
Precise Identification of Chromosome Constitution and Rearrangements in Wheat–Thinopyrum intermedium Derivatives by ND-FISH and Oligo-FISH Painting



Abstract
:1. Introduction
2. Results
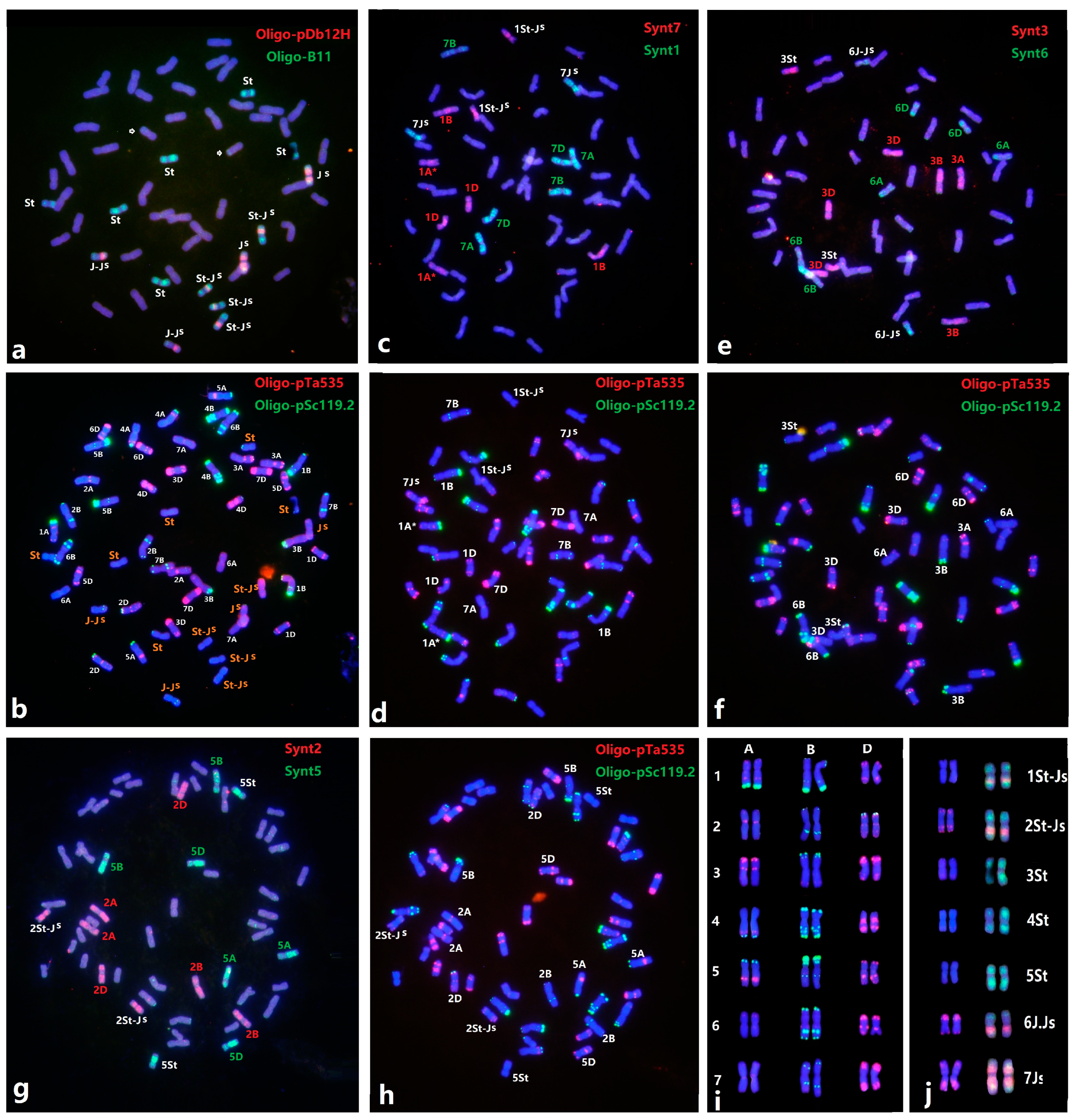
2.1. Identification of Individual Thinopyrum intermedium Chromosomes
2.2. Chromosome Identification of Wheat–Th. intermedium Amphiploids
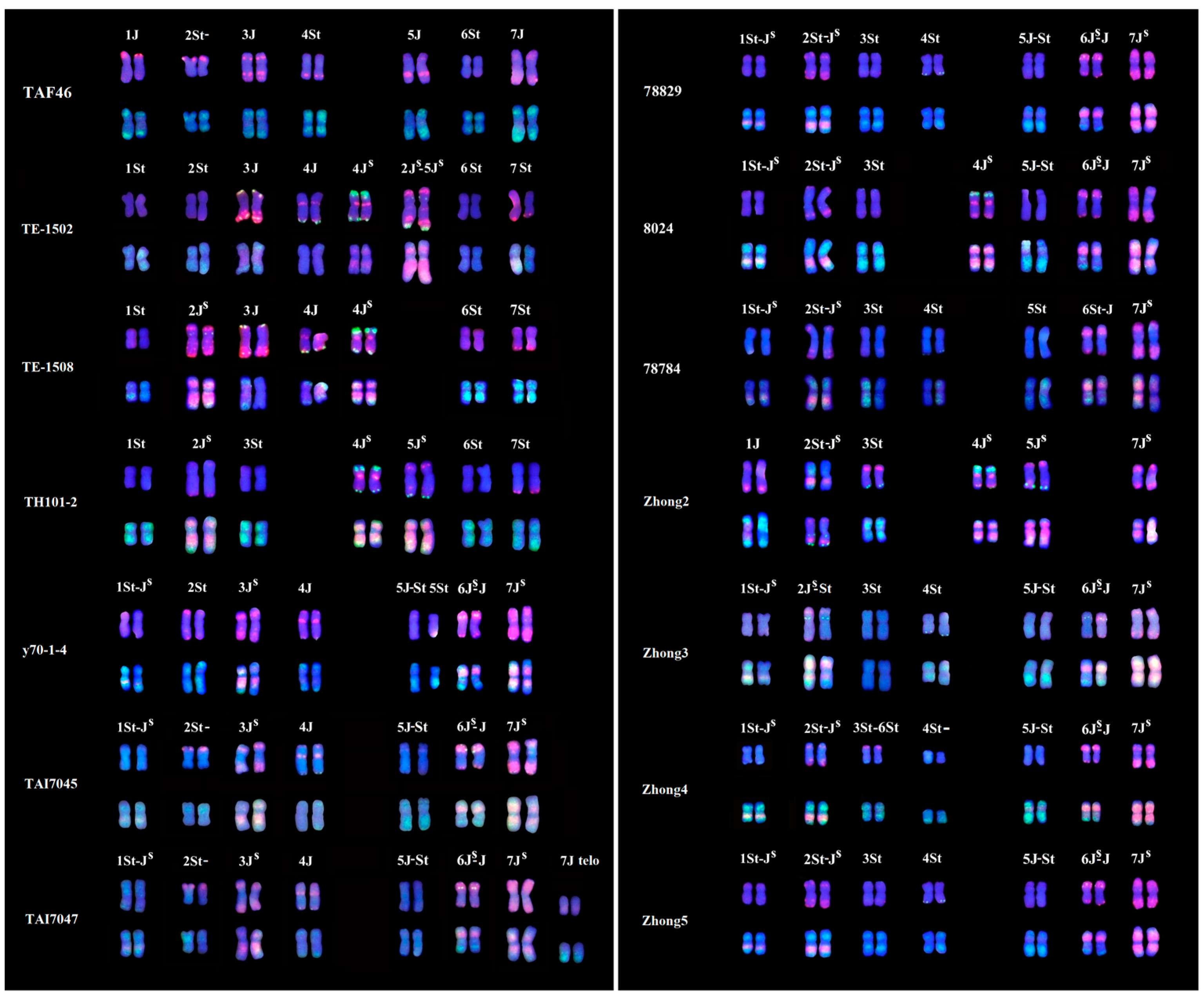
2.3. Constitution of Th. intermedium Chromosomes in 12 Wheat–Thinopyrum Amphiploids
2.4. Revisiting the Karyotype of Wheat–Th. intermedium Additions Z3
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Probe Preparation
4.3. Fluorescence In Situ Hybridization
4.4. Molecular Marker Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tsvelev, N.N. Grasses of the Soviet Union; Oxonian Press Pvt. Ltd.: New Delhi, India, 1983; pp. 196–298. [Google Scholar]
- Jensen, K.B.; Yan, X.B.; Larson, S.R.; Wang, R.R.C.; Robins, J.G. Agronomic and genetic diversity in intermediate wheatgrass (Thinopyrum intermedium). Plant Breed. 2016, 135, 751–758. [Google Scholar] [CrossRef]
- Chen, Q.; Conner, R.L.; Laroche, A.; Thomas, J.B. Genome analysis of Thinopyrum intermedium and Thinopyrum ponticum using genomic in situ hybridization. Genome 1998, 41, 580–586. [Google Scholar] [CrossRef]
- Chen, Q. Detection of alien chromatin introgression from Thinopyrum into wheat using S genomic DNA as a probe—A landmark approach for Thinopyrum genome research. Cytogenet. Genome Res. 2005, 109, 350–359. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, X. Thinopyrum ponticum and Th. intermedium: The promising source of resistance to fungal and viral diseases of wheat. J. Genet. Genom. 2009, 36, 557–565. [Google Scholar] [CrossRef]
- Chang, Z.J.; Zhang, X.J.; Yang, Z.J.; Zhan, H.X.; Li, X.; Liu, C.; Zhang, C.Z. Characterization of a partial wheat—Thinopyrum intermedium amphiploid and its reaction to fungal diseases of wheat. Hereditas 2010, 147, 304–312. [Google Scholar] [CrossRef]
- Fedak, G.; Han, F. Characterization of derivatives from wheat—Thinopyrum wide crosses. Cytogenet. Genom. Res. 2005, 109, 360–367. [Google Scholar] [CrossRef]
- Bao, Y.G.; Wu, X.; Zhang, C.; Li, X.F.; Han, F.P.; Qi, X.L.; Wang, H.G. Chromosomal constitutions and reactions to powdery mildew and stripe rust of four novel wheat—Thinopyrum intermedium partial amphiploids. J. Genet. Genom. 2014, 41, 663–666. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Xing, P.; Qi, X.; Bao, Y.; Wang, H.; Wang, R.R.C.; Li, X. Characterization of chromosome constitution in three wheat—Thinopyrum intermedium amphiploids revealed frequent rearrangement of alien and wheat chromosomes. BMC Plant Biol. 2021, 21, 129. [Google Scholar] [CrossRef]
- Mahelka, V.; Kopecky, D.; Baum, B.R. Contrasting patterns of evolution of 45S and 5S rDNA families uncover new aspects in the genome constitution of the agronomically important grass Thinopyrum intermedium (Triticeae). Mol. Biol. Evol. 2013, 30, 2065–2086. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chen, Q.; Zhang, P.; Lang, T.; Hoxha, S.; Li, G.; Yang, Z. Comparative FISH and molecular identification of new stripe rust resistant wheat—Thinopyrum intermedium ssp. trichophorum introgression lines. Crop J. 2019, 7, 819–829. [Google Scholar] [CrossRef]
- Cui, L.; Ren, Y.K.; Zhang, Y.M.; Tang, Z.H.; Guo, Q.; Niu, Y.Q.; Yan, W.Z.; Sun, Y.; Li, H.J. Characterization of resistance to cereal cyst nematode, agronomic performance, and end-use quality parameters in four perennial wheat—Thinopyrum intermedium lines. Front. Plant Sci. 2020, 11, 594197. [Google Scholar] [CrossRef]
- Li, G.; Zhang, T.; Yu, Z.; Wang, H.; Yang, E.; Yang, Z. An efficient Oligo-FISH painting system for revealing chromosome rearrangements and polyploidization in Triticeae. Plant J. 2021, 105, 978–993. [Google Scholar] [CrossRef]
- Li, G.; Yang, Z. Oligo-FISH paints in Triticeae. Curr. Protoc. 2022, 2, e364. [Google Scholar] [CrossRef]
- Tang, S.; Tang, Z.; Qiu, L.; Yang, Z.; Li, G.; Lang, T.; Zhu, W.; Zhang, J.; Fu, S. Developing new oligo probes to distinguish specific chromosomal segments and the A, B, D genomes of wheat (Triticum aestivum L.) using ND-FISH. Front. Plant Sci. 2018, 9, 1104. [Google Scholar] [CrossRef]
- Tang, Z.; Yang, Z.; Fu, S. Oligonucleotides replacing the roles of repetitive sequences pAs1, pSc119.2, pTa-535, pTa71, CCS1, and pAWRC.1 for FISH analysis. J. Appl. Genet. 2014, 5, 313–318. [Google Scholar] [CrossRef]
- Xi, W.; Tang, Z.; Tang, S.; Yang, Z.; Luo, J.; Fu, S. New ND-FISH-positive Oligo probes for identifying Thinopyrum chromosomes in wheat backgrounds. Int. J. Mol. Sci. 2019, 20, 2031. [Google Scholar] [CrossRef]
- Yu, Z.; Wang, H.; Xu, Y.; Li, Y.; Lang, T.; Yang, Z.; Li, G. Characterization of chromosomal rearrangement in new wheat—Thinopyrum intermedium addition lines carrying Thinopyrum-specific grain hardness genes. Agronomy 2019, 9, 18. [Google Scholar] [CrossRef]
- Fu, S.L.; Chen, L.; Wang, Y.; Li, M.; Yang, Z.; Qiu, L.; Yan, B.; Ren, Z.; Tang, Z. Oligonucleotide probes for ND-FISH analysis to identify rye and wheat chromosomes. Sci. Rep. 2015, 5, 10552. [Google Scholar] [CrossRef]
- Lang, T.; Li, G.R.; Wang, H.J.; Yu, Z.H.; Chen, Q.H.; Yang, E.N.; Fu, S.L.; Tang, Z.X.; Yang, Z.J. Physical location of tandem repeats in the wheat genome and application for chromosome identification. Planta 2019, 249, 663–675. [Google Scholar] [CrossRef]
- Chen, Q.; Conner, R.L.; Li, H.J.; Sun, S.C.; Ahmad, F.; Laroche, A.; Graf, R.J. Molecular cytogenetic discrimination and reaction to wheat streak mosaic virus and the wheat curl mite in Zhong series of wheat—Thinopyrum intermedium partial amphiploids. Genome 2003, 46, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.M.; Gill, B.S. Nonisotopic in situ hybridization and plant genome mapping: The first 10 years. Genome 1994, 37, 717–725. [Google Scholar] [CrossRef]
- Jiang, J.; Friebe, B.; Gill, B.S. Recent advances in alien gene transfer in wheat. Euphytica 1994, 73, 199–212. [Google Scholar] [CrossRef]
- Friebe, B.; Mukai, Y.; Gill, B.S.; Cauderon, Y. C-banding and in-situ hybridization analyses of Agropyron intermedium, a partial wheat x Ag. intermedium amphiploid, and six derived chromosome addition lines. Theor. Appl. Genet. 1992, 84, 899–905. [Google Scholar] [CrossRef] [PubMed]
- Larkin, P.J.; Banks, P.M.; Lagudah, E.S.; Appels, R.; Chen, X.; Xin, Z.Y.; Ohm, H.W.; McIntosh, R.A. Disomic Thinopyrum intermedium addition lines in wheat with barley yellow dwarf virus resistance and with rust resistances. Genome 1996, 38, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Lukaszewski, A.J.; Gustafson, J.P. Translocations and modifications of chromosomes in triticale × wheat hybrids. Theor. Appl. Genet. 1983, 64, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Gill, B.S.; Friebe, B.; Endo, T.R. Standard karyotype and nomenclature system for description of chromosome bands and structural aberrations in wheat (Triticum aestivum L.). Genome 1991, 34, 830–839. [Google Scholar] [CrossRef]
- Mukai, Y.; Gill, B.S. Detection of barley chromatin added to wheat by genomic in situ hybridization. Genome 1991, 34, 448–452. [Google Scholar] [CrossRef]
- Han, Y.H.; Zhang, T.; Thammapichai, P.; Weng, Y.Q.; Jiang, J.M. Chromosome-specific painting in Cucumis species using bulked oligonucleotides. Genetics 2015, 200, 771–779. [Google Scholar] [CrossRef]
- Jiang, J. Fluorescence in situ hybridization in plants: Recent developments and future applications. Chromosome Res. 2019, 27, 153–165. [Google Scholar] [CrossRef]
- Cauderon, Y. Genome analysis in the genus. Agropyron. Hered. 1966, 2, 218–234. [Google Scholar]
- Chen, Q.; Conner, R.L.; Laroche, A.; Ji, W.Q.; Armstrong, K.C.; Fedak, G. Genomic in situ hybridization analysis of Thinopyrum chromatin in a wheat—Th. intermedium partial amphiploid and six derived chromosome addition lines. Genome 1999, 42, 1217–1223. [Google Scholar] [CrossRef]
- Saigne, B.; Dauge, M. The resistance to wheat rusts of Agropyron intermedium and its use in wheat improvement. Proc. Int. Wheat Genet. Symp. 1973, 4, 401–407. [Google Scholar]
- Tang, S.; Li, Z.; Jia, X.; Larkin, P.J. Genomic in situ hybridization (GISH) analyses of Thinopyrum intermedium, its partial amphiploid Zhong 5, and disease-resistant derivatives in wheat. Theor. Appl. Genet. 2000, 100, 344–352. [Google Scholar] [CrossRef]
- Lang, T.; La, S.; Li, B.; Yu, Z.; Chen, Q.; Li, J.; Yang, E.; Li, G.; Yang, Z. Precise identification of wheat-Thinopyrum intermedium translocation chromosomes carrying resistance to wheat stripe rust in line Z4 and its derived progenies. Genome 2018, 61, 177–185. [Google Scholar] [CrossRef]
- Hu, L.J.; Li, G.R.; Zeng, Z.X.; Chang, Z.J.; Liu, C.; Zhou, J.P.; Yang, Z.J. Molecular cytogenetic identification of a new wheat-Thinopyrum substitution line with stripe rust resistance. Euphytica 2011, 177, 169–177. [Google Scholar] [CrossRef]
- Li, G.R.; Liu, C.; Li, C.H.; Zhao, J.M.; Zhou, L.; Dai, G.; Yang, E.N.; Yang, Z.J. Introgression of a novel Thinopyrum intermedium St-chromosome-specific HMW-GS gene into wheat. Mol. Breed. 2013, 31, 843–853. [Google Scholar] [CrossRef]
- Shewry, P.R.; Halford, N.G.; Lafiandra, D. Genetics of wheat gluten proteins. Adv. Genet. 2003, 49, 111–184. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; DeHaan, L.R.; Higgins, L.; Markowski, T.W.; Wyse, D.L.; Anderson, J.A. New insights into high-molecular-weight glutenin subunits and sub-genomes of the perennial crop Thinopyrum intermedium (Triticeae). J. Cereal Sci. 2014, 59, 203–210. [Google Scholar] [CrossRef]
- Niu, Z.X.; Klindworth, D.L.; Wang, R.R.C.; Jauhar, P.P.; Larkin, P.J.; Xu, S.S. Characterization of HMW glutenin subunits in Thinopyrum intermedium, Th. bessarabicum, Lophopyrum elongatum, Aegilops markgrafii, and their addition lines in wheat. Crop Sci. 2011, 51, 667–677. [Google Scholar] [CrossRef]
- Wang, Y.; Cao, Q.; Zhang, J.; Wang, S.; Chen, C.; Wang, C.; Zhang, H.; Wang, Y.; Ji, W. Cytogenetic analysis and molecular marker development for a new wheat–Thinopyrum ponticum 1Js (1D) disomic substitution line with resistance to stripe rust and powdery mildew. Front. Plant Sci. 2020, 11, 1282. [Google Scholar] [CrossRef]
- Li, M.; Wang, Y.; Liu, X.; Li, X.; Wang, H.; Bao, Y. Molecular cytogenetic identification of a novel wheat–Thinopyrum ponticum 1JS (1B) substitution line resistant to powdery mildew and leaf rust. Front. Plant Sci. 2020, 12, 727734. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fu, J.; Shen, Q.; Yang, D. High-molecular-weight glutenin subunits: Genetics, structures, and relation to end use qualities. Int. J. Mol. Sci. 2020, 22, 184. [Google Scholar] [CrossRef]
- Nikitina, E.; Kuznetsova, V.; Kroupin, P.; Karlov, G.I.; Divashuk, M.G. Development of specific Thinopyrum cytogenetic markers for wheat-wheatgrass hybrids using sequencing and qPCR data. Int. J. Mol. Sci. 2020, 21, 4495. [Google Scholar] [CrossRef]
- Qiao, L.; Liu, S.; Li, J.; Li, S.; Yu, Z.; Liu, C.; Li, X.; Liu, J.; Ren, Y.; Zhang, P.; et al. Development of sequence-tagged site marker set for identification of J, JS and St sub-genomes of Thinopyrum intermedium in wheat background. Front. Plant Sci. 2021, 12, 685216. [Google Scholar] [CrossRef] [PubMed]
- Cseh, A.; Yang, C.Y.; Hubbart-Edwards, S.; Scholefield, D.; Ashling, S.S.; Burridgem, A.J.; Wilkinson, P.A.; King, I.P.; King, J.; Grewal, S. Development and validation of an exome-based SNP marker set for identification of the St, Jr and Jvs genomes of Thinopyrym intermedium in a wheat background. Theor. Appl. Genet. 2019, 132, 1555–1570. [Google Scholar] [CrossRef]
- Han, F.P.; Lamb, J.C.; Birchler, J.A. High frequency of centromere inactivation resulting in stable dicentric chromosomes of maize. Proc. Natl. Acad. Sci. USA 2006, 103, 3238–3243. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, G.; Nakamura, T.; Ashida, T.; Saito, M.; Nasuda, S.; Endo, T.R.; Wu, J.; Matsumoto, T. Localization of anchor loci representing five hundred annotated rice genes to wheat chromosomes using PLUG markers. Theor. Appl. Genet. 2009, 118, 499–514. [Google Scholar] [CrossRef]
- Zhang, X.; Wei, X.; Xiao, J.; Yuan, C.; Wu, Y.; Cao, A.; Xing, L.; Chen, P.; Zhang, S.; Wang, X.; et al. Whole genome development of intron targeting (IT) markers specific for Dasypyrum villosum chromosomes based on next-generation sequencing technology. Mol. Breed. 2017, 37, 115. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Materials | Chromosome Number | Chromosome Constitution | Figure |
|---|---|---|---|
| Zhong2 | 2n = 56, four 6A | 1J, 2St-JS, 3St, 4JS, 5JS, 7JS | Figure S2-9 |
| Zhong3 | 2n = 56 | 1St-JS, 2JS-St, 3St, 4St, 5J-St, 6JS-J, 7JS | Figure S2-10 |
| Zhong4 | 2n = 56 | 1St-JS, 2St-JS, 3St-6St, 4St-, 5J-St, 6JS-J, 7JS | Figure S2-11 |
| Zhong5 | 2n = 56 | 1St-JS, 2St-JS, 3St, 4St, 5J-St, 6JS-J, 7JS | Figure S2-12 |
| TAI7045 | 2n = 56 | 1St-JS, 2St, 3JS, 4J, 5J-St, 6JS-J, 7JS | Figure S2-3 |
| y70-1-4 | 2n = 56 | 1St-JS, 2St, 3JS, 4J, 5J-St (1), 5St (1), 6JS-J, 7JS | Figure S2-8 |
| 78784 | 2n = 56 | 1St-JS, 2St-JS, 3St, 4St, 5St, 6JS-J, 7JS | Figure S2-1 |
| TAI7047 | 2n = 56 + t | 1St-JS, 2St, 3JS, 4J, 5J-St, 6JS-J, 7JS, 7J telo | Figure S2-4 |
| 78829 | 2n = 56 | 1St-JS, 2St-JS, 3St, 4St, 5J-St, 6JS-J, 7JS | Figure 3 |
| TE-1502 | 2n = 56, absence of 4B | 1St, 2St, 3J, 4J, 4JS, 2JS-5JS, 6St, 7St | Figure S2-5 |
| TE-1508 | 2n = 56 | 1St, 2JS, 3J, 4J, 4JS, 6St, 7St | Figure S2-6 |
| TH101-2 | 2n = 56 | 1St, 2JS, 3St, 4JS, 5JS, 6St, 7St | Figure S2-7 |
| 8024 | 2n = 54, absence of 7D | 1St-JS, 2St-JS, 3St, 4JS, 5J-St, 6JS-J, 7JS | Figure S2-2 |
| TAF46 | 2n = 56 | 1J, 2St-, 3J, 4St, 5J, 6St, 7J | Figure 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Z.; Wang, H.; Yang, E.; Li, G.; Yang, Z. Precise Identification of Chromosome Constitution and Rearrangements in Wheat–Thinopyrum intermedium Derivatives by ND-FISH and Oligo-FISH Painting. Plants 2022, 11, 2109. https://doi.org/10.3390/plants11162109
Yu Z, Wang H, Yang E, Li G, Yang Z. Precise Identification of Chromosome Constitution and Rearrangements in Wheat–Thinopyrum intermedium Derivatives by ND-FISH and Oligo-FISH Painting. Plants. 2022; 11(16):2109. https://doi.org/10.3390/plants11162109
Chicago/Turabian StyleYu, Zhihui, Hongjin Wang, Ennian Yang, Guangrong Li, and Zujun Yang. 2022. "Precise Identification of Chromosome Constitution and Rearrangements in Wheat–Thinopyrum intermedium Derivatives by ND-FISH and Oligo-FISH Painting" Plants 11, no. 16: 2109. https://doi.org/10.3390/plants11162109
APA StyleYu, Z., Wang, H., Yang, E., Li, G., & Yang, Z. (2022). Precise Identification of Chromosome Constitution and Rearrangements in Wheat–Thinopyrum intermedium Derivatives by ND-FISH and Oligo-FISH Painting. Plants, 11(16), 2109. https://doi.org/10.3390/plants11162109