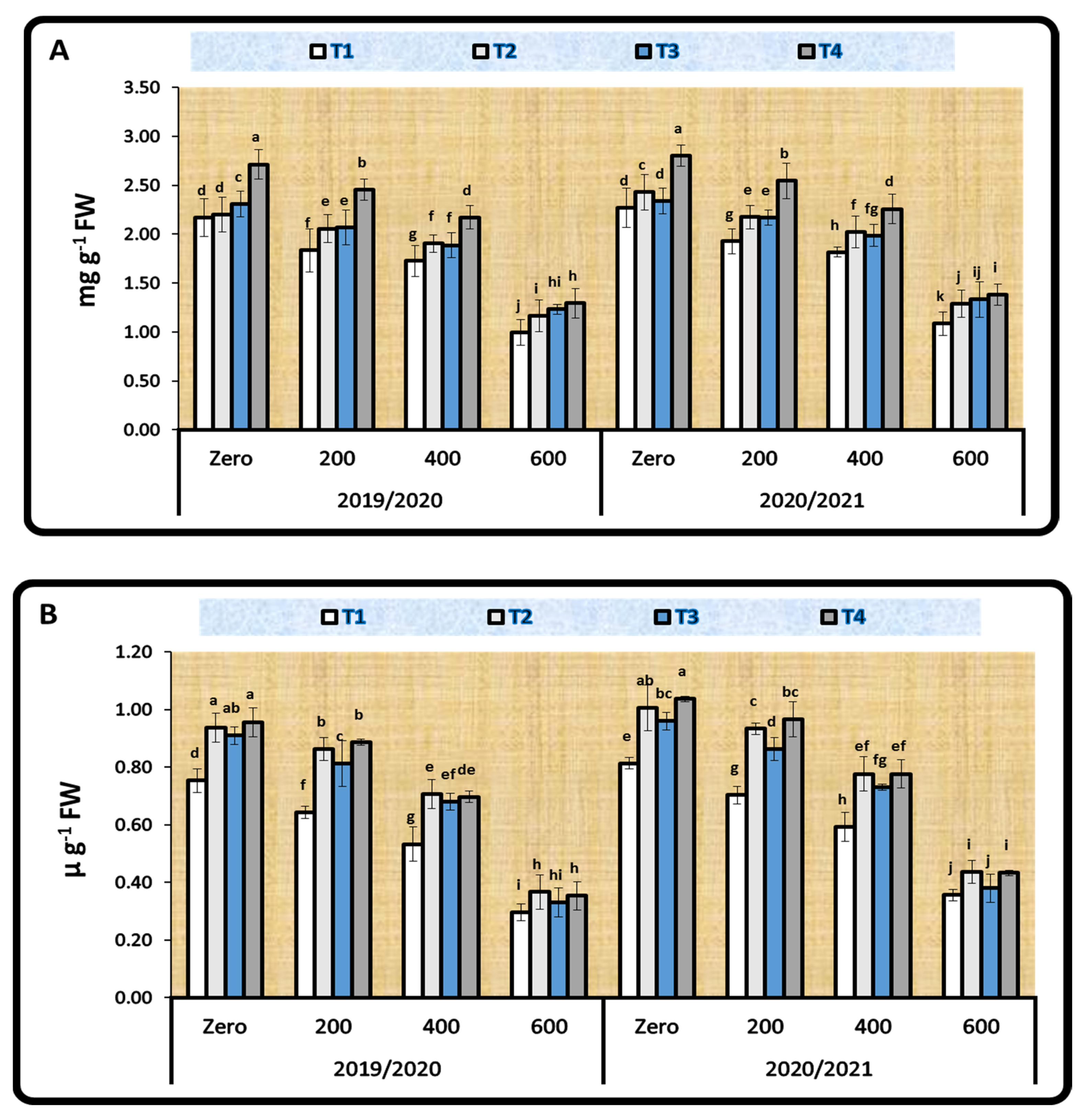
2.3.2. Photosynthetic Pigments
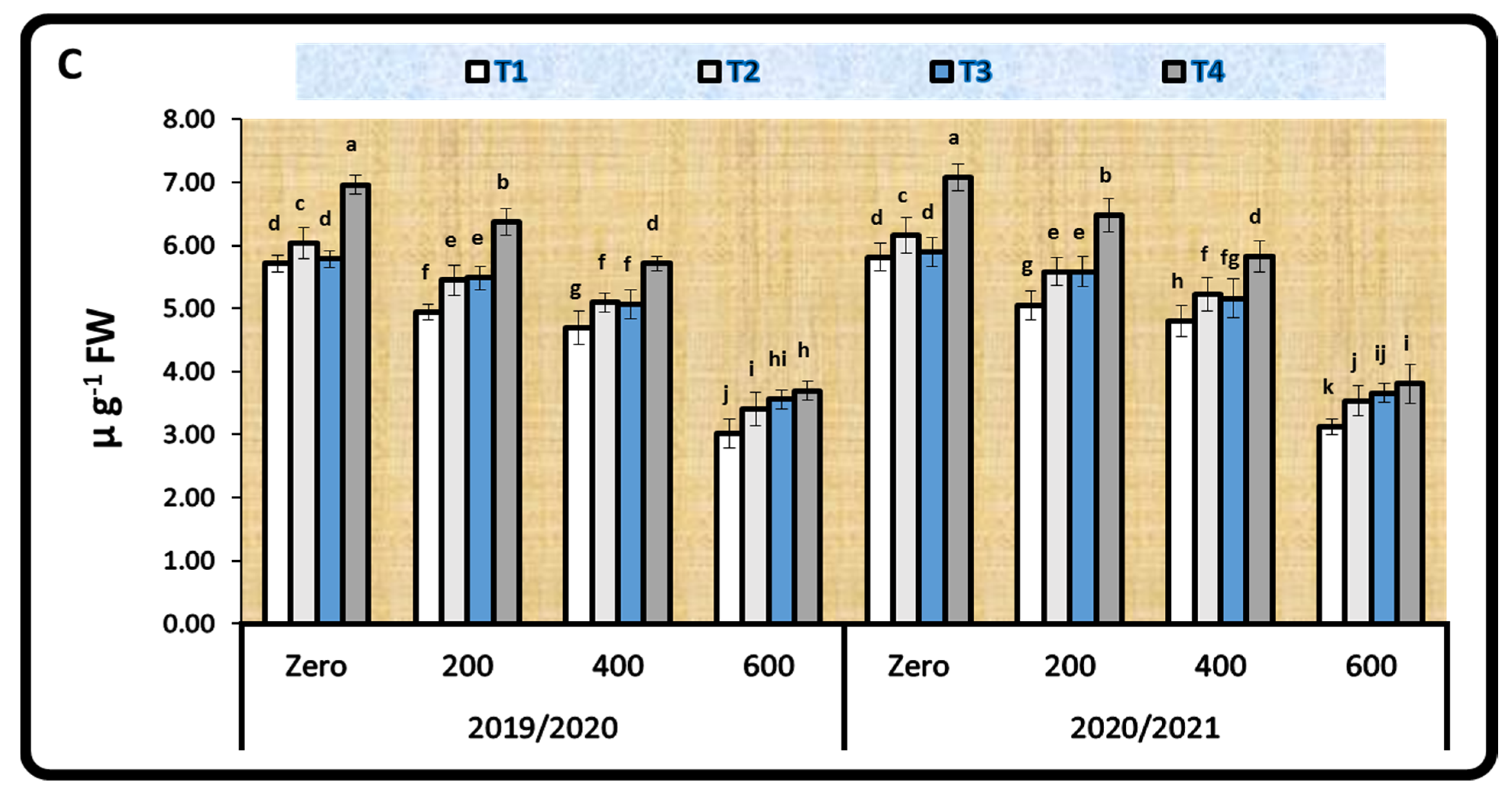
2.3.3. Antioxidant Enzyme Activity
2.3.4. Nickel Content
3. Discussion
3.1. Assessment of Different Rhizobium Isolates and Bacillus Strains for Ni Tolerance
3.2. Biosorption of Ni by Different Rhizobium Isolates and Bacillus Strains
3.3. Pot Trial
3.3.1. Parameters of Growth
3.3.2. Photosynthetic Pigments
3.3.3. Activity of Antioxidant Enzymes
3.3.4. Nickel Content
4. Materials and Methods
4.1. Microorganisms and Growth Medium
4.2. Assessment of Different Rhizobium Isolates and Bacillus Strains for Ni Tolerance
4.3. Biosorption of Ni by Different Rhizobium Isolates and Bacillus Strains
4.4. Pot Trial
4.4.1. Trait Measurements
4.4.2. Photosynthetic Pigments
4.4.3. Activity of Antioxidant Enzymes
4.4.4. Determination of Ni in the Roots and Shoots of the Faba Bean Plants
4.4.5. Bioconcentration and Translocation Factors
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rawal, V.; Navarro, D.K. The Global Economy of Pulses; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2019. [Google Scholar]
- Etemadi, F.; Barker, A.V.; Hashemi, M.; Zandvakili, O.R.; Park, Y. Nutrient Accumulation in Faba Bean Varieties. Commun. Soil Sci. Plant Anal. 2018, 49, 2064–2073. [Google Scholar] [CrossRef]
- Mínguez, M.I.; Rubiales, D. Faba Bean. In Crop Physiology Case Histories for Major Crops; Sadras, V.O., Calderini, D.F., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 452–481. [Google Scholar]
- Beladi, M.; Habibi, D.; Kashani, A.; Paknejad, F.; Nooralvandi, T. Phytoremediation of Lead and Copper by Sainfoiin (Onobrychis vicifolia): Role of antioxidant enzymes and biochemical biomarkers. Am. Eurasian J. Agric. Environ. Sci. 2011, 3, 440–449. [Google Scholar]
- Dary, M.; Chamber-Pérez, M.A.; Palomares, A.J.; Pajuelo, E. “In situ” phytostabilisation of heavy metal polluted soils using Lupinus luteus inoculated with metal resistant plant-growth promoting rhizobacteria. J. Hazard. Mater. 2010, 177, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Krujatz, F.; Haarstrick, A.; Nörtemann, B.; Greis, T. Assessing the Toxic Effects of Nickel, Cadmium and EDTA on Growth of the Plant Growth-Promoting Rhizobacterium Pseudomonas brassicacearum. Water Air Soil Pollut. 2012, 223, 1281–1293. [Google Scholar] [CrossRef]
- Woźniak, K.; Basiak, J. Free radicals-mediated induction of oxidized DNA-bases and DNA protein cross-links by nickel chloride. Mutat. Res. 2003, 514, 233–243. [Google Scholar] [CrossRef]
- Salt, D.E.; Kato, N.; Krämer, U.; Smith, R.D.; Raskin, I. The role of root exudates in nickel hyperaccumulation and tolerance in accumulator and nonaccumulator species of Thlaspi. In Phytoremediation of Contaminated Soil and Water; Terry, N., Banuelos, G., Eds.; CRS Press LLC: London, UK, 2000; pp. 189–200. [Google Scholar]
- Orlov, D.S.; Sadovnikova, L.K.; Lozanovskaya, I.N. Ecology and Protection of Biosphere under Chemical Pollution; Vysshaya Shkola: Moscow, Russia, 2002. [Google Scholar]
- Jamil, M.; Zeb, S.; Anees, M.; Roohi, A.; Ahmed, I.; Rehman, S.U.; Rha, E.S. Role of Bacillus licheniformis in Phytoremediation of Nickel Contaminated Soil Cultivated with Rice. Int. J. Phytoremediat. 2014, 16, 554–571. [Google Scholar] [CrossRef] [PubMed]
- Kupper, H.; Kroneck, P.M.H. Nickel in the environment and its role in the metabolism of plants and cyanobacteria. In Metal Ions in Life Sciences.; Sigel, A., Sigel, H., Sigel, R.K.O., Eds.; Wiley: Chichester, UK, 2007; pp. 31–62. [Google Scholar]
- Pandey, S.N.; Singh, K. Effect of nickel-stresses on uptake, pigments and antioxidative responses of water lettuce Pistia stratiotes L. J. Environ. Biol. 2011, 32, 391–394. [Google Scholar]
- Khan, W.U.; Yasin, N.A.; Ahmad, S.R.; Ali, A.; Ahmed, S.; Ahmad, A. Role of Ni-tolerant Bacillus spp. and Althea rosea L. in the phytoremediation of Ni-contaminated soils. Int. J. Phytoremediat. 2017, 19, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Edulamudi, P.; Masilamani, A.J.A.; Vanga, U.R.; Divi, V.R.S.G.; Konada, V.M. Nickel tolerance and biosorption potential of rhizobia associated with horse gram [Macrotyloma uniflorum (Lam.) Verdc.]. Int. J. Phytoremediat. 2021, 23, 1184–1190. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, B.M.; Lukoyanov, D.; Yang, Z.-Y.; Dean, D.R.; Seefeldt, L.C. Mechanism of Nitrogen Fixation by Nitrogenase: The Next Stage. Chem. Rev. 2014, 114, 4041–4062. [Google Scholar] [CrossRef]
- Higgins, K. Nickel Metalloregulators and Chaperones. Inorganics 2019, 7, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abd-Alla, M.H.; Issa, A.A.; Ohyama, T. Impact of harsh environmental conditions on nodule formation and dinitrogen fixation of legumes. In Advances in Biology and Ecology of Nitrogen Fixation; Ohyama, T., Ed.; InTech Open: Rijeka, Croatia, 2014; p. 9. [Google Scholar]
- Abd-Alla, M.H.; Morsy, F.M.; El-Enany, A.-W.E.; Ohyama, T. Isolation and characterization of a heavy-metal-resistant isolate of Rhizobium leguminosarum bv. viciae potentially applicable for biosorption of Cd2+ and Co2+. Int. Biodeterior. Biodegrad. 2012, 67, 48–55. [Google Scholar] [CrossRef]
- Maynaud, G.; Brunel, B.; Mornico, D.; Durot, M.; Severac, D.; Dubois, E.; Navarro, E.; Cleyet-Marel, J.-C.; Le Quéré, A. Genome-wide transcriptional responses of two metal-tolerant symbiotic Mesorhizobium isolates to Zinc and Cadmium exposure. BMC Genom. 2013, 14, 292. [Google Scholar] [CrossRef] [Green Version]
- Wani, P.A.; Khan, M.S. Nickel Detoxification and Plant Growth Promotion by Multi Metal Resistant Plant Growth Promoting Rhizobium Species RL9. Bull. Environ. Contam. Toxicol. 2013, 91, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Marzena, S.R.; Dorota, K.; Krzysztof, G.; Joanna, B.; Tomasz, S. Lotus corniculatus-rhizobia symbiosis under Ni, Co and Cr stress on ultramafic soil. Plant Soil 2020, 451, 459–484. [Google Scholar]
- Zheng, Y.; Xue, Q.-Y.; Xu, L.-L.; Xu, Q.; Lu, S.; Gu, C.; Guo, J.-H. A screening strategy of fungal biocontrol agents towards Verticillium wilt of cotton. Biol. Control 2011, 56, 209–216. [Google Scholar] [CrossRef]
- Tobin, J.M.; Cooper, D.G.; Neufeld, R.J. Uptake of Metal Ions by Rhizopus arrhizus Biomass. Appl. Environ. Microbiol. 1984, 47, 821–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pradhan, S.; Rai, L. Biotechnological potential of Microcystis sp. in Cu, Zn and Cd biosorption from single and multimetallic systems. BioMetals 2001, 14, 67–74. [Google Scholar] [CrossRef]
- Ansari, M.I.; Malik, A. Biosorption of nickel and cadmium by metal resistant bacterial isolates from agricultural soil irrigated with industrial wastewater. Bioresour. Technol. 2007, 98, 3149–3153. [Google Scholar] [CrossRef] [PubMed]
- Ajmal, A.W.; Saroosh, S.; Mulk, S.; Hassan, M.N.; Yasmin, H.; Jabeen, Z.; Nosheen, A.; Shah, S.M.U.; Naz, R.; Hasnain, Z.; et al. Bacteria Isolated from Wastewater Irrigated Agricultural Soils Adapt to Heavy Metal Toxicity While Maintaining Their Plant Growth Promoting Traits. Sustainability 2021, 13, 7792. [Google Scholar] [CrossRef]
- Outten, F.W.; Outten, C.E.; Halloran, T. Metallo regulatory systems at the interface between bacterial metal homeostasis and resistance. In Bacterial Stress Responses; Storz, G., Hengge, A.R., Eds.; ASM Press: Washington, DC, USA, 2000; pp. 29–42. [Google Scholar]
- Zaidi, S.; Usmani, S.; Singh, B.R.; Musarrat, J. Significance of Bacillus subtilis strain SJ-101 as a bioinoculant for concurrent plant growth promotion and nickel accumulation in Brassica juncea. Chemosphere 2006, 64, 991–997. [Google Scholar] [CrossRef]
- Hao, X.; Taghavi, S.; Xie, P.; Orbach, M.J.; Alwathnani, H.A.; Rensing, C.; Wei, G. Phytoremediation of Heavy and Transition Metals Aided by Legume-Rhizobia Symbiosis. Int. J. Phytoremediat. 2014, 16, 179–202. [Google Scholar] [CrossRef] [PubMed]
- Ibekwe, A.M.; Angle, J.S.; Chaney, R.L.; van Berkum, P. Sewage Sludge and Heavy Metal Effects on Nodulation and Nitrogen Fixation of Legumes. J. Environ. Qual. 1995, 24, 1199–1204. [Google Scholar] [CrossRef]
- Vijayarengan, P. Growth, nodulation and dry matter yield of black gram cultivars under nickel stress. J. Environ. Sci. Eng. 2000, 46, 151–158. [Google Scholar]
- Ishtiaq, S.; Mahmood, S. Phytotoxicity of nickel and its accumulation in tissues of three Vigna species at their early growth stages. J. Appl. Bot. Food Qual. 2011, 84, 223–228. [Google Scholar]
- Bethkey, P.C.; Drew, M.C. Stomatal and non-stomatal components to inhibition of photosynthesis in leaves of Capsium annum during progressive exposure to NaCl salinity. Plant Physiol. 1992, 99, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.M.; Hayat, S.; Ali, B.; Ahmad, A. Effect of 28-homobrassinolide treatment on nickel toxicity in Brassica juncea. Photosynthetica 2007, 45, 139–142. [Google Scholar] [CrossRef]
- Gajewska, E.; Skłodowska, M. Effect of nickel on ROS content and antioxidative enzyme activities in wheat leaves. BioMetals 2007, 20, 27–36. [Google Scholar] [CrossRef]
- Molas, J. Changes in morphological and anatomical structure of cabbage (Brassica oleracera L.) outer leaves and in ultrastructure of their chloroplasts caused by an in vitro excess of nickel. Photosynthetica 1997, 34, 513–522. [Google Scholar] [CrossRef]
- Esitken, A.; Pirlak, L.; Turan, M.; Sahin, F. Effects of floral and foliar application of plant growth promoting rhizobacteria (PGPR) on yield, growth and nutrition of sweet cherry. Sci. Hortic. 2006, 110, 324–327. [Google Scholar] [CrossRef]
- Nadeem, S.M.; Zahir, Z.A.; Naveed, M.; Arshad, M. Preliminary investigations on inducing salt tolerance in maize through inoculation with rhizobacteria containing ACC deaminase activity. Can. J. Microbiol. 2007, 53, 1141–1149. [Google Scholar] [CrossRef] [PubMed]
- Kohler, J.; Hernández, J.A.; Caravaca, F.; Roldán, A. Induction of antioxidant enzymes is involved in the greater effectiveness of a PGPR versus AM fungi with respect to increasing the tolerance of lettuce to severe salt stress. Environ. Exp. Bot. 2009, 65, 245–252. [Google Scholar] [CrossRef]
- Bhattacharyya, P.N.; Jha, D.K. Plant growth-promoting rhizobacteria (PGPR): Emergence in agriculture. World J. Microbiol. Biotechnol. 2012, 28, 1327–1350. [Google Scholar] [CrossRef]
- Heidari, M.; Golpayegani, A. Effects of water stress and inoculation with plant growth promoting rhizobacteria (PGPR) on antioxidant status and photosynthetic pigments in basil (Ocimum basilicum L.). J. Saudi Soc. Agric. Sci. 2012, 11, 57–61. [Google Scholar] [CrossRef] [Green Version]
- Elbagory, M.; Farrag, D.K.; Hashim, A.M.; Omara, A.E.-D. The Combined Effect of Pseudomonas stutzeri and Biochar on the Growth Dynamics and Tolerance of Lettuce Plants (Lactuca sativa) to Cadmium Stress. Horticulturae 2021, 7, 430. [Google Scholar] [CrossRef]
- Hafez, E.M.; Omara, A.E.D.; Alhumaydhi, F.A.; El-Esawi, M.A. Minimizing hazard impacts of soil salinity and water stress on wheat plants by soil application of vermicompost and biochar. Physiol. Plant. 2021, 172, 587–602. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Guo, H.; Wang, S.; Li, J.; Wang, Y.; Xing, B. Carbon dots alleviate the toxicity of cadmium ions (Cd2+) toward wheat seedlings. Environ. Sci. Nano 2019, 6, 1493–1506. [Google Scholar] [CrossRef]
- Unadkat, K.; Parikh, P. Localization of Cadmium metal ion in Lemna polyrhiza L. using SEM morphology and EDX analysis. Environ. Conserv. J. 2019, 20, 81–86. [Google Scholar] [CrossRef]
- Mamaril, J.C.; Paner, E.T.; Alpante, B.M. Biosorption and desorption studies of chromium (III) by free and immobilized Rhizobium (BJVr 12) cell biomass. Biogeochemistry 1997, 8, 275–285. [Google Scholar] [CrossRef]
- Callow, J.A. Review of A Manual for the Practical Study of Root-Nodule Bacteria; IBP Hand Book No. 15; Blackwell Scientific Publishing: Hoboken, NJ, USA, 1970; p. 164. [Google Scholar]
- Atlas, R.M. Handbook of Microbiological Media, 4th ed.; CRC Press: Boca Raton, FL, USA, 2010; p. 2040. [Google Scholar]
- Massadeh, A.M.; Al-Momani, F.A.; Haddad, H.I. Removal of Lead and Cadmium by Halophilic Bacteria Isolated from the Dead Sea Shore, Jordan. Biol. Trace Element Res. 2005, 108, 259–270. [Google Scholar] [CrossRef]
- Skradleta, V.; Gaudinova, A.; Necova, M.; Hydrakova, A. Behaviour of nodulated Pisum sativum L. under short term nitrate stress conditions. Biol. Plant 1984, 26, 364. [Google Scholar] [CrossRef]
- Page, A.L. Methods of Soil Analysis, Part 2. Chemical and Microbiological Properties, 2nd ed.; American Society of Agronomy, Inc.: Madison, WI, USA; Soil Science Society of America, Inc.: Madison, WI, USA, 1982. [Google Scholar]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. In Methods in Enzymology; Academic Press: San Diego, CA, USA, 1987; Volume 148, pp. 350–382. [Google Scholar]
- Hendrix, D.L. Rapid Extraction and Analysis of Nonstructural Carbohydrates in Plant Tissues. Crop Sci. 1993, 33, 1306–1311. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen Peroxide is Scavenged by Ascorbate-specific Peroxidase in Spinach Chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Rao, M.V.; Paliyath, C.; Ormrod, D.P.; Murr, D.P.; Watkins, C.B. Influence of salicylic acid on H2O2 production, oxidative stress and H2O2-metabolizing enzymes: Salicylic acidmediated oxidative damage requires H2O2. Plant Physiol. 1997, 115, 137–149. [Google Scholar] [CrossRef] [Green Version]
- Malik, C.P.; Singh, M.B. Plant Enzymology and Histoenzymology; Kalyani Publishers: Delhi, India, 1980; pp. 54–56. [Google Scholar]
- Humphries, E.C. Mineral components and ash analysis. Mod. Methods Plant Anal. 1956, 1, 468–502. [Google Scholar]
- Baker, A.J.M. Accumulators and excluders-strategies in the response of plants to heavy metals. J. Plant Nutr. 1981, 3, 643–654. [Google Scholar] [CrossRef]
- Embrandiri, A.; Rupani, P.F.; Shahadat, M.; Singh, R.P.; Ismail, S.A.; Ibrahim, M.H.; Kadir, M.O.A. The phytoextraction potential of selected vegetable plants from soil amended with oil palm decanter cake. Int. J. Recycl. Org. Waste Agric. 2017, 6, 37–45. [Google Scholar] [CrossRef] [Green Version]
- Duncan, D.B. Multiple range and multiple F tests. Biometrics 1955, 11, 1–42. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria Test | Ni Concentrations (mg L−1) | |||||
|---|---|---|---|---|---|---|
| 0 | 10 | 20 | 40 | 60 | 80 | |
| Rhizobiumisolates | ||||||
| ICARDA 441 | 4.83 ± 0.06 a | 4.30 ± 0.26 b | 3.70 ± 0.26 c | 3.00 ± 0.10 d | 2.60 ± 0.26 d | 1.84 ± 0.06 e |
| ICARDA 36 | 4.87 ± 0.06 a | 4.20 ± 0.16 b | 3.67 ± 0.15 c | 3.17 ± 0.21 d | 2.94 ± 0.04 d | 1.57 ± 0.04 e |
| ICARDA 39 | 4.87 ± 0.06 a | 4.43 ± 0.06 a | 3.40 ± 0.44 b | 2.40 ± 0.10 c | 1.60 ± 0.26 d | 0.82 ± 0.05 e |
| TAL–1148 | 4.93 ± 0.15 a | 4.73 ± 0.15 a | 4.07 ± 0.15 b | 3.63 ± 0.15 c | 2.97 ± 0.15 d | 2.53 ± 0.06 e |
| ARC–207 | 4.70 ± 0.02 a | 4.43 ± 0.59 a | 3.63 ± 0.15 b | 2.63 ± 0.10 c | 1.80 ± 0.10 d | 1.20 ± 0.10 d |
| Bacillusstrains | ||||||
| B. subtilis | 2.83 ± 0.06 a | 2.70 ± 0.10 a | 2.33 ± 0.06 b | 1.67 ± 0.15 c | 1.13 ± 0.15 d | 0.84 ± 0.06 e |
| B. circulance | 2.87 ± 0.06 a | 2.20 ± 0.10 b | 1.67 ± 0.15 c | 1.17 ± 0.21 d | 0.94 ± 0.04 d | 0.57 ± 0.04 e |
| B. coagulans | 2.87 ± 0.06 a | 2.43 ± 0.06 b | 2.07 ± 0.15 c | 1.40 ± 0.10 d | 1.03 ± 0.12 e | 0.82 ± 0.05 e |
| Treatments | Dry Weight (g Plant−1) | Plant Height (cm Plant−1) | Number of Nodules | N (%) |
|---|---|---|---|---|
| First Season (2019/2020) | ||||
| 0 T1 | 3.32 ± 0.55 h | 33.88 ± 1.48 h | 65.00 ± 5.00 f | 2.12 ± 0.15 h |
| 0 T2 | 4.17 ± 0.61 c | 42.59 ± 1.56 c | 74.00 ± 4.00 d | 2.97 ± 0.25 c |
| 0 T3 | 3.69 ± 0.64 e | 37.75 ± 1.41 e | 87.00 ± 6.00 b | 2.49 ± 0.14 e |
| 0 T4 | 4.46 ± 0.46 a | 45.56 ± 1.57 a | 98.00 ± 5.00 a | 3.26 ± 0.26 a |
| 200 T1 | 3.02 ± 0.60 j | 30.91 ± 1.41 j | 54.67 ± 4.51 h | 1.82 ± 0.14 j |
| 200 T2 | 3.98 ± 0.44 d | 40.63 ± 1.42 d | 63.67 ± 4.51 f | 2.78 ± 0.24 c |
| 200 T3 | 3.47 ± 0.53 fg | 35.45 ± 1.32 fg | 76.67 ± 4.51 c | 2.27 ± 0.23 fg |
| 200 T4 | 4.28 ± 0.70 b | 43.71 ± 1.46 b | 87.67 ± 4.51 b | 3.08 ± 0.14 b |
| 400 T1 | 2.87 ± 0.54 k | 29.31 ± 1.37 k | 43.67 ± 3.51 i | 1.67 ± 0.14 k |
| 400 T2 | 3.93 ± 0.62 d | 40.14 ± 1.19 d | 52.67 ± 3.51 h | 2.73 ± 0.22 c |
| 400 T3 | 3.45 ± 0.74 g | 35.28 ± 1.40 g | 65.67 ± 3.51 f | 2.25 ± 0.34 g |
| 400 T4 | 4.17 ± 0.54 c | 42.59 ± 1.37 c | 76.67 ± 3.51 c | 2.97 ± 0.24 c |
| 600 T1 | 2.53 ± 0.66 l | 25.85 ± 1.62 l | 25.33 ± 2.52 j | 1.33 ± 0.26 l |
| 600 T2 | 3.53 ± 0.75 f | 36.05 ± 1.56 f | 45.33 ± 3.06 i | 2.33 ± 0.15 f |
| 600 T3 | 3.16 ± 0.83 i | 32.28 ± 1.33 i | 58.33 ± 3.06 g | 1.96 ± 0.13 i |
| 600 T4 | 3.92 ± 0.44 d | 40.10 ± 1.40 d | 69.33 ± 3.06 e | 2.72 ± 0.24 c |
| Second Season (2020/2021) | ||||
| 0 T1 | 3.53 ± 0.56 h | 34.76 ± 1.14 h | 68.00 ± 7.20 g | 2.25 ± 0.37 i |
| 0 T2 | 4.35 ± 0.49 c | 43.41 ± 1.90 c | 78.00 ± 2.10 e | 3.08 ± 0.27 c |
| 0 T3 | 3.91 ± 0.24 f | 38.60 ± 1.19 e | 90.00 ± 6.30 c | 2.58 ± 0.36 f |
| 0 T4 | 4.62 ± 0.66 a | 46.47 ± 1.35 a | 103.00 ± 4.20 a | 3.33 ± 0.52 a |
| 200 T1 | 3.23 ± 0.82 j | 31.79 ± 1.90 j | 57.67 ± 3.81 i | 1.95 ± 0.61 k |
| 200 T2 | 4.16 ± 0.94 d | 41.45 ± 1.95 d | 67.67 ± 3.71 g | 2.89 ± 0.49 d |
| 200 T3 | 3.69 ± 0.53 g | 36.30 ± 1.78 fg | 79.67 ± 6.71 de | 2.36 ± 0.48 h |
| 200 T4 | 4.44 ± 0.74 b | 44.62 ± 1.39 b | 92.67 ± 6.31 b | 3.15 ± 0.76 b |
| 400 T1 | 3.08 ± 0.94 k | 30.19 ± 1.62 k | 46.67 ± 7.11 k | 1.80 ± 0.92 l |
| 400 T2 | 4.11 ± 0.35 de | 40.96 ± 1.83 d | 56.67 ± 7.81 i | 2.84 ± 0.19 de |
| 400 T3 | 3.67 ± 0.63 g | 36.13 ± 1.78 g | 68.67 ± 3.61 g | 2.34 ± 0.37 h |
| 400 T4 | 4.33 ± 0.81 c | 43.50 ± 1.11 c | 81.67 ± 5.41 d | 3.04 ± 0.28 c |
| 600 T1 | 2.74 ± 0.93 l | 26.73 ± 1.77 l | 28.33 ± 4.32 l | 1.46 ± 0.17 m |
| 600 T2 | 3.71 ± 0.89 g | 36.87 ± 1.41 f | 49.33 ± 6.26 j | 2.44 ± 0.14 g |
| 600 T3 | 3.38 ± 0.49 i | 33.13 ± 1.48 i | 61.33 ± 6.16 h | 2.05 ± 0.51 j |
| 600 T4 | 4.08 ± 0.77 e | 41.01 ± 1.39 d | 74.33 ± 3.16 f | 2.79 ± 0.61 e |
| F-test | ||||
| Main | ** | ** | ** | ** |
| Sub main | ** | ** | ** | ** |
| Interaction | ** | ** | ** | ** |
| Treatments | CAT | APX | PPO |
|---|---|---|---|
| First Season (2019/2020) | |||
| 0 T1 | 10.41 ± 1.16 i | 275.21 ± 17.29 l | 0.35 ± 0.09 i |
| 0 T2 | 19.05 ± 1.59 ef | 393.07 ± 22.55 f | 0.63 ± 0.12 ef |
| 0 T3 | 16.64 ± 1.39 g | 347.36 ± 34.86 i | 0.55 ± 0.11 g |
| 0 T4 | 22.15 ± 1.98 d | 439.50 ± 41.67 d | 0.74 ± 0.13 d |
| 200 T1 | 14.13 ± 1.05 h | 315.93 ± 40.33 k | 0.47 ± 0.10 h |
| 200 T2 | 22.39 ± 1.91 d | 441.29 ± 32.23 d | 0.75 ± 0.12 d |
| 200 T3 | 18.20 ± 1.14 f | 382.71 ± 34.67 g | 0.61 ± 0.08 f |
| 200 T4 | 23.90 ± 2.65 c | 468.79 ± 41.33 c | 0.80 ± 0.05 c |
| 400 T1 | 14.47 ± 1.28 h | 334.86 ± 24.83 j | 0.48 ± 0.08 h |
| 400 T2 | 24.02 ± 2.64 c | 447.00 ± 43.91 d | 0.80 ± 0.09 c |
| 400 T3 | 18.67 ± 1.44 ef | 385.57 ± 38.46 fg | 0.62 ± 0.11 ef |
| 400 T4 | 27.00 ± 2.21 b | 482.00 ± 35.39 b | 0.90 ± 0.09 b |
| 600 T1 | 17.00 ± 1.10 g | 369.86 ± 25.67 h | 0.57 ± 0.02 g |
| 600 T2 | 27.54 ± 3.74 b | 470.93 ± 29.50 c | 0.92 ± 0.05 b |
| 600 T3 | 19.56 ± 2.47 e | 413.07 ± 34.83 e | 0.65 ± 0.11 e |
| 600 T4 | 32.13 ± 2.33 a | 503.79 ± 36.69 a | 1.07 ± 0.11 a |
| Second season (2020/2021) | |||
| 0 T1 | 11.07 ± 2.66 i | 289.21 ± 19.11 m | 0.41 ± 0.01 i |
| 0 T2 | 19.76 ± 1.09 ef | 414.07 ± 22.05 g | 0.68 ± 0.09 e |
| 0 T3 | 17.19 ± 1.19 g | 364.36 ± 39.26 j | 0.58 ± 0.08 g |
| 0 T4 | 22.77 ± 2.28 d | 459.50 ± 24.17 e | 0.81 ± 0.06 d |
| 200 T1 | 14.79 ± 2.31 h | 329.93 ± 34.63 l | 0.54 ± 0.09 h |
| 200 T2 | 23.10 ± 3.91 d | 462.29 ± 32.03 de | 0.80 ± 0.12 d |
| 200 T3 | 18.75 ± 1.67 f | 399.71 ± 29.57 h | 0.64 ± 0.10 f |
| 200 T4 | 24.52 ± 2.28 c | 488.79 ± 36.13 c | 0.87 ± 0.11 c |
| 400 T1 | 15.13 ± 1.90 h | 348.86 ± 27.55 k | 0.55 ± 0.08 gh |
| 400 T2 | 24.73 ± 2.74 c | 468.00 ± 44.08 d | 0.85 ± 0.09 c |
| 400 T3 | 19.22 ± 2.33 ef | 402.57 ± 42.12 h | 0.65 ± 0.01 ef |
| 400 T4 | 27.62 ± 3.88 b | 502.00 ± 45.25 b | 0.97 ± 0.08 b |
| 600 T1 | 17.66 ± 2.19 g | 383.86 ± 35.50 i | 0.63 ± 0.13 f |
| 600 T2 | 28.25 ± 2.94 b | 491.93 ± 25.29 c | 0.97 ± 0.08 b |
| 600 T3 | 20.11 ± 3.97 e | 430.07 ± 32.41 f | 0.68 ± 0.12 e |
| 600 T4 | 32.75 ± 3.09 a | 523.79 ± 46.31 a | 1.14 ± 0.14 a |
| F-test | |||
| Main | ** | ** | ** |
| Sub main | ** | ** | ** |
| Interaction | ** | ** | ** |
| Treatments | Ni Content in Root (µg g−1) | Ni Content in Shoots (µg g−1) | Bioconcentration Factor (BCF) | Translocation Factor (TF) |
|---|---|---|---|---|
| First Season (2019/2020) | ||||
| 0 T1 | 0.00 | 0.00 | 0.00 | 0.00 |
| 0 T2 | 0.00 | 0.00 | 0.00 | 0.00 |
| 0 T3 | 0.00 | 0.00 | 0.00 | 0.00 |
| 0 T4 | 0.00 | 0.00 | 0.00 | 0.00 |
| 200 T1 | 46.87 ± 6.16 e | 15.19 ± 1.87 fg | 0.23 ± 0.03 a | 0.32 ± 0.03 e |
| 200 T2 | 32.08 ± 2.75 g | 11.74 ± 2.47 g | 0.16 ± 0.09 d | 0.36 ± 0.07 e |
| 200 T3 | 33.54 ± 3.47 g | 15.27 ± 2.33 fg | 0.16 ± 0.05 cd | 0.45 ± 0.06 d |
| 200 T4 | 27.84 ± 4.33 h | 14.54 ± 1.98 fg | 0.13 ± 0.02 e | 0.52 ± 0.03 c |
| 400 T1 | 85.17 ± 7.66 b | 56.83 ± 4.30 b | 0.21 ± 0.05 b | 0.66 ± 0.04 b |
| 400 T2 | 42.03 ± 7.21 f | 17.69 ± 2.53 f | 0.10 ± 0.06 fg | 0.42 ± 0.06 e |
| 400 T3 | 47.49 ± 6.55 de | 21.82 ± 3.11 e | 0.11 ± 0.09 f | 0.45 ± 0.04 d |
| 400 T4 | 41.72 ± 4.55 f | 18.72 ± 3.67 f | 0.10 ± 0.01 fg | 0.44 ± 0.07 e |
| 600 T1 | 97.71 ± 5.50 a | 77.37 ± 3.95 a | 0.16 ± 0.05 c | 0.79 ± 0.01 a |
| 600 T2 | 50.90 ± 5.17 d | 29.90 ± 2.39 d | 0.08 ± 0.01 h | 0.58 ± 0.02 c |
| 600 T3 | 56.67 ± 4.17 c | 38.67 ± 2.27 c | 0.09 ± 0.01 gh | 0.68 ± 0.03 b |
| 600 T4 | 47.70 ± 6.57 de | 24.28 ± 2.69 e | 0.07 ± 0.01 h | 0.50 ± 0.02 d |
| Second season (2020/2021) | ||||
| 0 T1 | 0.00 | 0.00 | 0.00 | 0.00 |
| 0 T2 | 0.00 | 0.00 | 0.00 | 0.00 |
| 0 T3 | 0.00 | 0.00 | 0.00 | 0.00 |
| 0 T4 | 0.00 | 0.00 | 0.00 | 0.00 |
| 200 T1 | 47.21 ± 4.33 d | 17.32 ± 1.20 f | 0.23 ± 0.01 a | 0.36 ± 0.01 e |
| 200 T2 | 34.47 ± 3.11 fg | 13.02 ± 2.41 g | 0.17 ± 0.03 d | 0.37 ± 0.03 e |
| 200 T3 | 34.32 ± 2.41 f | 15.61 ± 2.23 fg | 0.17 ± 0.02 d | 0.45 ± 0.04 d |
| 200 T4 | 30.40 ± 5.25 g | 15.99 ± 1.45 f | 0.15 ± 0.05 e | 0.52 ± 0.08 c |
| 400 T1 | 90.81 ± 4.17 b | 53.96 ± 4.13 b | 0.22 ± 0.02 b | 0.59 ± 0.09 b |
| 400 T2 | 44.42 ± 5.56 e | 16.97 ± 2.21 f | 0.11 ± 0.04 fg | 0.38 ± 0.05 e |
| 400 T3 | 50.27 ± 5.36 d | 23.16 ± 3.45 e | 0.12 ± 0.03 f | 0.46 ± 0.07 d |
| 400 T4 | 44.28 ± 5.78 e | 17.17 ± 3.72 f | 0.11 ± 0.02 fg | 0.38 ± 0.04 e |
| 600 T1 | 115.73 ± 4.32 a | 79.50 ± 3.54 a | 0.19 ± 0.02 c | 0.68 ± 0.05 a |
| 600 T2 | 53.46 ± 6.22 d | 27.35 ± 2.61 d | 0.08 ± 0.01 h | 0.51 ± 0.11 c |
| 600 T3 | 59.45 ± 5.09 c | 36.01 ± 2.52 c | 0.09 ± 0.02 gh | 0.60 ± 0.08 b |
| 600 T4 | 50.09 ± 4.47 d | 22.56 ± 2.86 e | 0.08 ± 0.01 h | 0.45 ± 0.06 d |
| F-test | ||||
| Main | ** | ** | ** | ** |
| Sub main | ** | ** | ** | ** |
| Interaction | ** | ** | ** | ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elbagory, M.; El-Nahrawy, S.; Omara, A.E.-D. Synergistic Interaction between Symbiotic N2 Fixing Bacteria and Bacillus strains to Improve Growth, Physiological Parameters, Antioxidant Enzymes and Ni Accumulation in Faba Bean Plants (Vicia faba) under Nickel Stress. Plants 2022, 11, 1812. https://doi.org/10.3390/plants11141812
Elbagory M, El-Nahrawy S, Omara AE-D. Synergistic Interaction between Symbiotic N2 Fixing Bacteria and Bacillus strains to Improve Growth, Physiological Parameters, Antioxidant Enzymes and Ni Accumulation in Faba Bean Plants (Vicia faba) under Nickel Stress. Plants. 2022; 11(14):1812. https://doi.org/10.3390/plants11141812
Chicago/Turabian StyleElbagory, Mohssen, Sahar El-Nahrawy, and Alaa El-Dein Omara. 2022. "Synergistic Interaction between Symbiotic N2 Fixing Bacteria and Bacillus strains to Improve Growth, Physiological Parameters, Antioxidant Enzymes and Ni Accumulation in Faba Bean Plants (Vicia faba) under Nickel Stress" Plants 11, no. 14: 1812. https://doi.org/10.3390/plants11141812
APA StyleElbagory, M., El-Nahrawy, S., & Omara, A. E. -D. (2022). Synergistic Interaction between Symbiotic N2 Fixing Bacteria and Bacillus strains to Improve Growth, Physiological Parameters, Antioxidant Enzymes and Ni Accumulation in Faba Bean Plants (Vicia faba) under Nickel Stress. Plants, 11(14), 1812. https://doi.org/10.3390/plants11141812