
Comprehensive Comparison of Chemical Composition and Antioxidant Activity of Panax ginseng Sprouts by Different Cultivation Systems in a Plant Factory

 ,
, 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Comparison of Fatty Acid Content in Different Cultivation Systems
2.2. Comparison of Free Amino Acid Contents in Different Cultivation Systems
2.3. Comparison of Mineral Contents in different Cultivation Systems
2.4. Comparison of Phytochemical Contents of Different Cultivation Systems
2.4.1. Ginsenosides
2.4.2. Phenolic Acids and Flavonols
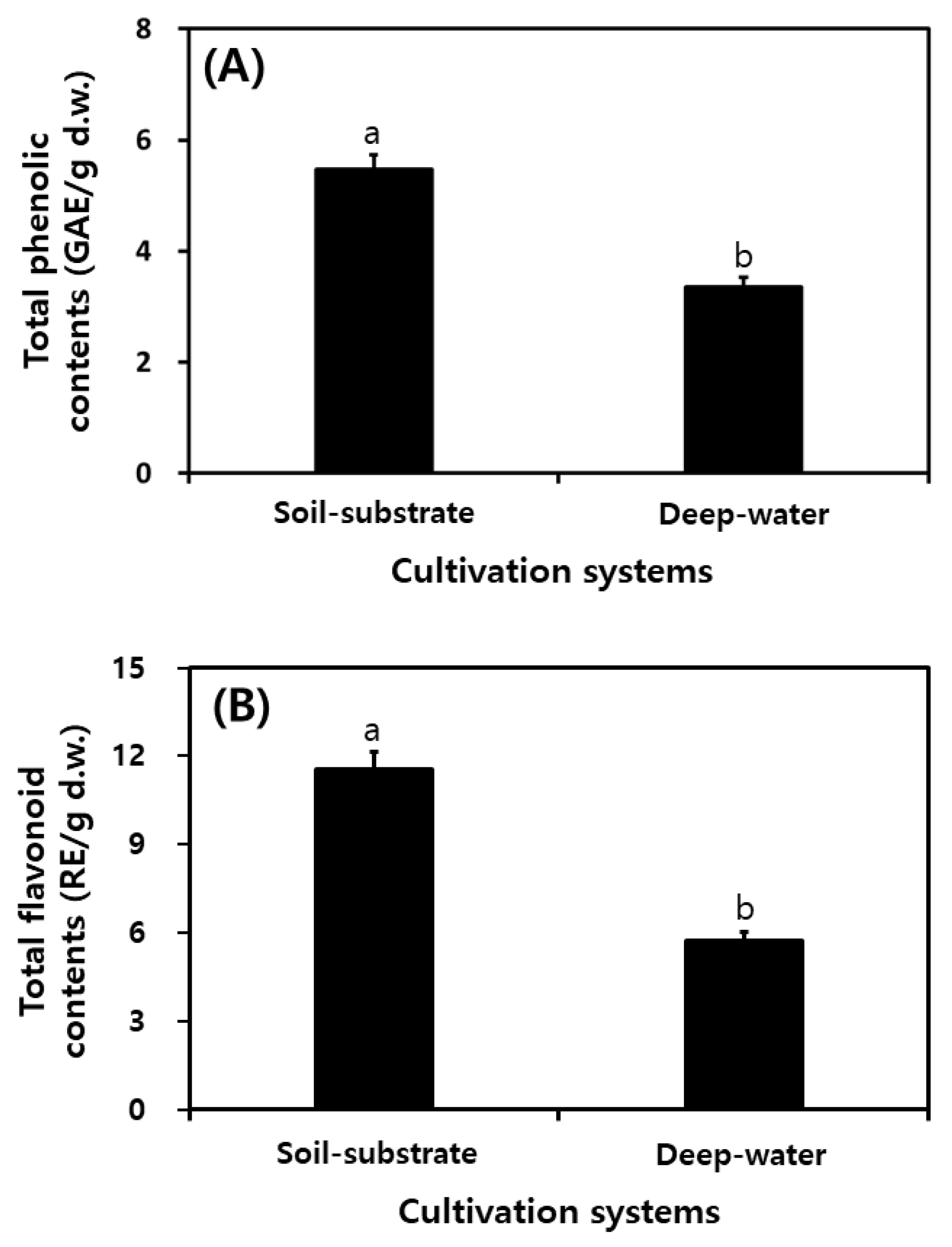
2.5. Comparison of Total Phenolic Contents and Total Flavonoid Contents in Different Cultivation Systems
2.6. Comparison of Antioxidant Effects in different Cultivation Systems
3. Materials and Methods
3.1. Chemicals and Instruments
3.2. Growth of PGSs by Different Cultivation Methods
3.3. Nutrient Compounds Analysis
3.3.1. FA Analysis
3.3.2. FAAs Analysis
3.3.3. Mineral Analysis
3.4. Phytochemical Compounds Analysis
3.4.1. Extract Preparation
3.4.2. Ginsenoside Compounds Analysis
3.4.3. Phenolic Acid and Flavonol Compounds Analysis
3.5. TPCs and TFCs Analysis
3.5.1. Preparation of Samples for TPCs and TFCs
3.5.2. TPCs Analysis
3.5.3. TFCs Analysis
3.6. Antioxidant Activities
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- O’Grady, M.J.O.; O’Hare, G.M.P. Modelling the smart farm. Inf. Process. Agric. 2017, 4, 179–187. [Google Scholar] [CrossRef]
- Zamora-Izquierdo, M.A.; Santa, J.; Martínez, J.A.; Martínez, V.; Skarmeta, A.F. Smart farming IoT platform based on edge and cloud computing. Biosyst. Eng. 2019, 177, 4–17. [Google Scholar] [CrossRef]
- Kuo, Y.H.; Ikegami, F.; Lambein, F. Neuroactive and other free amino acids in seed and young plants of Panax ginseng. Phytochemistry 2003, 62, 1087–1091. [Google Scholar] [CrossRef]
- Kim, G.S.; Hyun, D.Y.; Kim, Y.O.; Lee, S.E.; Kwon, H.; Cha, S.W.; Park, C.B.; Kim, Y.B. Investigation of ginsenosides in different parts of Panax ginseng cultured by hydroponics. Korean J. Hortic. Sci. 2010, 28, 216–226. [Google Scholar]
- Kim, S.C.; Kang, Y.M.; Seong, J.A.; Lee, H.Y.; Cho, D.Y.; Joo, O.S.; Lee, J.H.; Cho, K.M. Comprehensive changes of nutritional constituents and antioxidant activities of ginseng sprouts according to the roasting process. Korean J. Food Preserv. 2021, 28, 72–87. [Google Scholar] [CrossRef]
- Hwang, S.H.; Kim, S.C.; Seong, J.A.; Lee, H.Y.; Cho, D.Y.; Kim, M.J.; Jung, J.G.; Jeong, E.H.; Son, K.H.; Cho, K.M. Comparison of ginsenoside contents and antioxidant activity according to the size of ginseng sprout has produced in a plant factory. J. Appl. Biol. Chem. 2021, 64, 253–261. [Google Scholar] [CrossRef]
- Hwang, S.I.; Joo, J.M.; Joo, S.Y. ICT-Based smart farm factory systems through the case of hydroponic ginseng plant factory. J. Korean Inst. Commun. Inf. Sci. 2015, 40, 780–790. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.J.; Huang, L.L.; Cai, X.J.; Li, P.; Wang, Y.T.; Wan, J.R. Fatty acid variability in three medicinal herbs of Panax species. Chem. Cent. J. 2013, 7, 12. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.H.; Popova, E.V.; Hahn, E.J.; Paek, K.Y. Linoleic and α-linolenic fatty acids affect biomass and secondary metabolite production and nutritive properties of Panax ginseng adventitious roots cultured in bioreactors. Biochem. Eng. J. 2009, 47, 109–115. [Google Scholar] [CrossRef]
- Kim, Y.S.; Lim, Y.H.; Wang, S.G.; Park, S.J.; Park, C.R. The physicochemical properties and antioxidant effects of Samul Chol-Pyon. J. Korean Soc. Food Sci. Nutr. 1999, 28, 990–996. [Google Scholar]
- Cho, E.J.; Piao, X.L.; Jang, M.H.; Baek, S.H.; Kim, H.Y.; Kang, K.S.; Kwon, S.W.; Park, J.H. The effect of steaming on the free amino acid contents and antioxidant activity of Panax ginseng. Food Chem. 2008, 107, 876–882. [Google Scholar] [CrossRef]
- Wan, J.Y.; Fan, Y.; Yu, Q.T.; Ge, Y.Z.; Yan, C.P.; Alolga, R.N.; Li, P.; Ma, Z.H.; Qi, L.W. Integrated evaluation of malonyl ginsenosides, amino acids and polysaccharides in fresh and processed ginseng. J. Pharm. Biomed. Anal. 2015, 107, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Lee, G.A.; Heo, S.J.; Jeong, H.N.; Song, B.H. Comparative analysis on concentration and uptake amount of major mineral nutrients in plant tissues and years old of Panax ginseng C. A. Meyer. Korean J. Med. Crop Sci. 2012, 20, 195–201. [Google Scholar] [CrossRef] [Green Version]
- Shin, Y.A. Effect of physical activity, and sodium and potassium intakes on blood pressure and obesity. Korean Soc. Wellness 2016, 11, 409–423. [Google Scholar] [CrossRef]
- Shibata, S.; Fujita, M.; Itokawa, H.; Tanaka, O.; Ishii, T. Studies on the constituents of Japanese and Chinese crude drugs. XI. Panaxadiol, a sapogenin of ginseng roots. Chem. Pharm. Bull. 1963, 11, 759–761. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.H.; Lee, D.; Lee, H.L.; Kim, C.E.; Jung, K.; Kang, K.S. Beneficial effects of Panax ginseng for the treatment and prevention of neurodegenerative diseases: Past findings and future directions. J. Ginseng Res. 2018, 42, 239–247. [Google Scholar] [CrossRef]
- Abe, I.; Rohmer, M.; Prestwich, G.D. Enzymatic cyclization of squalene and oxidosqualene to sterols and triterpenes. Chem. Rev. 1993, 93, 2189–2206. [Google Scholar] [CrossRef]
- Kim, T.D.; Kim, Y.S.; Han, J.Y.; Lim, S.; Choi, Y.E. Metabolic engineering for production of ginsenosides on panax ginseng. J. Plant Biotechnol. 2009, 36, 352–359. [Google Scholar] [CrossRef] [Green Version]
- Shibuya, M.; Hoshino, M.; Katsube, Y.; Hayashi, H.; Kuchiro, T.; Ebizuka, Y. Identification of β-amyrin and sophoradiol 24-hydroxylase by expressed sequence tah mining and functional expression assay. FEBS J. 2006, 273, 948–959. [Google Scholar] [CrossRef]
- Kushiro, T.; Shibuya, M.; Ebuzuka, Y. β-amyrin synthase: Cloning of oxidosqualene cyclase that catalyzes the formation of the most popular triterpene among higher plants. Eur. J. Biochem. 1998, 256, 238–244. [Google Scholar] [CrossRef]
- Haralampidis, K.; Trojanowska, M.; Osbourn, A.E. Biosynthesis of triterpenoid saponin in plants. Adv. Biochem. Eng. Biotechnol. 2001, 75, 31–49. [Google Scholar] [CrossRef]
- Choi, D.W.; Jung, J.D.; Ha, Y.I.; Park, H.W.; In, D.S.; Chung, H.J.; Liu, J.R. Analysis of transcripts in methyl jasmonate-treated ginseng hairy roots to identify genes involved in the bio synthesis of ginsenosides and other secondary metabolites. Plant Cell Rep. 2005, 23, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Nguyen, T.K.L.; Oh, M.M. Growth and Ginsenosides Content of Ginseng Sprouts According to LED-Based Light Quality Changes. Agronomy 2020, 10, 1979. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, S.C.; Lee, H.Y.; Cho, D.Y.; Jung, J.G.; Kang, D.; Kang, S.S.; Cho, K.M. Changes in nutritional compositions of processed mountain-cultivated ginseng sprouts (Panax ginseng) and screening for their antioxidant and anti-inflammatory properties. J. Funct. Foods 2021, 86, 104668. [Google Scholar] [CrossRef]
- Eom, S.J.; Hwang, J.E.; Kim, H.S.; Kim, K.T.; Paik, H.D. Anti-inflammatory and cytotoxic effects of ginseng extract bioconverted by Leuconostoc mesenteroides KCCM 12010P isolated from kimchi. Int. J. Food Sci. Technol. 2018, 53, 1331–1337. [Google Scholar] [CrossRef]
- Hua, M.; Lu, J.; Qu, D.; Liu, C.; Zhang, L.; Li, S.; Chen, J.; Sun, Y. Structure, physicochemical properties and adsorption function of insoluble dietary fiber from ginseng residue: A potential functional ingredient. Food Chem. 2019, 286, 522–529. [Google Scholar] [CrossRef]
- Kim, S.S.; Cha, H.C. Comparison of the total phenolic and flavonoid contents and antioxidant activities of four kinds of sand dune plants living in Taean, Korea. Plant Resour. Soc. Korea 2017, 30, 8–16. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.H.; Lee, Y.C.; Kim, S.S.; Hong, H.D.; Kim, K.T. Quality and antioxidant activity of ginseng seed processed by fermentation strains. J. Ginseng Res. 2015, 39, 178–182. [Google Scholar] [CrossRef] [Green Version]
- Cho, K.M.; Lee, H.Y.; Lee, Y.M.; Seo, E.Y.; Kim, D.H.; Son, K.H.; Lee, J.H.; Cho, D.Y.; Lee, J.H. Comparative assessment of compositional constituents and antioxidant effects in ginseng sprouts (Panax ginseng) through aging and fermentation processes. LWT 2002, 164, 113644. [Google Scholar] [CrossRef]
- Song, Y.N.; Hong, H.G.; Son, J.S.; Kwon, Y.O.; Lee, H.H.; Kim, H.J.; Park, J.H.; Son, M.J.; Oh, J.G.; Yoon, M.H. Investigation of ginsenosides and antioxidant activities in the roots, leaves, and stems of hydroponic-cultured ginseng (Panax ginseng Meyer). Prev. Nutr. Food Sci. 2019, 24, 283–292. [Google Scholar] [CrossRef]
- Lee, M.H.; Rhee, Y.K.; Choi, S.Y.; Cho, C.W.; Hong, H.D.; Kim, K.T. Quality and characteristics of fermented ginseng seed oil based on bacterial strain and extraction method. J. Ginseng Res. 2017, 41, 428–433. [Google Scholar] [CrossRef] [PubMed]
- Chung, I.M.; Lim, J.J.; Ahn, M.S.; Jeong, H.N.; An, T.J.; Kim, S.H. Comparative phenolic compound profiles and antioxidant activity of the fruit, leaves, and roots of Korean ginseng (Panax ginseng Meyer) according to cultivation years. J. Ginseng Res. 2016, 40, 68–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.S.; Seong, B.J.; Kim, G.H.; Kim, S.I.; Han, S.H.; Baik, N.D. Ginsenoside, phenolic acid composition and physiological significances of fermented ginseng leaf. J. Korean Soc. Food Sci. Nutr. 2010, 39, 1194–1200. [Google Scholar] [CrossRef]
- Singh, B.N.; Shankar, S.; Srivastava, R.K. Green tea catechin, epigallocatechin-3-gallate (EGCG): Mechanisms, perspectives and clinical applications. Biochem. Pharmacol. 2011, 82, 1807–1821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miean, K.H.; Mohamed, S. Flavonoid (myricetin, quercetin, kaempferol, luteolin, and apigenin) content of edible tropical plants. J. Agric. Food Chem. 2001, 49, 3106–3112. [Google Scholar] [CrossRef] [PubMed]
- Coleman, C.I.; Hebert, J.H.; Reddy, P. The effects of Panax ginseng on quality of life. J. Clin. Pharm. Ther. 2003, 28, 5–15. [Google Scholar] [CrossRef]
- Hyun, S.H.; Kim, S.W.; Seo, H.W.; Youn, S.H.; Kyung, J.S.; Lee, Y.Y.; In, G.; Park, C.-K.; Han, C.-K. Physiological and pharmacological features of the non-saponin components in Korean Red Ginseng. J. Ginseng Res. 2020, 44, 527–537. [Google Scholar] [CrossRef]
- Valanciene, E.; Jonuskiene, I.; Syrpas, M.; Augustiniene, E.; Matulis, P.; Simonavicus, A.; Malys, N. Advances and prospects of phenolic acids production biorefinery and analysis. Biomolecules 2020, 10, 874. [Google Scholar] [CrossRef]
- Hwang, C.R.; Lee, S.H.; Jang, G.Y.; Hwang, I.G.; Kim, H.Y.; Woo, K.S.; Lee, J.S.; Jeong, H.S. Changes in ginsenoside compositions and antioxidant activities of hydroponic-cultured ginseng roots and leaves with heating temperature. J. Ginseng Res. 2014, 38, 180–186. [Google Scholar] [CrossRef] [Green Version]
- Hwang, J.E.; Suh, D.H.; Kim, K.T.; Paik, H.D. Comparative study on anti-oxidant and anti-inflammatory properties of hydroponic ginseng and soil-cultured ginseng. Food Sci. Biotechnol. 2019, 28, 215–224. [Google Scholar] [CrossRef]
- Hwang, C.E.; Kim, S.C.; Kim, D.H.; Lee, H.Y.; Suh, H.K.; Cho, K.M.; Lee, J.H. Enhancement of isoflavone aglycone, amino acid, and CLA contents in fermented soybean yogurts using different strains: Screening of antioxidant and digestive enzyme inhibition properties. Food Chem. 2021, 340, 128199. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Hwang, C.E.; Son, K.S.; Cho, K.M. Comparisons of nutritional constituents in soybeans during solid state fermentation times and screening for their glucosides enzymes and antioxidant properties. Food Chem. 2019, 272, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, C.A.; Nicácio, A.E.; Boeing, J.S.; Garcia, F.P.; Nakamura, C.V.; Visentainer, J.V.; Maldaner, L. Rapid extraction method followed by a d-SPE clean-up step for determination of phenolic composition and antioxidant and antiproliferative activities from berry fruits. Food Chem. 2019, 309, 125694. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Hwang, C.E.; Cho, E.J.; Song, Y.H.; Kim, S.C.; Cho, K.M. Improvement of nutritional components and in vitro antioxidative properties of soy-powder yogurts using Lactobacillus plantarum. J. Food Drug Anal. 2018, 26, 1054–1065. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Yang, W.T.; Cho, K.M.; Lee, J.H. Comparative analysis of isoflavone aglycones using microwave-assisted acid hydrolysis from soybean organs at different growth times and screening for their digestive enzyme inhibition and antioxidant properties. Food Chem. 2020, 305, 125462. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Contents 1 (mg/100 g d.w.) | Cultivation Systems | |
|---|---|---|
| Soil–Substrate | Deep-Water | |
| Saturated fatty acids | ||
| Myristic acid (C14:0) | 5.2 ± 0.26 a | 4.9 ± 0.25 a |
| Palmitic acid (C16:0) | 207.4 ± 10.37 ab | 238.5 ± 11.93 a |
| Stearic acid (C18:0) | 33.1 ± 1.66 b | 56.6 ± 2.83 a |
| Arachidic acid (C20:0) | 4.7 ± 0.24 b | 6.8 ± 0.34 a |
| Behenic acid (C22:0) | 9.5 ± 0.48 b | 13.5 ± 0.68 a |
| Lignoceric acid (C24:0) | 8.3 ± 0.42 a | 9.0 ± 0.45 a |
| Total | 268.2 | 329.3 |
| Unsaturated fatty acids | ||
| Palmitoleic acid (C16:1) | 7.7 ± 0.39 | Nd 2 |
| Oleic acid (C18:1n9c) | 35.6 ± 1.78 a | 34.2 ± 1.71 a |
| Linoleic acid (C18:2c) | 397.6 ± 19.88 a | 390.7 ± 19.54 a |
| α-Linolenic acid (C18:3n3) | 222.6 ± 1.13 a | 176.0 ± 8.80 b |
| Eicosenic acid (C20:1) | 3.6 ± 0.18 a | 3.3 ± 0.17 a |
| Eicosadienoic acid (C20:2) | 3.7 ± 0.19 a | 3.6 ± 0.18 a |
| Eicosatrienoic acid (C20:3n3) | 6.1 ± 0.31 a | 5.7 ± 0.29 a |
| Tricosanoic acid (C23:0) | 5.5 ± 0.28 a | 4.8 ± 0.24 ab |
| Total | 682.4 | 618.3 |
| Total fatty acids | 950.6 | 947.6 |
| Contents 1 (mg/100 g d.w.) | Cultivation Systems | |
|---|---|---|
| Soil–Substrate | Deep-Water | |
| Non-essential amino acids (NEAAs) | ||
| Proline | 172.58 ± 8.63 a | 37.69 ± 1.88 b |
| Aspartic acid | 241.67 ± 12.08 a | 102.71 ± 5.14 b |
| Serine | 97.06 ± 4.85 a | 66.51 ± 3.33 b |
| Aspartic acid-NH2 | 313.37 ± 15.67 a | 131.57 ± 6.58 b |
| Glutamic acid | 165.22 ± 8.26 a | 56.96 ± 2.85 a |
| Sarcosine | 5.49 ± 0.27 a | nd 2 |
| Aminoadipic acid | 9.53 ± 0.48 a | 4.40 ± 0.22 a |
| Glycine | 18.32 ± 0.92 a | 12.50 ± 0.63 ab |
| Alanine | 108.21 ± 5.41 a | 86.98 ± 4.35 ab |
| Citrulline | 24.81 ± 1.24 a | 3.82 ± 0.19 b |
| α-aminobutyric acid | 13.58 ± 0.68 ab | 17.66 ± 0.88 a |
| Cystine | 9.58 ± 0.48 a | 8.45 ± 0.42 a |
| Tyrosine | 71.33 ± 3.57 a | 41.01 ± 2.05 b |
| β-alanine | 28.28 ± 1.41 a | 27.40 ± 1.37 a |
| β-aminoisobutyric acid | 27.09 ± 1.35 a | 16.25 ± 0.81 b |
| γ-aminobutyric acid | 247.75 ± 12.39 a | 163.38 ± 8.17 b |
| Aminoethanol | 24.47 ± 1.22 a | 20.90 ± 1.05 a |
| Hydroxylysine | 2.80 ± 0.14 a | 1.88 ± 0.09 b |
| Ornithine | 45.41 ± 2.27 a | 17.99 ± 0.90 b |
| 3-methylhistidine | 0.75 ± 0.04 | nd 2 |
| Arginine | 1365.53 ± 68.28 a | 746.80 ± 78.24 b |
| Total | 2992.83 | 1564.86 |
| Essential amino acids (EAAs) | ||
| Threonine | 72.59 ± 3.63 a | 47.31 ± 2.37 b |
| Valine | 102.81 ± 5.14 a | 66.75 ± 3.34 b |
| Methionine | 18.71 ± 0.94 a | 16.28 ± 0.81 a |
| Isoleucine | 68.73 ± 3.44 a | 44.12 ± 2.21 b |
| Leucine | 98.88 ± 4.94 a | 54.89 ± 2.74 b |
| Phenylalanine | 101.89 ± 5.09 a | 52.47 ± 2.62 b |
| Lysine | 90.05 ± 4.50 a | 66.03 ± 3.30 b |
| Histidine | 45.87 ± 2.29 a | 29.52 ± 1.48 b |
| Total | 599.53 | 377.37 |
| Total amino acids | 3592.36 | 1942.23 |
| Contents 1 (mg/g d.w.) | Cultivation Systems | |
|---|---|---|
| Soil–Substrate | Deep-Water | |
| Protopanaxtriol types | ||
| Ginsenoside Rg1 (1) | 4.11 ± 0.21 b | 12.63 ± 0.63 a |
| Ginsenoside Re (2) | 19.18 ± 0.96 a | 9.42 ± 0.47 b |
| Ginsenoside Rf (4) | 0.36 ± 0.02 b | 0.61 ± 0.03 a |
| Ginsenoside F5 (5) | 0.62 ± 0.03 b | 1.26 ± 0.06 a |
| Ginsenoside F3 (6) | 3.02 ± 0.15 a | 2.98 ± 0.15 a |
| Ginsenoside Rg2 (7) | 1.4 ± 0.07 a | 0.45 ± 0.02 b |
| Ginsenoside Rh1 (8) | nd 2 | nd |
| Ginsenoside F1 (11) | 1.13 ± 0.06 a | 0.62 ± 0.03 b |
| Protopanaxtriol (18) | 1.06 ± 0.05 a | 0.39 ± 0.02 b |
| Total | 30.88 | 28.36 |
| Protopanaxdiol types | ||
| Ginsenoside Rb1 (9) | 3.32 ± 0.17 a | 2.80 ± 0.14 ab |
| Ginsenoside Rc (10) | 2.80 ± 0.14 ab | 3.13 ± 0.16 a |
| Ginsenoside Rb2 (12) | 3.76 ± 0.19 a | 2.69 ± 0.13 b |
| Ginsenoside Rb3 (13) | 0.32 ± 0.02 b | 0.41 ± 0.02 a |
| Ginsenoside Rd (14) | 12.65 ± 0.63 a | 6.87 ± 0.34 b |
| Ginsenoside Rd2 (15) | 2.34 ± 0.12 b | 6.42 ± 0.32 a |
| Ginsenoside F2 (16) | 4.02 ± 0.20 b | 9.61 ± 0.48 a |
| Ginsenoside Rg3 (17) | 0.55 ± 0.03 a | 0.38 ± 0.02 b |
| Compound K (19) | 1.27 ± 0.06 a | 0.78 ± 0.04 b |
| Ginsenoside Rh2 (20) | 0.76 ± 0.04 a | 0.69 ± 0.03 a |
| Protopanaxdiol (21) | 0.55 ± 0.03 b | 1.05 ± 0.05 a |
| Total | 32.34 | 34.83 |
| Oleanane types | ||
| Ginsenoside Ro (3) | 1.93 ± 0.10b | 2.83 ± 0.14a |
| Total | 1.93 | 2.83 |
| Total ginsenosides | 65.15 | 66.01 |
| Contents 1 (μg/g d.w.) | Cultivation Systems | |
|---|---|---|
| Soil–Substrate | Deep-Water | |
| Phenolic acids | ||
| Gallic acid | 9.43 ± 0.47 a | 10.20 ± 0.51 a |
| Protocatechuic acid | 11.22 ± 0.56 a | 4.38 ± 0.22 b |
| Chlorogenic acid | 32.31 ± 1.62 a | 7.34 ± 0.37 b |
| p-hydroxybenzoic acid | 11.86 ± 0.59 a | 5.85 ± 0.29 b |
| Vanillic acid | 3.55 ± 0.18 a | 2.03 ± 0.10 b |
| p-coumaric acid | nd 2 | 1.23 ± 0.06 a |
| Ferulic acid | nd | nd |
| Veratric acid | 2.53 ± 0.13 a | nd |
| Benzoic acid | 62.16 ± 3.11 a | 27.97 ± 1.40 b |
| t-cinnamic acid | 0.30 ± 0.02 a | 0.23 ± 0.01 ab |
| Total | 133.36 | 59.23 |
| Flavonols | ||
| Epigallocatechin | 90.52 ± 4.53 ab | 115.19 ± 5.76 a |
| Catechin | 81.22 ± 4.06 a | 21.30 ± 1.07 b |
| Epicatechin | 9.59 ± 0.48 a | 5.38 ± 0.27 b |
| Epigallocatechin gallate | 21.83 ± 1.09 a | 21.44 ± 1.07 a |
| Vanillin | nd | nd |
| Rutin | nd | nd |
| Catechin gallate | nd | nd |
| Quercetin | 177.79 ± 8.89 a | 112.94 ± 5.65 b |
| Naringin | 1.87 ± 0.09 a | 0.62 ± 0.03 b |
| Naringenin | 5.37 ± 0.27 a | 1.29 ± 0.06 b |
| Formonoetin | nd | nd |
| Total | 388.19 | 278.16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, K.M.; Lee, H.Y.; Cho, D.Y.; Jung, J.G.; Kim, M.J.; Jeong, J.B.; Jang, S.-N.; Lee, G.O.; Sim, H.-S.; Kang, M.J.; et al. Comprehensive Comparison of Chemical Composition and Antioxidant Activity of Panax ginseng Sprouts by Different Cultivation Systems in a Plant Factory. Plants 2022, 11, 1818. https://doi.org/10.3390/plants11141818
Cho KM, Lee HY, Cho DY, Jung JG, Kim MJ, Jeong JB, Jang S-N, Lee GO, Sim H-S, Kang MJ, et al. Comprehensive Comparison of Chemical Composition and Antioxidant Activity of Panax ginseng Sprouts by Different Cultivation Systems in a Plant Factory. Plants. 2022; 11(14):1818. https://doi.org/10.3390/plants11141818
Chicago/Turabian StyleCho, Kye Man, Hee Yul Lee, Du Yong Cho, Jea Gack Jung, Min Ju Kim, Jong Bin Jeong, Seong-Nam Jang, Ga Oun Lee, Han-Sol Sim, Min Ji Kang, and et al. 2022. "Comprehensive Comparison of Chemical Composition and Antioxidant Activity of Panax ginseng Sprouts by Different Cultivation Systems in a Plant Factory" Plants 11, no. 14: 1818. https://doi.org/10.3390/plants11141818
APA StyleCho, K. M., Lee, H. Y., Cho, D. Y., Jung, J. G., Kim, M. J., Jeong, J. B., Jang, S. -N., Lee, G. O., Sim, H. -S., Kang, M. J., Kim, Y. L., Lee, J. H., Lim, S., & Son, K. -H. (2022). Comprehensive Comparison of Chemical Composition and Antioxidant Activity of Panax ginseng Sprouts by Different Cultivation Systems in a Plant Factory. Plants, 11(14), 1818. https://doi.org/10.3390/plants11141818