
Callus Induction from Diverse Explants and Genotypes Enables Robust Transformation of Perennial Ryegrass (Lolium perenne L.)


 , and
, and
Abstract
:1. Introduction
2. Results
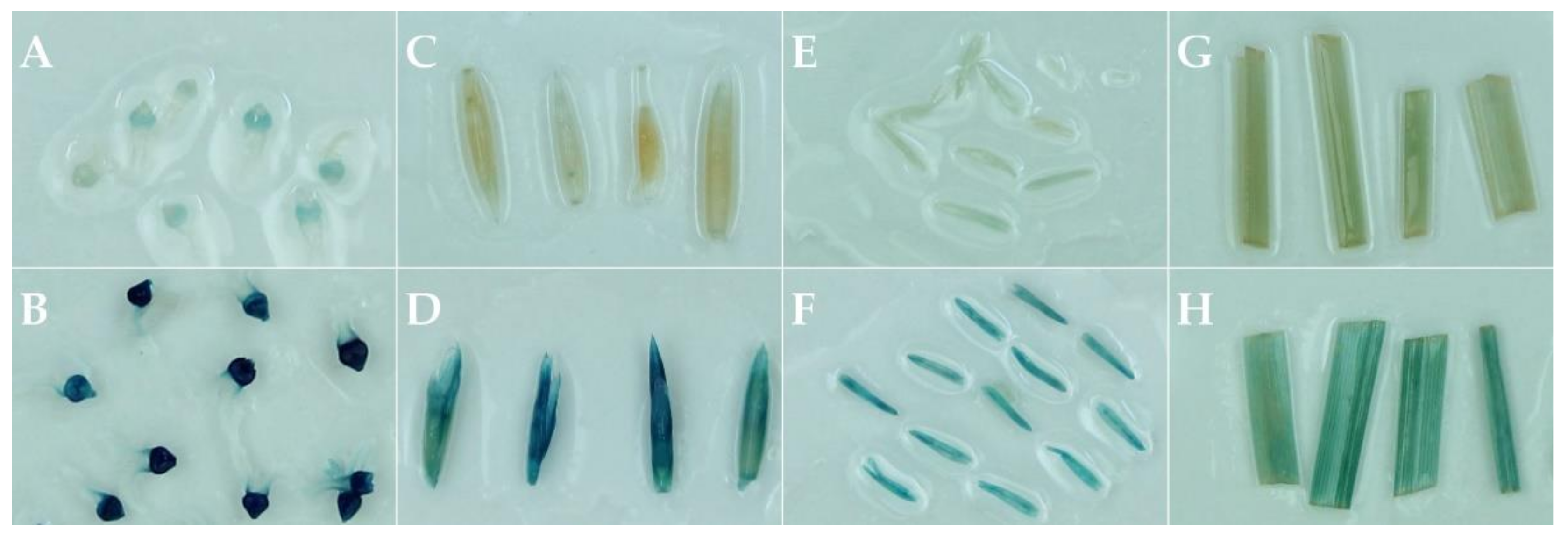
2.1. Callus Induction and Agrobacterium-Mediated Transformation
2.2. T-DNA Characterization in Transgenic Plants
2.3. Genes Activity in Transgenic Lines
3. Discussion
4. Materials and Methods
4.1. Plant Growth Conditions
4.1.1. Induction of Flowering
4.1.2. Establishment in the Soil
4.2. Callus Induction from Various Explants and Genotypes
4.3. Agrobacterium-Mediated Transformation
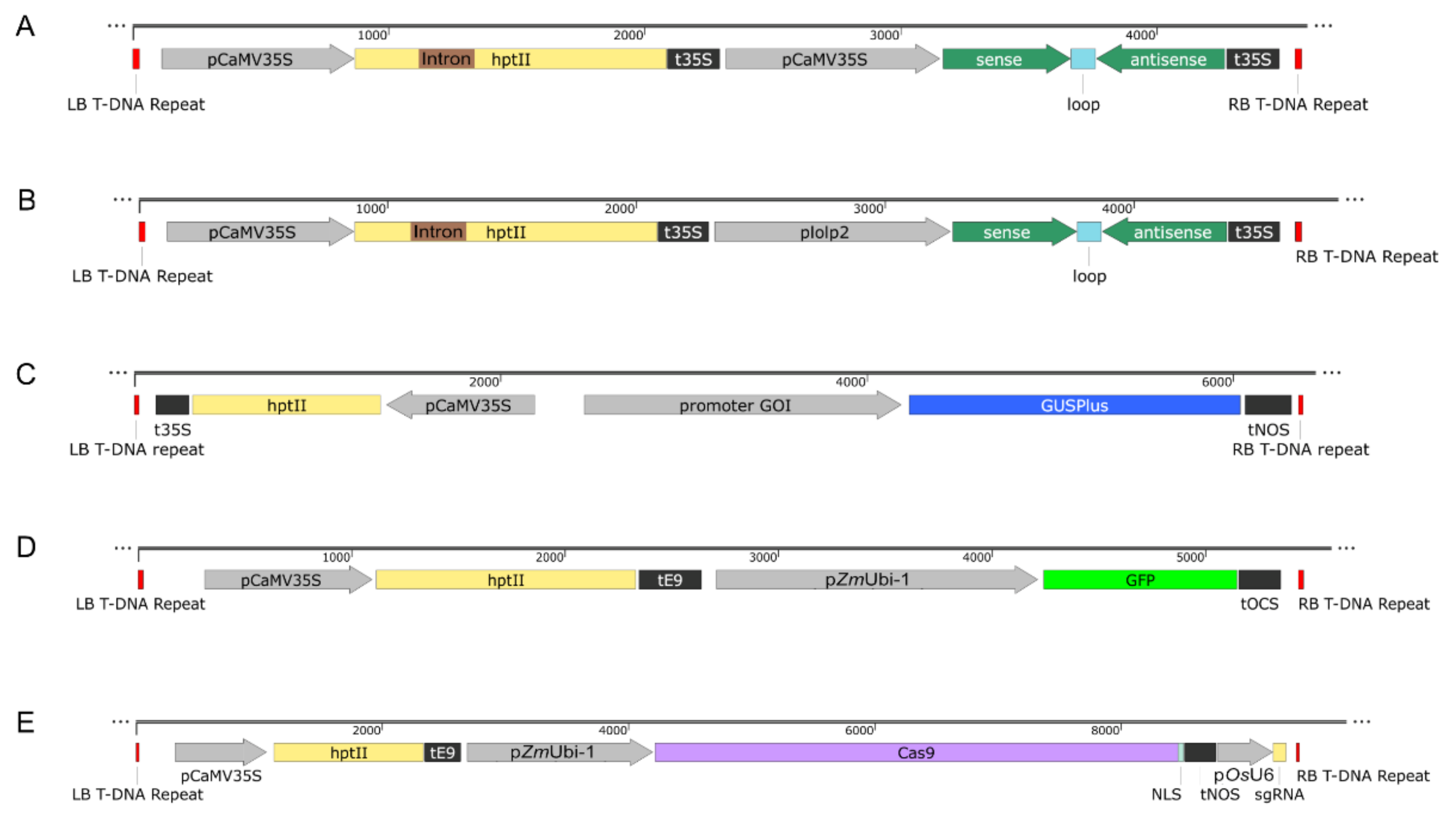
4.4. Plasmid Construction and Transformation into A. tumefaciens
4.5. Constructs Preparation
4.5.1. Creation of the Hairpin-Based RNAi Constructs
4.5.2. Creation of Promoter GUS Constructs
4.5.3. Acquisition of the GFP Construct
4.5.4. Creation of CRISPR Constructs
4.6. DNA Extraction and PCR Confirmation of T-DNA Integration
4.7. Fluorescence Microscopy
4.8. GUS Assay
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Humphreys, M.; Feuerstein, U.; Vandewalle, M.; Baert, J. Ryegrasses. In Fodder Crops and Amenity Grasses; Boller, B., Posselt, U.K., Veronesi, F., Eds.; Springer: New York, NY, USA, 2010; pp. 211–260. [Google Scholar]
- McDonagh, J.; McEvoy, M.; O’Donovan, M.; Gilliland, T.J. Genetic gain in yield of perennial ryegrass (Lolium perenne), Italian ryegrass (Lolium multiflorum Lam.) and hybrid ryegrass (Lolium x boucheanum Kunth) cultivars in Northern Ireland Recommended Lists 1972–2013. Futur. Eur. Grassl. 2014, 19, 836–839. [Google Scholar]
- Laidig, F.; Piepho, H.P.; Drobek, T.; Meyer, U. Genetic and non-genetic long-term trends of 12 different crops in German official variety performance trials and on-farm yield trends. Theor. Appl. Genet. 2014, 127, 2599–2617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, B.; Thorogood, D.; Armstead, I.; Barth, S. How far are we from unravelling self-incompatibility in grasses? New Phytol. 2008, 178, 740–753. [Google Scholar] [CrossRef] [PubMed]
- Frei, D.; Veekman, E.; Grogg, D.; Stoffel-Studer, I.; Morishima, A.; Shimizu-Inatsugi, R.; Yates, S.; Shimizu, K.K.; Frey, J.E.; Studer, B.; et al. Ultralong oxford nanopore reads enable the development of a reference-grade perennial ryegrass genome assembly. Genome Biol. Evol. 2021, 13, evab159. [Google Scholar] [CrossRef]
- Copetti, D.; Yates, S.A.; Vogt, M.M.; Russo, G.; Grieder, C.; Kölliker, R.; Studer, B. Evidence for high intergenic sequence variation in heterozygous Italian ryegrass (Lolium multiflorum Lam.) genome revealed by a high-quality draft diploid genome assembly. bioRxiv 2021. [Google Scholar] [CrossRef]
- Huang, L.; Feng, G.; Yan, H.; Zhang, Z.; Bushman, B.S.; Wang, J.; Bombarely, A.; Li, M.; Yang, Z.; Nie, G.; et al. Genome assembly provides insights into the genome evolution and flowering regulation of orchardgrass. Plant Biotechnol. J. 2020, 18, 373–388. [Google Scholar] [CrossRef]
- Benevenuto, J.; Ferrão, L.F.V.; Amadeu, R.R.; Munoz, P. How can a high-quality genome assembly help plant breeders? Gigascience 2019, 8, 1–4. [Google Scholar] [CrossRef]
- Martin, R.C.; Glover-Cutter, K.; Martin, R.R.; Dombrowski, J.E. Virus induced gene silencing in Lolium temulentum. Plant Cell Tissue Organ Cult. 2013, 113, 163–171. [Google Scholar] [CrossRef]
- Yu, G.; Cheng, Q.; Xie, Z.; Xu, B.; Huang, B.; Zhao, B. An efficient protocol for perennial ryegrass mesophyll protoplast isolation and transformation, and its application on interaction study between LpNOL and LpNYC1. Plant Methods 2017, 13, 46. [Google Scholar] [CrossRef]
- Adamski, N.; Borrill, P.; Brinton, J.; Harrington, S.; Marchal, C.; Bentley, A.; Bovill, W.; Cattivelli, L.; Cockram, J.; Contreras-Moreira, B.; et al. A roadmap for gene functional characterisation in wheat. PeerJ 2019, 7, e26877v2. [Google Scholar] [CrossRef] [Green Version]
- Altpeter, F.; Springer, N.M.; Bartley, L.E.; Blechl, A.E.; Brutnell, T.P.; Citovsky, V.; Conrad, L.J.; Gelvin, S.B.; Jackson, D.P.; Kausch, A.P.; et al. Advancing crop transformation in the era of genome editing. Plant Cell 2016, 28, 1510–1520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Maas, H.M.; de Jong, E.R.; Rueb, S.; Hensgens, L.A.M.; Krens, F.A. Stable transformation and long-term expression of the gusA reporter gene in callus lines of perennial ryegrass (Lolium perenne L.). Plant Mol. Biol. 1994, 24, 401–405. [Google Scholar] [CrossRef] [PubMed]
- Spangenberg, G.; Wang, Z.; Wu, X.; Nagel, J.; Potrykus, I. Transgenic perennial ryegrass (Lolium perenne) plants from microprojectile bombardment of embryogenic suspension cells. Plant Sci. 1995, 108, 209–217. [Google Scholar] [CrossRef]
- Wang, G.R.; Binding, H.; Posselt, U.K. Fertile transgenic plants from direct gene transfer to protoplasts of Lolium perenne L. and Lolium multiflorum Lam. J. Plant Physiol. 1997, 151, 83–90. [Google Scholar] [CrossRef]
- Dalton, S.J.; Bettany, A.J.E.; Timms, E.; Morris, P. Transgenic plants of Lolium multiflorum, Lolium perenne, Festuca arundinacea and Agrostis stolonifera by silicon carbide fibre-mediated transformation of cell suspension cultures. Plant Sci. 1998, 132, 31–43. [Google Scholar] [CrossRef]
- Dalton, S.J.; Bettany, A.J.E.; Timms, E.; Morris, P. Co-transformed, diploid Lolium perenne (perennial ryegrass), Lolium multiflorum (Italian ryegrass) and Lolium temulentum (darnel) plants produced by microprojectile bombardment. Plant Cell Rep. 1999, 18, 721–726. [Google Scholar] [CrossRef]
- Altpeter, F.; Xu, J.; Ahmed, S. Generation of large numbers of independently transformed fertile perennial ryegrass (Lolium perenne L.) plants of forage- and turf-type cultivars. Mol. Breed. 2000, 6, 519–528. [Google Scholar] [CrossRef]
- Xu, J.; Schubert, J.; Altpeter, F. Dissection of RNA-mediated ryegrass mosaic virus resistance in fertile transgenic perennial ryegrass (Lolium perenne L.). Plant J. 2001, 26, 265–274. [Google Scholar] [CrossRef] [Green Version]
- Altpeter, F.; Fang, Y.-D.; Xu, J.; Ma, X. Comparison of transgene expression stability after Agrobacterium-mediated or biolistic gene transfer into perennial ryegrass (Lolium perenne L.). In Molecular Breeding of Forage and Turf; Springer: Berlin/Heidelberg, Germany, 2004; pp. 255–260. [Google Scholar]
- Hisano, H.; Kanazawa, A.; Kawakami, A.; Yoshida, M.; Shimamoto, Y.; Yamada, T. Transgenic perennial ryegrass plants expressing wheat fructosyltransferase genes accumulate increased amounts of fructan and acquire increased tolerance on a cellular level to freezing. Plant Sci. 2004, 167, 861–868. [Google Scholar] [CrossRef] [Green Version]
- Petrovska, N.; Wu, X.; Donato, R.; Wang, Z.; Ong, E.-K.; Jones, E.; Forster, J.; Emmerling, M.; Sidoli, A.; O’Hehir, R.; et al. Transgenic ryegrasses (Lolium spp.) with down-regulation of main pollen allergens. Mol. Breed. 2004, 14, 489–501. [Google Scholar] [CrossRef]
- Chen, X.; Yang, W.; Sivamani, E.; Bruneau, A.H.; Wang, B.; Qu, R. Selective elimination of perennial ryegrass by activation of a pro-herbicide through engineering E. coli argE gene. Mol. Breed. 2005, 15, 339–347. [Google Scholar] [CrossRef]
- Wu, Y.-Y.; Chen, Q.-J.; Chen, M.; Chen, J.; Wang, X.-C. Salt-tolerant transgenic perennial ryegrass (Lolium perenne L.) obtained by Agrobacterium tumefaciens-mediated transformation of the vacuolar Na+/H+ antiporter gene. Plant Sci. 2005, 169, 65–73. [Google Scholar] [CrossRef]
- Altpeter, F. Perennial ryegrass (Lolium perenne L.). In Agrobacterium Protocols; Springer: Berlin/Heidelberg, Germany, 2006; Volume 2, pp. 55–64. [Google Scholar] [CrossRef]
- Bajaj, S.; Ran, Y.; Phillips, J.; Kularajathevan, G.; Pal, S.; Cohen, D.; Elborough, K.; Puthigae, S. A high throughput Agrobacterium tumefaciens-mediated transformation method for functional genomics of perennial ryegrass (Lolium perenne L.). Plant Cell Rep. 2006, 25, 651–659. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.X.; Huang, J.Q.; He, Y.L.; Liu, S.J.; Wang, C.L.; Jiang, W.Z.; Wei, Z.M. Transformation of recalcitrant turfgrass cultivars through improvement of tissue culture and selection regime. Plant Cell Tissue Organ Cult. 2006, 85, 307–316. [Google Scholar] [CrossRef]
- Petersen, K.; Kolmos, E.; Folling, M.; Salchert, K.; Storgaard, M.; Jensen, C.S.; Didion, T.; Nielsen, K.K. Two MADS-box genes from perennial ryegrass are regulated by vernalization and involved in the floral transition. Physiol. Plant. 2006, 126, 268–278. [Google Scholar] [CrossRef]
- Sato, H.; Takamizo, T. Agrobacterium tumefaciens-mediated transformation of forage-type perennial ryegrass (Lolium perenne L.). Grassl. Sci. 2006, 52, 95–98. [Google Scholar] [CrossRef]
- Wu, Y.Y.; Chen, Q.J.; Cui, X.H.; Chen, H.; Chen, J.; Wang, X.C. Efficient regeneration and Agrobacterium-mediated stable transformation of perennial ryegrass. Russ. J. Plant Physiol. 2007, 54, 524–529. [Google Scholar] [CrossRef]
- Gadegaard, G.; Didion, T.; Folling, M.; Storgaard, M.; Andersen, C.H.; Nielsen, K.K. Improved fructan accumulation in perennial ryegrass transformed with the onion fructosyltransferase genes 1-SST and 6G-FFT. J. Plant Physiol. 2008, 165, 1214–1225. [Google Scholar] [CrossRef]
- Li, X.; Cheng, X.; Liu, J.; Zeng, H.; Han, L.; Tang, W. Heterologous expression of the Arabidopsis DREB1A/CBF3 gene enhances drought and freezing tolerance in transgenic Lolium perenne plants. Plant Biotechnol. Rep. 2011, 5, 61–69. [Google Scholar] [CrossRef]
- Wu, J.; Zhang, Z.; Zhang, Q.; Lang, Z.; Sun, X. Scarabaeid larvae-and herbicide-resistant transgenic perennial ryegrass (Lolium perenne L.) obtained by Agrobacterium tumefaciens-mediated transformation of cry8Ca2, cry8Ga and bar genes. J. Integr. Agric. 2012, 11, 53–61. [Google Scholar] [CrossRef]
- Patel, M.; Dewey, R.E.; Qu, R. Enhancing Agrobacterium tumefaciens-mediated transformation efficiency of perennial ryegrass and rice using heat and high maltose treatments during bacterial infection. Plant Cell Tissue Organ Cult. 2013, 114, 19–29. [Google Scholar] [CrossRef]
- Zhang, W.-J.; Dewey, R.E.; Boss, W.; Phillippy, B.Q.; Qu, R. Enhanced Agrobacterium-mediated transformation efficiencies in monocot cells is associated with attenuated defense responses. Plant Mol. Biol. 2013, 81, 273–286. [Google Scholar] [CrossRef] [PubMed]
- Vail, A.W.; Wang, P.; Uefuji, H.; Samac, D.A.; Vance, C.P.; Wackett, L.P.; Sadowsky, M.J. Biodegradation of atrazine by three transgenic grasses and alfalfa expressing a modified bacterial atrazine chlorohydrolase gene. Transgenic Res. 2015, 24, 475–488. [Google Scholar] [CrossRef]
- Cen, H.; Ye, W.; Liu, Y.; Li, D.; Wang, K.; Zhang, W. Overexpression of a chimeric gene, OsDST-SRDX, improved salt tolerance of perennial ryegrass. Sci. Rep. 2016, 6, 27320. [Google Scholar] [CrossRef]
- Panter, S.; Mouradov, A.; Badenhorst, P.; Martelotto, L.; Griffith, M.; Smith, K.F.; Spangenberg, G. Re-programming photosynthetic cells of perennial ryegrass (Lolium perenne L) for fructan biosynthesis through transgenic expression of fructan biosynthetic genes under the control of photosynthetic promoters. Agronomy 2017, 7, 36. [Google Scholar] [CrossRef] [Green Version]
- Badenhorst, P.E.; Panter, S.; Palanisamy, R.; Georges, S.; Smith, K.F.; Mouradov, A.; Mason, J.; Spangenberg, G.C. Molecular breeding of transgenic perennial ryegrass (Lolium perenne L.) with altered fructan biosynthesis through the expression of fructosyltransferases. Mol. Breed. 2018, 38, 21. [Google Scholar] [CrossRef]
- Xu, B.; Yu, G.; Li, H.; Xie, Z.; Wen, W.; Zhang, J.; Huang, B. Knockdown of STAYGREEN in perennial ryegrass (Lolium perenne L.) leads to transcriptomic alterations related to suppressed leaf senescence and improved forage quality. Plant Cell Physiol. 2019, 60, 202–212. [Google Scholar] [CrossRef]
- Esmaeili, S.; Salehi, H.; Khosh-Khui, M.; Niazi, A.; Tohidfar, M.; Aram, F. Isopentenyl transferase (IPT) gene transfer to perennial ryegrass through sonication-assisted Agrobacterium-mediated transformation (SAAT), vacuum and heat treatment. Mol. Biotechnol. 2019, 61, 332–344. [Google Scholar] [CrossRef]
- Dalton, S.J. A reformulation of Murashige and Skoog medium (WPBS medium) improves embryogenesis, morphogenesis and transformation efficiency in temperate and tropical grasses and cereals. Plant Cell Tissue Organ Cult. 2020, 141, 257. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Ran, Y.; Nagy, I.; Lenk, I.; Qiu, J.L.; Asp, T.; Jensen, C.S.; Gao, C. Targeted mutagenesis in ryegrass (Lolium spp.) using the CRISPR/Cas9 system. Plant Biotechnol. J. 2020, 18, 1854–1856. [Google Scholar] [CrossRef] [Green Version]
- Cooney, L.J.; Beechey-Gradwell, Z.; Winichayakul, S.; Richardson, K.A.; Crowther, T.; Anderson, P.; Scott, R.W.; Bryan, G.; Roberts, N.J. Changes in leaf-level nitrogen partitioning and mesophyll conductance deliver increased photosynthesis for Lolium perenne leaves engineered to accumulate lipid carbon sinks. Front. Plant Sci. 2021, 12, 330. [Google Scholar] [CrossRef] [PubMed]
- Hang, N.; Shi, T.; Liu, Y.; Ye, W.; Taier, G.; Sun, Y.; Wang, K.; Zhang, W. Overexpression of Os-microRNA408 enhances drought tolerance in perennial ryegrass. Physiol. Plant. 2021, 172, 733–747. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Xie, Z.; Zhang, J.; Lei, S.; Lin, W.; Xu, B.; Huang, B. NOL-mediated functional stay-green traits in perennial ryegrass (Lolium perenne L.) involving multifaceted molecular factors and metabolic pathways regulating leaf senescence. Plant J. 2021, 106, 1219–1232. [Google Scholar] [CrossRef] [PubMed]
- Beechey-Gradwell, Z.; Kadam, S.; Bryan, G.; Cooney, L.; Nelson, K.; Richardson, K.; Cookson, R.; Winichayakul, S.; Reid, M.; Anderson, P.; et al. Lolium perenne engineered for elevated leaf lipids exhibits greater energy density in field canopies under defoliation. Field Crops Res. 2022, 275, 108340. [Google Scholar] [CrossRef]
- Shrawat, A.K.; Lörz, H. Agrobacterium-mediated transformation of cereals: A promising approach crossing barriers. Plant Biotechnol. J. 2006, 4, 575–603. [Google Scholar] [CrossRef]
- Wang, Z.-Y.; Ge, Y. Recent advances in genetic transformation of forage and turf grasses. Vitr. Cell. Dev. Biol.-Plant 2006, 42, 1–18. [Google Scholar] [CrossRef]
- Gelvin, S.B. Agrobacterium-mediated plant transformation: The biology behind the “gene-jockeying” tool. Microbiol. Mol. Biol. Rev. 2003, 67, 16–37. [Google Scholar] [CrossRef] [Green Version]
- Kumlehn, J.; Serazetdinova, L.; Hensel, G.; Becker, D.; Loerz, H. Genetic transformation of barley (Hordeum vulgare L.) via infection of androgenetic pollen cultures with Agrobacterium tumefaciens. Plant Biotechnol. J. 2006, 4, 251–261. [Google Scholar] [CrossRef]
- Manzanares, C.; Barth, S.; Thorogood, D.; Byrne, S.L.; Yates, S.; Czaban, A.; Asp, T.; Yang, B.; Studer, B. A Gene encoding a DUF247 domain protein cosegregates with the S self-incompatibility locus in perennial ryegrass. Mol. Biol. Evol. 2016, 33, 870–884. [Google Scholar] [CrossRef] [Green Version]
- Rohner, M.; Manzanares, C.; Yates, S.; Thorogood, D.; Copetti, D.; Lübberstedt, T.; Asp, T.; Studer, B. Fine-mapping and comparative genomic analysis reveal the gene composition at the S and Z self-incompatibility loci in grasses. bioRxiv 2022. [Google Scholar] [CrossRef]
- Shinozuka, H.; Cogan, N.O.I.; Smith, K.F.; Spangenberg, G.C.; Forster, J.W. Fine-scale comparative genetic and physical mapping supports map-based cloning strategies for the self-incompatibility loci of perennial ryegrass (Lolium perenne L.). Plant Mol. Biol. 2010, 72, 343–355. [Google Scholar] [CrossRef] [PubMed]
- Raissig, M.T.; Matos, J.L.; Gil, M.X.A.; Kornfeld, A.; Bettadapur, A.; Abrash, E.; Allison, H.R.; Badgley, G.; Vogel, J.P.; Berry, J.A.; et al. Mobile MUTE specifies subsidiary cells to build physiologically improved grass stomata. Science 2017, 355, 1215–1218. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.L.-C.; Yoshida, H.; Lurin, C.; Ecker, J.R. Regulation of ethylene gas biosynthesis by the Arabidopsis ETO1 protein. Nature 2004, 428, 945–950. [Google Scholar] [CrossRef]
- Haas, J.H.; Moore, L.W.; Ream, W.; Manulis, S. Universal PCR primers for detection of phytopathogenic Agrobacterium strains. Appl. Environ. Microbiol. 1995, 61, 2879–2884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.M.; Roche, J.R.; Donaghy, D.J.; Thrush, A.; Sathish, P. Validation of reference genes for quantitative RT-PCR studies of gene expression in perennial ryegrass (Lolium perenne L.). BMC Mol. Biol. 2010, 11, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Begheyn, R.F.; Roulund, N.; Vangsgaard, K.; Kopecky, D.; Studer, B. Inheritance patterns of the response to in vitro doubled haploid induction in perennial ryegrass (Lolium perenne L.). Plant Cell Tissue Organ Cult. 2017, 130, 667–679. [Google Scholar] [CrossRef] [Green Version]
- Kumari, M.; Clarke, H.J.; Small, I.; Siddique, K.H.M. Albinism in plants: A major bottleneck in wide hybridization, androgenesis and doubled haploid culture. Crit. Rev. Plant Sci. 2009, 28, 393–409. [Google Scholar] [CrossRef]
- Brew-Appiah, R.A.T.; Ankrah, N.; Liu, W.; Konzak, C.F.; von Wettstein, D.; Rustgi, S. Generation of doubled haploid transgenic wheat lines by microspore transformation. PLoS ONE 2013, 8, e80155. [Google Scholar] [CrossRef]
- Chauhan, H.; Khurana, P. Use of doubled haploid technology for development of stable drought tolerant bread wheat (Triticum aestivum L.) transgenics. Plant Biotechnol. J. 2011, 9, 408–417. [Google Scholar] [CrossRef] [Green Version]
- Valentine, J.; Charles, A.H. Variation in plasticity within the S23 cultivar of Lolium perenne L. J. Agric. Sci. 1975, 85, 111–121. [Google Scholar] [CrossRef]
- Weigel, D.; Glazebrook, J. Transformation of Agrobacterium using the freeze-thaw method. CSH Protoc. 2006, 7. [Google Scholar] [CrossRef] [PubMed]
- Stanke, M.; Morgenstern, B. AUGUSTUS: A web server for gene prediction in eukaryotes that allows user-defined constraints. Nucleic Acids Res. 2005, 33, 465–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byrne, S.L.; Nagy, I.; Pfeifer, M.; Armstead, I.; Swain, S.; Studer, B.; Mayer, K.; Campbell, J.D.; Czaban, A.; Hentrup, S.; et al. A synteny-based draft genome sequence of the forage grass Lolium perenne. Plant J. 2015, 84, 816–826. [Google Scholar] [CrossRef]
- Engler, C.; Gruetzner, R.; Kandzia, R.; Marillonnet, S. Golden gate shuffling: A one-pot DNA shuffling method based on type IIs restriction enzymes. PLoS ONE 2009, 4, e5553. [Google Scholar] [CrossRef] [Green Version]
- Engler, C.; Youles, M.; Gruetzner, R.; Ehnert, T.M.; Werner, S.; Jones, J.D.G.G.; Patron, N.J.; Marillonnet, S. A Golden Gate modular cloning toolbox for plants. ACS Synth. Biol. 2014, 3, 839–843. [Google Scholar] [CrossRef]
- Bull, S.E.; Seung, D.; Chanez, C.; Mehta, D.; Kuon, J.E.; Truernit, E.; Hochmuth, A.; Zurkirchen, I.; Zeeman, S.C.; Gruissem, W.; et al. Accelerated ex situ breeding of GBSS- and PTST1-edited cassava for modified starch. Sci. Adv. 2018, 4, eaat6086. [Google Scholar] [CrossRef] [Green Version]
- Manzanares, C.; Yates, S.; Ruckle, M.; Nay, M.; Studer, B. TILLING in forage grasses for gene discovery and breeding improvement. New Biotechnol. 2016, 33, 594–603. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Doench, J.G.; Fusi, N.; Sullender, M.; Hegde, M.; Vaimberg, E.W.; Donovan, K.F.; Smith, I.; Tothova, Z.; Wilen, C.; Orchard, R.; et al. Optimized sgRNA design to maximize activity and minimize off-target effects of CRISPR-Cas9. Nat. Biotechnol. 2016, 34, 184–191. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Construct ID | A. tumefaciens Strain | Size [bp] 1 | Genotype | Explant for Callus Induction | Number of Calli/Transformation | Regenerated Plants/Events | PCR Positive (Transgenic) Plants/Events | Percentage of Calli Regenerating Transgenic Events |
|---|---|---|---|---|---|---|---|---|
| G150 | LBA4044 | 4564 | S23 Z 2 | shoot tips | 60 | 15/15 | 15/15 | 25.0 |
| G151 | LBA4044 | 4564 | S23 Z 2 | shoot tips | 60 | 19/19 | 19/19 | 31.7 |
| G152 | LBA4044 | 4564 | S23 Z 2 | shoot tips | 60 | 6/6 | 6/6 | 10.0 |
| G153 | LBA4044 | 4564 | S23 Z 2 | shoot tips | 72 | 10/10 | 10/10 | 13.9 |
| G154 | LBA4044 | 4564 | S23 Z 2 | shoot tips | 60 | 16/16 | 16/16 | 26.7 |
| G155 | LBA4044 | 4564 | S23 Z 2 | shoot tips | 72 | 23/23 | 23/23 | 31.9 |
| G156 | LBA4044 | 4564 | S23 Z 2 | shoot tips | 60 | 12/12 | 12/12 | 20.0 |
| G157 | LBA4044 | 4564 | S23 Z 2 | shoot tips | 72 | 22/22 | 22/22 | 30.6 |
| G158 | LBA4044 | 4564 | S23 Z 2 | shoot tips | 60 | 12/12 | 12/12 | 20.0 |
| G159 | LBA4044 | 4564 | S23 Z 2 | shoot tips | 72 | 14/14 | 14/14 | 19.4 |
| G160 | LBA4044 | 6349 | S23 Z 2 | shoot tips | 36 | 20/20 | 20/20 | 55.6 |
| G161 | LBA4044 | 6335 | S23 Z 2 | shoot tips | 36 | 14/14 | 14/14 | 38.9 |
| G162 | LBA4044 | 6362 | S23 Z 2 | shoot tips | 36 | 4/4 | 4/4 | 11.1 |
| G163 | LBA4044 | 6308 | S23 Z 2 | shoot tips | 36 | 8/8 | 8/8 | 22.2 |
| G164 | LBA4044 | 6280 | S23 Z 2 | shoot tips | 36 | 15/15 | 15/15 | 41.7 |
| G166 | LBA4044 | 5593 | S23 Z 2 | shoot tips | 36 | 3/3 | 3/3 | 8.3 |
| C801_GFP | GV3101::pMP90RK | 5415 | S23 Z 2 | shoot tips | 36 | 11/11 | 11/11 | 30.6 |
| C801_GFP | GV3101::pMP90RK | 5415 | Arolus 3 | seeds | 24 | 3/2 | 1/1 | 4.2 |
| B330_MUTE_2 | GV3101::pMP90RK | 9446 | Arolus 3 | seeds | 192 | 83/42 | 70/36 | 18.8 |
| B330_ETO1_807 | GV3101::pMP90RK | 9446 | 6–10 4 | anthers | 144 5 | 10/2 | 9/1 5 | 0.7 |
| Total | 1260 | 320/270 | 304/262 | 20.8 |
| Construct ID | A. tumefaciens Strain | Insert | Backbone | Selectable Marker | Cloning Method |
|---|---|---|---|---|---|
| G150 | LBA4044 | p35S::dsRNAbp-hp::t35S | pAGM4673 | kanamycin | Golden Gate |
| G151 | LBA4044 | plolp2::dsRNAbp-hp::t35S | pAGM4673 | kanamycin | Golden Gate |
| G152 | LBA4044 | p35S::SDUF247-I-hp::t35S | pAGM4673 | kanamycin | Golden Gate |
| G153 | LBA4044 | plolp2::SDUF247-I-hp::35S | pAGM4673 | kanamycin | Golden Gate |
| G154 | LBA4044 | p35S::SDUF247-II-hp::t35S | pAGM4673 | kanamycin | Golden Gate |
| G155 | LBA4044 | plolp2::SDUF247-II-hp::t35S | pAGM4673 | kanamycin | Golden Gate |
| G156 | LBA4044 | p35S::ZDUF247-I-hp::t35S | pAGM4673 | kanamycin | Golden Gate |
| G157 | LBA4044 | plolp2::ZDUF247-I-hp::t35S | pAGM4673 | kanamycin | Golden Gate |
| G158 | LBA4044 | p35S::ZDUF247-II-hp::t35S | pAGM4673 | kanamycin | Golden Gate |
| G159 | LBA4044 | plolp2::ZDUF247-II-hp::t35S | pAGM4673 | kanamycin | Golden Gate |
| G160 | LBA4044 | pdsRNAbp::GUS::tNOS | pCambia1305.1 | kanamycin | restriction enzyme |
| G161 | LBA4044 | pSDUF247-I::GUS::tNOS | pCambia1305.1 | kanamycin | restriction enzyme |
| G162 | LBA4044 | pSDUF247-II::GUS::tNOS | pCambia1305.1 | kanamycin | restriction enzyme |
| G163 | LBA4044 | pZDUF247-I::GUS::tNOS | pCambia1305.1 | kanamycin | restriction enzyme |
| G164 | LBA4044 | pZDUF247-II::GUS::tNOS | pCambia1305.1 | kanamycin | restriction enzyme |
| G166 | LBA4044 | p35S::GUS::tNOS | pCambia1305.1 | kanamycin | none |
| C801_GFP | GV3101::pMP90RK | pZmUbi-1::GFP::tOCS | C801p6o2x35s-pZmUbi-1-GFP | spectinomycin | none |
| B330_MUTE_2 | GV3101::pMP90RK | pOsU6::MUTE_2sgRNA | B330p6i2xoR-UcasW-pOsU6 | spectinomycin | restriction enzyme |
| B330_ETO1_807 | GV3101::pMP90RK | pOsU6::ETO1_807sgRNA | B330p6i2xoR-UcasW-pOsU6 | spectinomycin | restriction enzyme |
| Gene Name | Gene Annotation i n Lolium multiflorum 1 | Gene Orthologs in Oryza sativa subsp. Japonica | Gene Description NCBI |
|---|---|---|---|
| dsRNAbp | Lmu01_818G00001402 & Lmu01_1212G0000450 | Os05g0150400 | Double-stranded RNA-binding protein 2 |
| SDUF247-I | Lmu01_818G00001902 & Lmu01_1212G0000400 | Os05g0198100 | DUF247; Plant protein of unknown function |
| SDUF247-II | Lmu01_818G0000210 2 & Scf3448 4724980..4726635 3 | Os05g0197900 | DUF247; Plant protein of unknown function |
| ZDUF247-I | Lmu01_1905G0001490 & Lmu01_3448G0000640 2 | Os04g0647425 | DUF247; Plant protein of unknown function |
| ZDUF247-II | Lmu01_1905G00015002 & Scf34481166016..1167647 3 | not annotated | DUF247; Plant protein of unknown function |
| MUTE | Lmu01_7136G0001450 & Lmu01_2560G0000140 | Os03g0294700 | Ethylene-overproduction protein1 |
| ETO1 | Lmu01_1530G0000400 & Lmu01_16G0000470 | Os05g0597000 | DNA binding protein |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grogg, D.; Rohner, M.; Yates, S.; Manzanares, C.; Bull, S.E.; Dalton, S.; Bosch, M.; Studer, B.; Broggini, G.A.L. Callus Induction from Diverse Explants and Genotypes Enables Robust Transformation of Perennial Ryegrass (Lolium perenne L.). Plants 2022, 11, 2054. https://doi.org/10.3390/plants11152054
Grogg D, Rohner M, Yates S, Manzanares C, Bull SE, Dalton S, Bosch M, Studer B, Broggini GAL. Callus Induction from Diverse Explants and Genotypes Enables Robust Transformation of Perennial Ryegrass (Lolium perenne L.). Plants. 2022; 11(15):2054. https://doi.org/10.3390/plants11152054
Chicago/Turabian StyleGrogg, Daniel, Marius Rohner, Steven Yates, Chloe Manzanares, Simon E. Bull, Sue Dalton, Maurice Bosch, Bruno Studer, and Giovanni A. L. Broggini. 2022. "Callus Induction from Diverse Explants and Genotypes Enables Robust Transformation of Perennial Ryegrass (Lolium perenne L.)" Plants 11, no. 15: 2054. https://doi.org/10.3390/plants11152054
APA StyleGrogg, D., Rohner, M., Yates, S., Manzanares, C., Bull, S. E., Dalton, S., Bosch, M., Studer, B., & Broggini, G. A. L. (2022). Callus Induction from Diverse Explants and Genotypes Enables Robust Transformation of Perennial Ryegrass (Lolium perenne L.). Plants, 11(15), 2054. https://doi.org/10.3390/plants11152054