
Seed Priming with ZnO and Fe3O4 Nanoparticles Alleviate the Lead Toxicity in Basella alba L. through Reduced Lead Uptake and Regulation of ROS

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
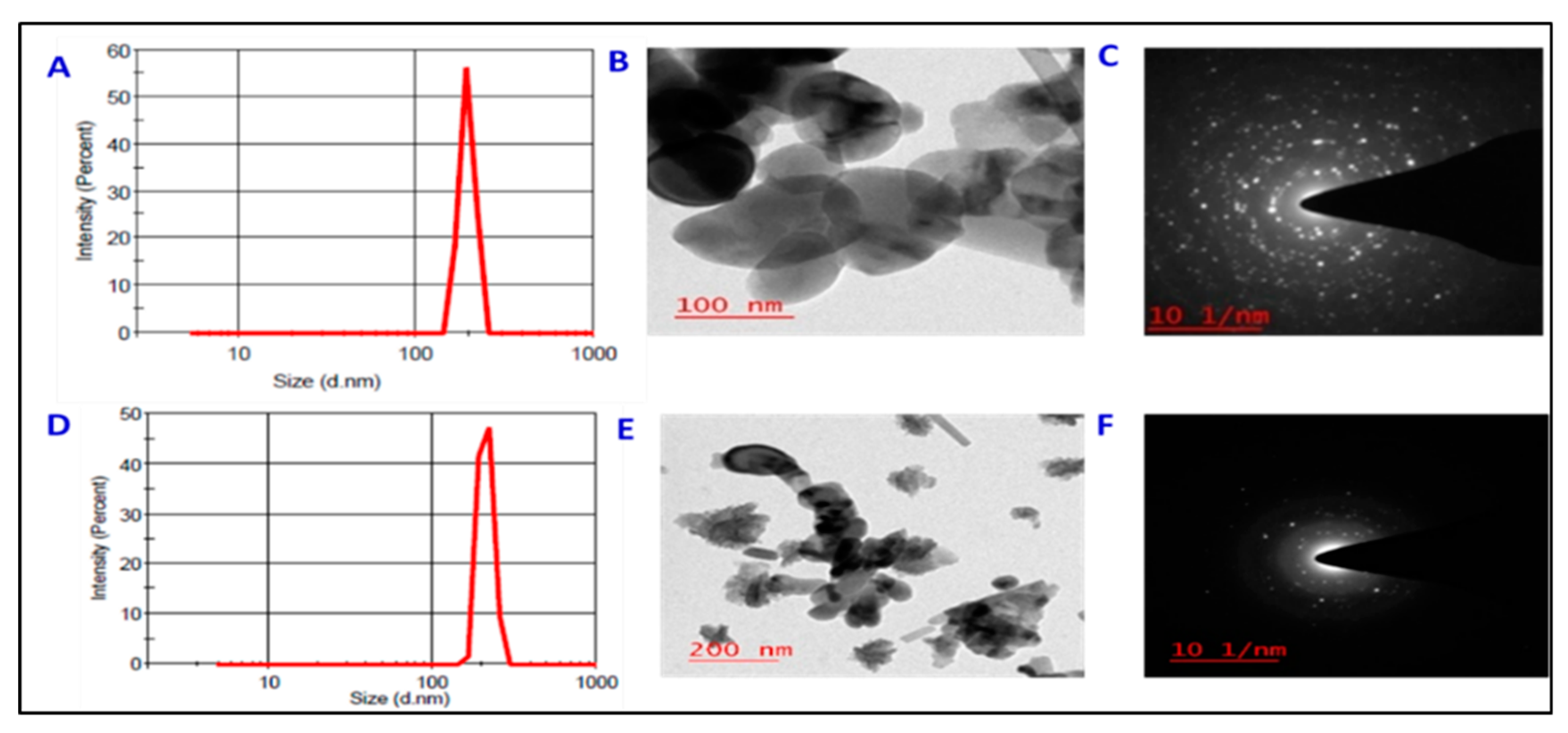
2.1. Synthesis and Characterization of ZnO- and Fe3O4-NPs
- ZnO-NPs
- Fe3O4-NPs
2.2. Preparation of Pb and NPs Solutions and Seed Exposure
2.3. Germination Assay and Seedling Growth Measurement
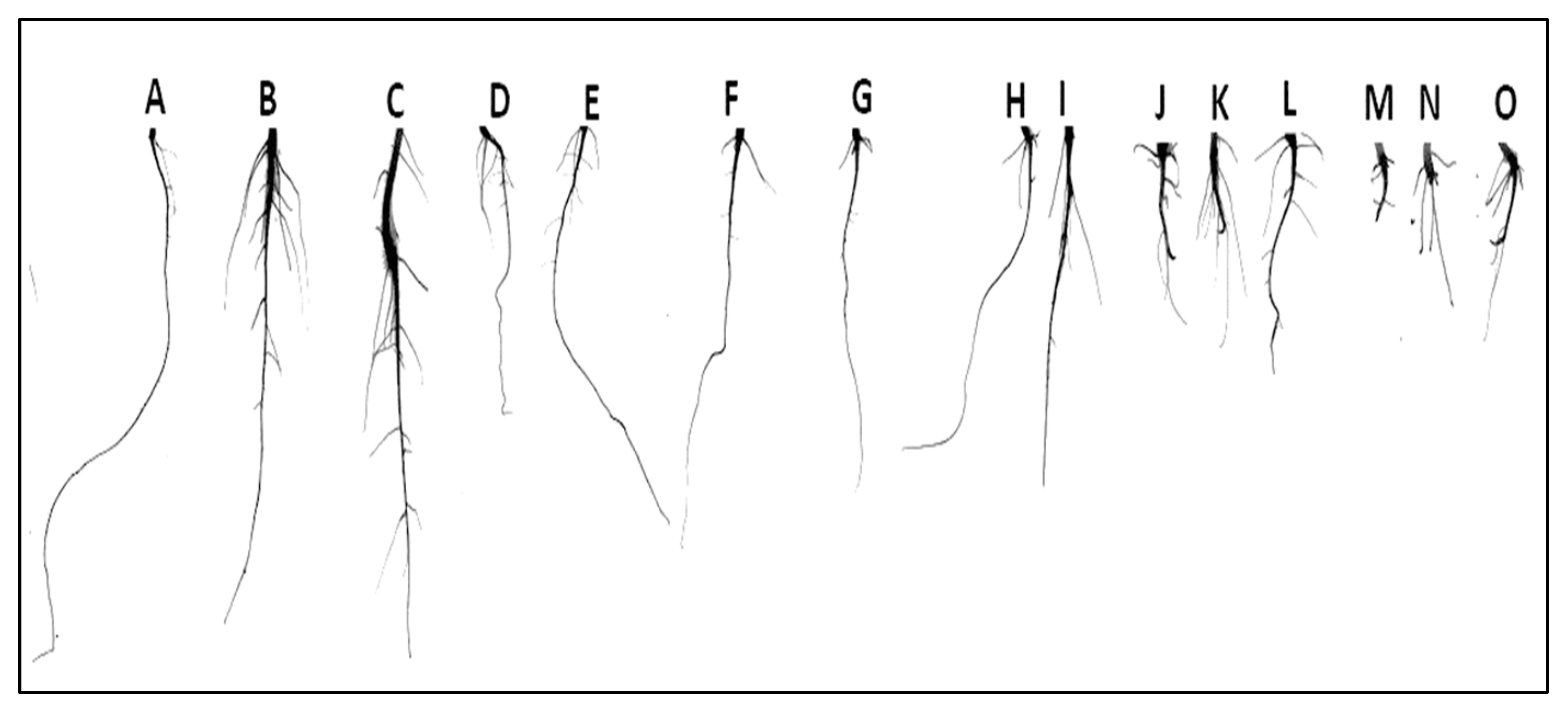
2.4. Root Growth Analysis
2.5. Estimation of Hydrogen Peroxide, Malondialdehyde (MDA) and Proline
2.6. Assay of Antioxidant Enzyme Activities
2.7. Estimation of Lead Concentration
2.8. Computation of NPs Superiority in Alleviating Pb Stress
2.9. Statistical Analysis
3. Results
3.1. Characterization of ZnO- and Fe3O4-NPs
3.2. Preliminary Study with Pb and NPs Treatments
3.3. Effect of NPs Seed Priming under Lead Stressed Conditions
3.4. Effect of NPs Priming on Seed Germination and Seedling Growth under Lead Stress
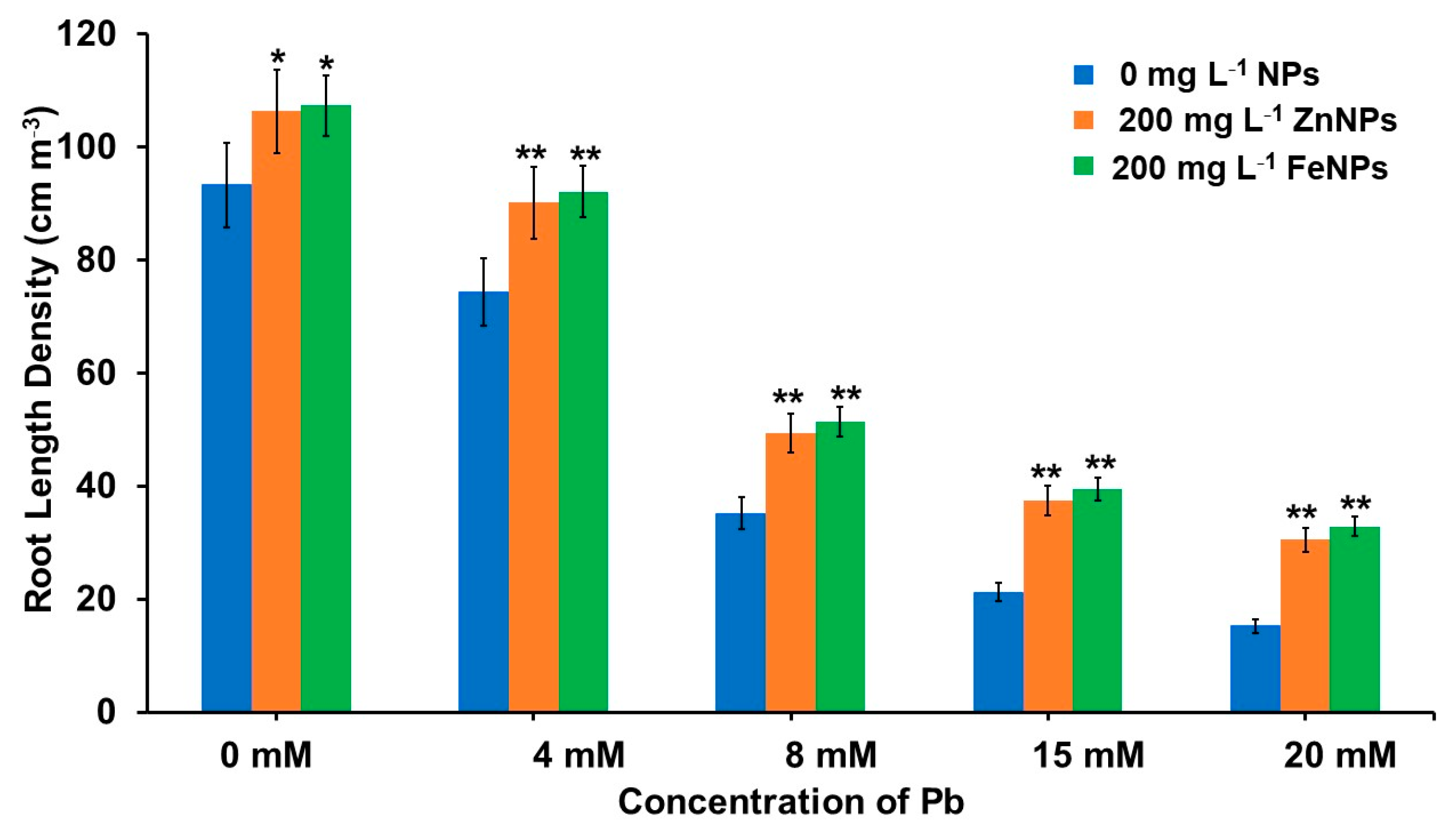
3.5. Effect of Pb and NPs on Root Growth Parameters
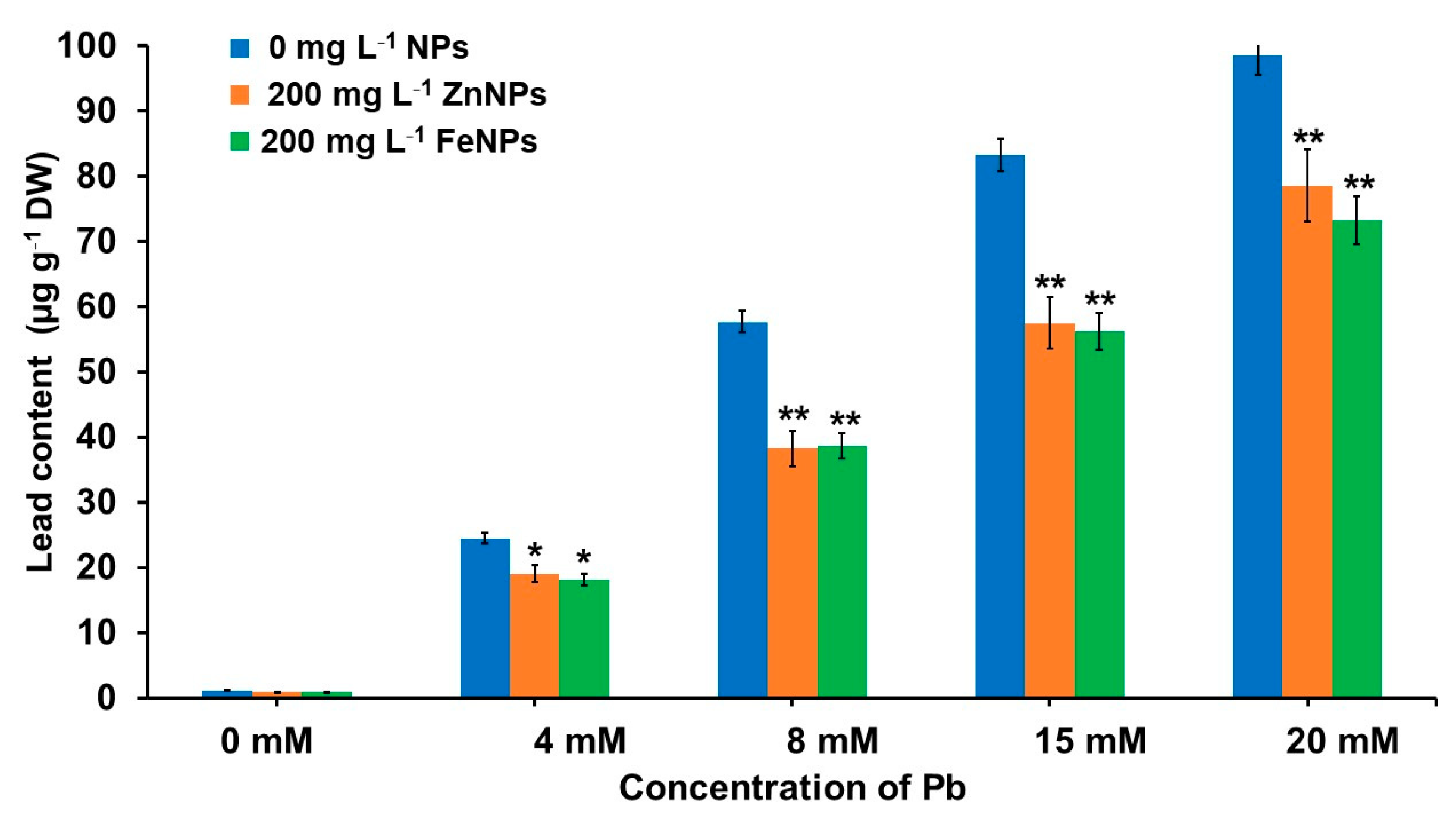
3.6. Effect of NPs Priming on Pb Content under Pb Stress Condition
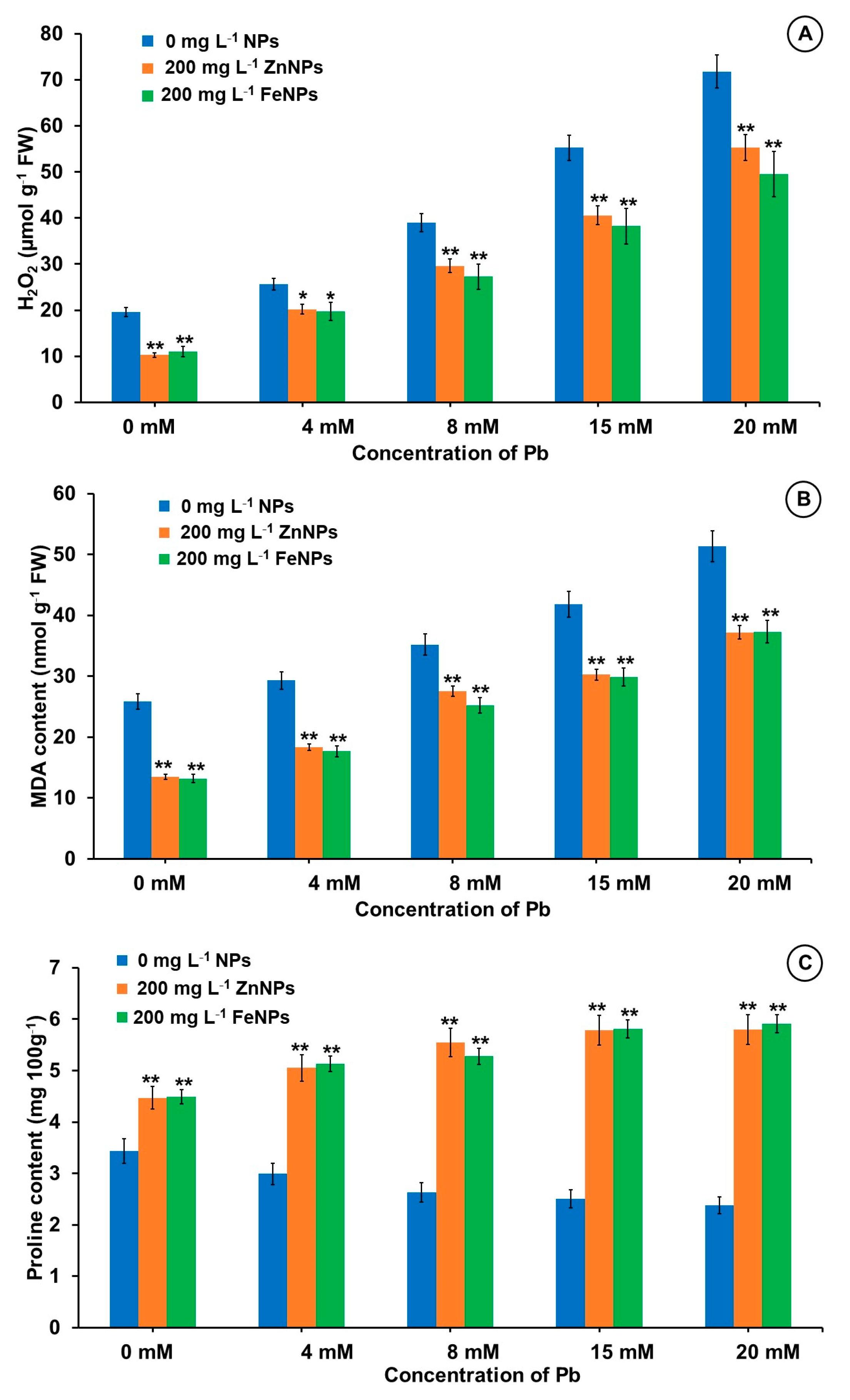
3.7. Effect of NPs Priming on H2O2, MDA and Proline under Pb Stress
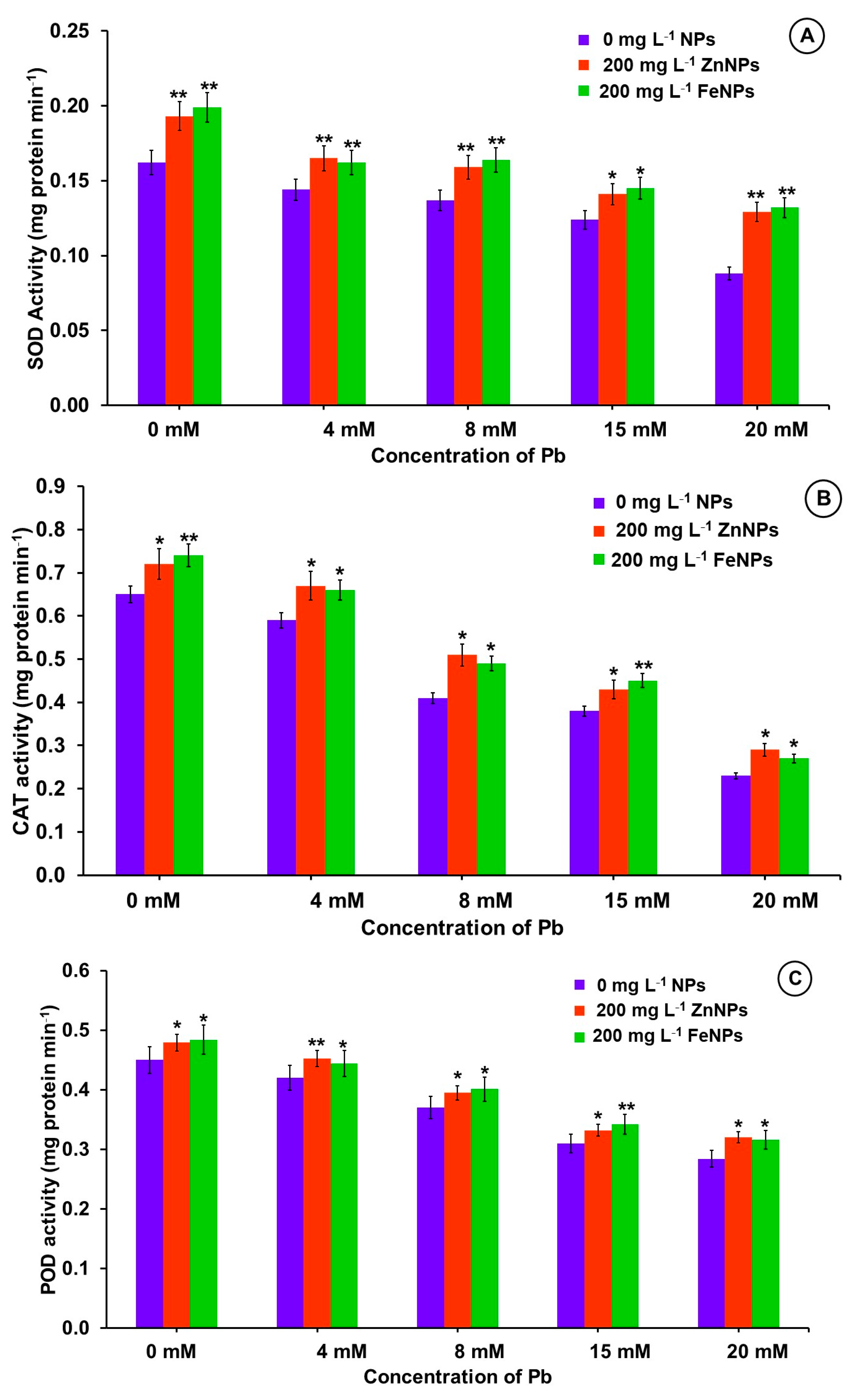
3.8. Effect of NPs Priming on Antioxidant Enzymes under Pb Stress
4. Discussion
4.1. Effect of Lead and NPs Priming on Germination and Seedling Growth
4.2. Effect of NPs Priming on Pb Uptake
4.3. Pb Induced Physio-Biochemical Changes and Its Mitigation by NPs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Costa, R.G.; Araújo, C.F.d.S.; Ferreol Bah, A.H.; Junior, E.A.G.; Rodrigues, Y.J.d.M.; Menezes-Filho, J.A. Lead in mangrove root crab (Goniopsis Cruentata) and risk assessment due to exposure for estuarine villagers. Food Addit. Contam. Part B 2018, 11, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Debnath, B.; Singh, W.; Manna, K. Sources and toxicological effects of lead on human health. Indian J. Med. Spec. 2019, 10, 66. [Google Scholar] [CrossRef]
- Pourrut, B.; Shahid, M.; Dumat, C.; Winterton, P.; Pinelli, E. Lead uptake, toxicity, and detoxification in plants. In Reviews of Environmental Contamination and Toxicology; Springer: Berlin/Heidelberg, Germany, 2011; pp. 113–136. [Google Scholar]
- Flora, G.; Gupta, D.; Tiwari, A. Toxicity of lead: A review with recent updates. Interdiscip. Toxicol. 2012, 5, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.; Xi, S. The effects of heavy metals on human metabolism. Toxicol. Mech. Methods 2020, 30, 167–176. [Google Scholar] [CrossRef]
- Kumar, A.; Kumar, A.; M.M.S., C.-P.; Chaturvedi, A.K.; Shabnam, A.A.; Subrahmanyam, G.; Mondal, R.; Gupta, D.K.; Malyan, S.K.; Kumar, S.S.; et al. Lead toxicity: Health hazards, influence on food chain, and sustainable remediation approaches. Int. J. Environ. Res. Public Health 2020, 17, 2179. [Google Scholar] [CrossRef]
- Zulfiqar, U.; Farooq, M.; Hussain, S.; Maqsood, M.; Hussain, M.; Ishfaq, M.; Ahmad, M.; Anjum, M.Z. Lead toxicity in plants: Impacts and remediation. J. Environ. Manag. 2019, 250, 109557. [Google Scholar] [CrossRef]
- Purushotam, D.; Lone, M.A.; Rashid, M.; Rao, A.N.; Ahmed, S. Deciphering heavy metal contamination zones in soils of a granitic terrain of Southern India using factor analysis and GIS. J. Earth Syst. Sci. 2012, 121, 1059–1070. [Google Scholar] [CrossRef]
- Hadi, F.; Aziz, T. A mini review on lead (Pb) toxicity in plants. J. Biol. Life Sci. 2015, 6, 91. [Google Scholar] [CrossRef]
- Aslam, M.; Aslam, A.; Sheraz, M.; Ali, B.; Ulhassan, Z.; Najeeb, U.; Zhou, W.; Gill, R.A. Lead toxicity in cereals: Mechanistic insight into toxicity, mode of action, and management. Front. Plant Sci. 2021, 11. [Google Scholar] [CrossRef]
- Seneviratne, M.; Rajakaruna, N.; Rizwan, M.; Madawala, H.M.S.P.; Ok, Y.S.; Vithanage, M. Heavy metal-induced oxidative stress on seed germination and seedling development: A critical review. Environ. Geochem. Health 2019, 41, 1813–1831. [Google Scholar] [CrossRef]
- Singh, H.P.; Kaur, G.; Batish, D.R.; Kohli, R.K. Lead (Pb)-inhibited radicle emergence in Brassica campestris involves alterations in starch-metabolizing enzymes. Biol. Trace Elem. Res. 2011, 144, 1295–1301. [Google Scholar] [CrossRef]
- Karalija, E.; Selović, A.; Dahija, S.; Demir, A.; Samardžić, J.; Vrobel, O.; Ćavar Zeljković, S.; Parić, A. Use of seed priming to improve cd accumulation and tolerance in Silene sendtneri, novel cd hyper-accumulator. Ecotoxicol. Environ. Saf. 2021, 210, 111882. [Google Scholar] [CrossRef]
- Khan, A.R.; Azhar, W.; Wu, J.; Ulhassan, Z.; Salam, A.; Zaidi, S.H.R.; Yang, S.; Song, G.; Gan, Y. Ethylene participates in zinc oxide nanoparticles induced biochemical, molecular and ultrastructural changes in rice seedlings. Ecotoxicol. Environ. Saf. 2021, 226, 112844. [Google Scholar] [CrossRef]
- Monreal, C.M.; DeRosa, M.; Mallubhotla, S.C.; Bindraban, P.S.; Dimkpa, C. Nanotechnologies for increasing the crop use efficiency of fertilizer-micronutrients. Biol. Fertil. Soils 2016, 52, 423–437. [Google Scholar] [CrossRef]
- Sharma, V.; Kumar, A.; Dhawan, A. Nanomaterials: Exposure, effects and toxicity assessment. Proc. Natl. Acad. Sci. USA India Sect. B Biol. Sci. 2012, 82, 3–11. [Google Scholar] [CrossRef]
- Khan, A.R.; Wakeel, A.; Muhammad, N.; Liu, B.; Wu, M.; Liu, Y.; Ali, I.; Zaidi, S.H.R.; Azhar, W.; Song, G.; et al. Involvement of ethylene signaling in zinc oxide nanoparticle-mediated biochemical changes in Arabidopsis thaliana leaves. Environ. Sci. Nano 2019, 6, 341–355. [Google Scholar] [CrossRef]
- Salam, A.; Khan, A.R.; Liu, L.; Yang, S.; Azhar, W.; Ulhassan, Z.; Zeeshan, M.; Wu, J.; Fan, X.; Gan, Y. Seed priming with zinc oxide nanoparticles downplayed ultrastructural damage and improved photosynthetic apparatus in maize under cobalt stress. J. Hazard. Mater. 2022, 423, 127021. [Google Scholar] [CrossRef]
- Singh, J.; Lee, B.-K. Influence of Nano-TiO2 particles on the bioaccumulation of cd in soybean plants (Glycine max): A possible mechanism for the removal of Cd from the contaminated soil. J. Environ. Manag. 2016, 170, 88–96. [Google Scholar] [CrossRef]
- Ji, Y.; Zhou, Y.; Ma, C.; Feng, Y.; Hao, Y.; Rui, Y.; Wu, W.; Gui, X.; Le, V.N.; Han, Y.; et al. Jointed toxicity of TiO2 NPs and Cd to rice seedlings: NPs alleviated Cd toxicity and Cd promoted NPs uptake. Plant Physiol. Biochem. 2017, 110, 82–93. [Google Scholar] [CrossRef]
- Andreini, C.; Banci, L.; Bertini, I.; Rosato, A. Zinc through the three domains of life. J. Proteome Res. 2006, 5, 3173–3178. [Google Scholar] [CrossRef]
- Coleman, J.E. Zinc enzymes. Curr. Opin. Chem. Biol. 1998, 2, 222–234. [Google Scholar] [CrossRef]
- Chang, C.Y.; Yu, H.Y.; Chen, J.J.; Li, F.B.; Zhang, H.H.; Liu, C.P. Accumulation of heavy metals in leaf vegetables from agricultural soils and associated potential health risks in the pearl river delta, South China. Environ. Monit. Assess. 2014, 186, 1547–1560. [Google Scholar] [CrossRef]
- Zhou, H.; Yang, W.-T.; Zhou, X.; Liu, L.; Gu, J.-F.; Wang, W.-L.; Zou, J.-L.; Tian, T.; Peng, P.-Q.; Liao, B.-H. Accumulation of heavy metals in vegetable species planted in contaminated soils and the health risk assessment. Int. J. Environ. Res. Public Health 2016, 13, 289. [Google Scholar] [CrossRef]
- Ulhassan, Z.; Gill, R.A.; Huang, H.; Ali, S.; Mwamba, T.M.; Ali, B.; Huang, Q.; Hamid, Y.; Khan, A.R.; Wang, J.; et al. Selenium mitigates the chromium toxicity in Brassicca napus L. by ameliorating nutrients uptake, amino acids metabolism and antioxidant defense system. Plant Physiol. Biochem. 2019, 145, 142–152. [Google Scholar] [CrossRef]
- Ulhassan, Z.; Gill, R.A.; Ali, S.; Mwamba, T.M.; Ali, B.; Wang, J.; Huang, Q.; Aziz, R.; Zhou, W. Dual behavior of Selenium: Insights into physio-biochemical, anatomical and molecular analyses of four Brassica napus cultivars. Chemosphere 2019, 225, 329–341. [Google Scholar] [CrossRef]
- Sagar, V.; Pragya; Bhardwaj, R.; Devi, J.; Singh, S.K.; Singh, P.; Singh, J. The inheritance of betalain pigmentation in Basella alba L. S. Afr. J. Bot. 2022, 145, 360–369. [Google Scholar] [CrossRef]
- Singh, J.; Devi, J.; Sagar, V. Vegetable biofortification: An underexploited silver lining for malnutrition management. In Biofortification of Staple Crops; Springer: Singapore, 2022; pp. 379–416. [Google Scholar]
- Vallabani, N.V.S.; Karakoti, A.S.; Singh, S. ATP-mediated Intrinsic peroxidase-like activity of Fe3O4 -based nanozyme: One step detection of blood glucose at physiological Ph. Colloids Surf. B Biointerfaces 2017, 153, 52–60. [Google Scholar] [CrossRef]
- ISTA. Seed Testing Rules; International Seed Testing Association: Cham, Switzerland, 2015. [Google Scholar]
- Abdul-Baki, A.A.; Anderson, J.D. Vigor determination in soybean seed by multiple criteria. Crop Sci. 1973, 13, 630–633. [Google Scholar] [CrossRef]
- Wilkins, D.A. The measurement of tolerance to edaphic factors by means of root growth. New Phytol. 1978, 80, 623–633. [Google Scholar] [CrossRef]
- Mukherjee, S.P.; Choudhuri, M.A. Implications of water stress-induced changes in the levels of endogenous ascorbic acid and hydrogen peroxide in vigna seedlings. Physiol. Plant. 1983, 58, 166–170. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Bradford, M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Rico, C.M.; Morales, M.I.; McCreary, R.; Castillo-Michel, H.; Barrios, A.C.; Hong, J.; Tafoya, A.; Lee, W.-Y.; Varela-Ramirez, A.; Peralta-Videa, J.R.; et al. Cerium oxide nanoparticles modify the antioxidative stress enzyme activities and macromolecule composition in rice seedlings. Environ. Sci. Technol. 2013, 47, 14110–14118. [Google Scholar] [CrossRef]
- Aebi, H. [13] Catalase in vitro. In Methods in Enzymology; Academic press: Cambridge, MA, USA, 1984; pp. 121–126. [Google Scholar]
- Zhang, R.; Zhang, H.; Tu, C.; Hu, X.; Li, L.; Luo, Y.; Christie, P. Phytotoxicity of ZnO nanoparticles and the released Zn(II) ion to corn (Zea mays L.) and cucumber (Cucumis sativus L.) during germination. Environ. Sci. Pollut. Res. 2015, 22, 11109–11117. [Google Scholar] [CrossRef]
- Zaier, H.; Ghnaya, T.; Lakhdar, A.; Baioui, R.; Ghabriche, R.; Mnasri, M.; Sghair, S.; Lutts, S.; Abdelly, C. Comparative study of pb-phytoextraction potential in Sesuvium portulacastrum and Brassica juncea: Tolerance and accumulation. J. Hazard. Mater. 2010, 183, 609–615. [Google Scholar] [CrossRef]
- Monni, S.; Salemaa, M.; Millar, N. The tolerance of Empetrum nigrum to copper and nickel. Environ. Pollut. 2000, 109, 221–229. [Google Scholar] [CrossRef]
- Bastida, F.; Luis Moreno, J.; Hernández, T.; García, C. Microbiological degradation index of soils in a semiarid climate. Soil Biol. Biochem. 2006, 38, 3463–3473. [Google Scholar] [CrossRef]
- Sagar, V.; Yadav, R.; Gaikwad, K.B.; Gupta, S. Exploring indicator scoring as a selection tool in plant breeding: A study under conservation vs conventional tillage systems. Indian J. Genet. Plant Breed. 2016, 76, 266. [Google Scholar] [CrossRef]
- Kohli, S.K.; Handa, N.; Bali, S.; Khanna, K.; Arora, S.; Sharma, A.; Bhardwaj, R. Current scenario of Pb toxicity in plants: Unraveling plethora of physiological responses. In BT-Reviews of Environmental Contamination and Toxicology; de Voogt, P., Ed.; Springer International Publishing: Cham, Switzerland, 2020; Volume 249, pp. 153–197. ISBN 978-3-030-20194-4. [Google Scholar]
- Romdhane, L.; Panozzo, A.; Radhouane, L.; Dal Cortivo, C.; Barion, G.; Vamerali, T. Root characteristics and metal uptake of maize (Zea mays L.) under extreme soil contamination. Agronomy 2021, 11, 178. [Google Scholar] [CrossRef]
- Ali, S.; Shahbaz, M.; Shahzad, A.N.; Khan, H.A.A.; Anees, M.; Haider, M.S.; Fatima, A. Impact of copper toxicity on stone-head cabbage (Brassica oleracea Var. capitata) in hydroponics. PeerJ 2015, 3, e1119. [Google Scholar] [CrossRef]
- Usman, K.; Abu-Dieyeh, M.H.; Zouari, N.; Al-Ghouti, M.A. Lead (Pb) bioaccumulation and antioxidative responses in Tetraena qataranse. Sci. Rep. 2020, 10, 17070. [Google Scholar] [CrossRef]
- Cokkizgin, A.; Cokkizgin, H. Effects of lead (PbCl2) stress on germination of lentil (Lens culinaris Medic.) lines. African J. Biotechnol. 2010, 9, 8608–8612. [Google Scholar] [CrossRef]
- Huang, T.-L.; Huang, H.-J. ROS and CDPK-like kinase-mediated activation of MAP kinase in rice roots exposed to lead. Chemosphere 2008, 71, 1377–1385. [Google Scholar] [CrossRef]
- Finch-Savage, W.E.; Bassel, G.W. Seed vigour and crop establishment: Extending performance beyond adaptation. J. Exp. Bot. 2016, 67, 567–591. [Google Scholar] [CrossRef]
- Mahakham, W.; Sarmah, A.K.; Maensiri, S.; Theerakulpisut, P. Nanopriming technology for enhancing germination and starch metabolism of aged rice seeds using phytosynthesized silver nanoparticles. Sci. Rep. 2017, 7, 8263. [Google Scholar] [CrossRef]
- Xin, X.; Zhao, F.; Rho, J.Y.; Goodrich, S.L.; Sumerlin, B.S.; He, Z. Use of polymeric nanoparticles to improve seed germination and plant growth under copper stress. Sci. Total Environ. 2020, 745, 141055. [Google Scholar] [CrossRef]
- Tondey, M.; Kalia, A.; Singh, A.; Dheri, G.S.; Taggar, M.S.; Nepovimova, E.; Krejcar, O.; Kuca, K. Seed priming and coating by nano-scale zinc oxide particles improved vegetative growth, yield and quality of fodder maize (Zea mays). Agronomy 2021, 11, 729. [Google Scholar] [CrossRef]
- Sharifan, H.; Moore, J.; Ma, X. Zinc oxide (ZnO) nanoparticles elevated iron and copper contents and mitigated the bioavailability of lead and cadmium in different leafy greens. Ecotoxicol. Environ. Saf. 2020, 191, 110177. [Google Scholar] [CrossRef]
- Dimkpa, C.O.; McLean, J.E.; Latta, D.E.; Manangón, E.; Britt, D.W.; Johnson, W.P.; Boyanov, M.I.; Anderson, A.J. CuO and ZnO nanoparticles: Phytotoxicity, metal speciation, and induction of oxidative stress in sand-grown wheat. J. Nanopartic. Res. 2012, 14, 1125. [Google Scholar] [CrossRef]
- Boonyanitipong, P.; Kositsup, B.; Kumar, P.; Baruah, S.; Dutta, J. Toxicity of ZnO and TiO2 nanoparticles on germinating rice seed Oryza sativa L. Int. J. Biosci. Biochem. Bioinf. 2011, 1, 282–285. [Google Scholar] [CrossRef]
- Kaur, G.; Singh, H.P.; Batish, D.R.; Mahajan, P.; Kohli, R.K.; Rishi, V. Exogenous Nitric oxide (NO) interferes with lead (pb)-induced toxicity by detoxifying reactive oxygen species in hydroponically grown wheat (Triticum aestivum) roots. PLoS ONE 2015, 10, e0138713. [Google Scholar] [CrossRef]
- Zanganeh, R.; Jamei, R.; Rahmani, F. Role of salicylic acid and hydrogen sulfide in promoting lead stress tolerance and regulating free amino acid composition in Zea mays L. Acta Physiol. Plant. 2019, 41, 94. [Google Scholar] [CrossRef]
- Khan, F.; Hussain, S.; Tanveer, M.; Khan, S.; Hussain, H.A.; Iqbal, B.; Geng, M. Coordinated effects of lead toxicity and nutrient deprivation on growth, oxidative status, and elemental composition of primed and non-primed rice seedlings. Environ. Sci. Pollut. Res. 2018, 25, 21185–21194. [Google Scholar] [CrossRef]
- Fahr, M.; Laplaze, L.; Bendaou, N.; Hocher, V.; El Mzibri, M.; Bogusz, D.; Smouni, A. Effect of lead on root growth. Front. Plant Sci. 2013, 4, 175. [Google Scholar] [CrossRef]
- Ueno, D.; Koyama, E.; Yamaji, N.; Ma, J.F. Physiological, genetic, and molecular characterization of a high-Cd-accumulating rice cultivar, Jarjan. J. Exp. Bot. 2011, 62, 2265–2272. [Google Scholar] [CrossRef]
- Wang, M.; Chen, L.; Chen, S.; Ma, Y. Alleviation of cadmium-induced root growth inhibition in crop seedlings by nanoparticles. Ecotoxicol. Environ. Saf. 2012, 79, 48–54. [Google Scholar] [CrossRef]
- Blaylock, M.J.; Salt, D.E.; Dushenkov, S.; Zakharova, O.; Gussman, C.; Kapulnik, Y.; Ensley, B.D.; Raskin, I. Enhanced accumulation of pb in indian mustard by soil-applied chelating agents. Environ. Sci. Technol. 1997, 31, 860–865. [Google Scholar] [CrossRef]
- Siripornadulsil, S.; Traina, S.; Verma, D.P.S.; Sayre, R.T. Molecular mechanisms of proline-mediated tolerance to toxic heavy metals in transgenic microalgae. Plant Cell 2002, 14, 2837–2847. [Google Scholar] [CrossRef]
- Hanikenne, M.; Talke, I.N.; Haydon, M.J.; Lanz, C.; Nolte, A.; Motte, P.; Kroymann, J.; Weigel, D.; Krämer, U. Evolution of metal hyperaccumulation required cis-regulatory changes and triplication of HMA4. Nature 2008, 453, 391–395. [Google Scholar] [CrossRef]
- Yuan, L.; Yang, S.; Liu, B.; Zhang, M.; Wu, K. Molecular characterization of a rice metal tolerance protein, OsMTP1. Plant Cell Rep. 2012, 31, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Derakhshani, B.; Jafary, H.; Maleki Zanjani, B.; Hasanpur, K.; Mishina, K.; Tanaka, T.; Kawahara, Y.; Oono, Y. Combined QTL mapping and RNA-Seq profiling reveals candidate genes associated with cadmium tolerance in barley. PLoS ONE 2020, 15, e0230820. [Google Scholar] [CrossRef] [PubMed]
- Sundaria, N.; Singh, M.; Upreti, P.; Chauhan, R.P.; Jaiswal, J.P.; Kumar, A. Seed priming with iron oxide nanoparticles triggers iron acquisition and biofortification in wheat (Triticum aestivum L.) grains. J. Plant Growth Regul. 2019, 38, 122–131. [Google Scholar] [CrossRef]
- Gokak, I.B.; Taranath, T.C. Morphological and biochemical responses of Abelmoschus esculantus (L.) Moench to zinc nanoparticles. Adv. Nat. Sci. Nanosci. Nanotechnol. 2015, 6, 025017. [Google Scholar] [CrossRef]
- Sharma, S.S.; Dietz, K.-J. The relationship between metal toxicity and cellular redox imbalance. Trends Plant Sci. 2009, 14, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Lin, Z.-F.; Lin, G.-Z.; Song, L.-Y.; Chen, S.-W.; Mo, H.; Peng, C.-L. Lead and cadmium induced alterations of cellular functions in leaves of Alocasia macrorrhiza L. Schott. Ecotoxicol. Environ. Saf. 2010, 73, 1238–1245. [Google Scholar] [CrossRef] [PubMed]
- Malecka, A.; Piechalak, A.; Tomaszewska, B. Reactive oxygen species production and antioxidative defense system in pea root tissues treated with lead ions: The whole roots level. Acta Physiol. Plant. 2009, 31, 1053–1063. [Google Scholar] [CrossRef]
- Kaur, G.; Singh, H.P.; Batish, D.R.; Kohli, R.K. Lead (Pb)-induced biochemical and ultrastructural changes in wheat (Triticum aestivum) roots. Protoplasma 2013, 250, 53–62. [Google Scholar] [CrossRef]
- Holmström, K.M.; Finkel, T. Cellular mechanisms and physiological consequences of redox-dependent signalling. Nat. Rev. Mol. Cell Biol. 2014, 15, 411–421. [Google Scholar] [CrossRef]
- Xia, C.; Hong, L.; Yang, Y.; Yanping, X.; Xing, H.; Gang, D. Protein changes in response to lead stress of lead-tolerant and lead-sensitive industrial hemp using SWATH technology. Genes 2019, 10, 396. [Google Scholar] [CrossRef] [Green Version]
- Dietz, K.J.; Baier, M.; Krämer, U. Free radicals and reactive oxygen species as mediators of heavy metal toxicity in plants. In Heavy Metal Stress in Plants; Springer: Berlin/Heidelberg, Germany, 1999. [Google Scholar]
- Meng, J.; Wang, W.-X.; Li, L.; Zhang, G. Tissue-specific molecular and cellular toxicity of pb in the oyster (Crassostrea gigas): mRNA expression and physiological studies. Aquat. Toxicol. 2018, 198, 257–268. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Table. Cont. | Germination (%) | Root Length (cm) | Shoot Length (cm) | Seedling Biomass (mg Seedling−1) | Vigour Index-I | Vigour Index-II | NPI Score | GTI (%) | ||
|---|---|---|---|---|---|---|---|---|---|---|
| Root Tissue | Shoot Tissue | Seedling Biomass | ||||||||
| 0 mM Pb | 71.0 ± 0.82 b | 15.1 ± 0.24 b | 11.6 ± 0.11 c | 26.86 ± 0.08 b | 1895.0 ± 12.1 c | 1907.1 ± 3.7 b | 0.840 | - | - | - |
| ZnNPs | 82.3 ± 0.62 a | 17.3 ± 0.11 a | 13.9 ± 0.15 b | 30.42 ± 0.19 a | 2566.1 ± 8.5 b | 2503.6 ± 3.3 a | 0.931 | 114.9 | 119.4 | 113.3 |
| FeNPs | 84.3 ± 0.67 a | 18.2 ± 0.10 a | 15.0 ± 0.10 a | 31.24 ± 0.17 a | 2796.2 ± 10.5 a | 2633.5 ± 5.1 a | 0.936 | 120.9 | 128.7 | 116.3 |
| 4 mM Pb | 65.0 ± 0.72 cd | 13.1 ± 0.18 c | 9.4 ± 0.09 e | 18.96 ± 0.11 c | 1461.9 ± 13.5 e | 1232.4 ± 2.2 d | 0.721 | 86.7 | 81.2 | 70.6 |
| 4 mM Pb + ZnNPs | 68.7 ± 0.87 bc | 14.7 ± 0.15 b | 10.0 ± 0.07 d | 19.40 ± 0.16 c | 1696.9 ± 9.7 d | 1332.8 ± 6.1 cd | 0.879 | 97.5 | 86.1 | 72.2 |
| 4 mM Pb + FeNPs | 71.0 ± 0.59 b | 15.7 ± 0.16 b | 11.8 ± 0.10 c | 20.69 ± 0.14 c | 1951.1 ± 11.3 c | 1469.0 ± 2.8 c | 0.900 | 104.2 | 101.4 | 77.0 |
| 8 mM Pb | 49.3 ± 0.61 e | 8.1 ± 0.20 e | 7.9 ± 0.13 fg | 11.65 ± 0.09 e | 786.3 ± 14.5 g | 574.3 ± 1.8 f | 0.192 | 53.7 | 67.6 | 43.4 |
| 8 mM Pb + ZnNPs | 62.7 ± 0.45 d | 8.9 ± 0.17 e | 8.8 ± 0.08 ef | 12.63 ± 0.21 de | 1106.0 ± 9.9 f | 791.9 ± 4.3 e | 0.524 | 59.0 | 75.3 | 47.0 |
| 8 mM Pb + FeNPs | 66.3 ± 0.71 bcd | 11.2 ± 0.32 d | 9.9 ± 0.07 d | 13.64 ± 0.07 d | 1398.3 ± 10.1 e | 904.3 ± 3.8 e | 0.643 | 74.1 | 85.4 | 50.8 |
| 15 mM Pb | 31.3 ± 0.69 g | 3.5 ± 0.28 g | 5.3 ± 0.11 i | 4.53 ± 0.14 g | 276.4 ± 12.5 j | 141.8 ± 2.9 ghi | 0.044 | 23.3 | 45.8 | 16.9 |
| 15 mM Pb + ZnNPs | 38.0 ± 0.55 f | 4.0 ± 0.14 g | 6.5 ± 0.14 h | 5.37 ± 0.15 fg | 399.4 ± 11.9 i | 204.1 ± 5.7 gh | 0.192 | 26.8 | 55.7 | 20.0 |
| 15 mM Pb + FeNPs | 41.0 ± 0.72 f | 6.5 ± 0.18 f | 7.7 ± 0.07 g | 6.51 ± 0.09 f | 581.8 ± 7.8 h | 266.9 ± 4.9 g | 0.235 | 42.9 | 66.4 | 24.2 |
| 20 mM Pb | 19.3 ± 0.19 h | 1.2 ± 0.22 h | 3.6 ± 0.10 j | 1.82 ± 0.11 i | 92.6 ± 5.2 k | 35.1 ± 1.6 i | 0.010 | 8.0 | 30.9 | 6.8 |
| 20 mM Pb + ZnNPs | 26.0 ± 0.23 g | 3.1 ± 0.16 g | 4.7 ± 0.09 i | 2.41 ± 0.17 hi | 202.0 ± 4.4 j | 62.7 ± 4.1 hi | 0.110 | 20.3 | 40.5 | 9.0 |
| 20 mM Pb + FeNPs | 29.9 ± 0.35 g | 4.3 ± 0.23 g | 5.3 ± 0.14 i | 3.81 ± 0.19 gh | 284.6 ± 8.8 j | 113.9 ± 1.9 ghi | 0.116 | 28.3 | 45.2 | 14.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gupta, N.; Singh, P.M.; Sagar, V.; Pandya, A.; Chinnappa, M.; Kumar, R.; Bahadur, A. Seed Priming with ZnO and Fe3O4 Nanoparticles Alleviate the Lead Toxicity in Basella alba L. through Reduced Lead Uptake and Regulation of ROS. Plants 2022, 11, 2227. https://doi.org/10.3390/plants11172227
Gupta N, Singh PM, Sagar V, Pandya A, Chinnappa M, Kumar R, Bahadur A. Seed Priming with ZnO and Fe3O4 Nanoparticles Alleviate the Lead Toxicity in Basella alba L. through Reduced Lead Uptake and Regulation of ROS. Plants. 2022; 11(17):2227. https://doi.org/10.3390/plants11172227
Chicago/Turabian StyleGupta, Nakul, Prabhakar Mohan Singh, Vidya Sagar, Alok Pandya, Manimurugan Chinnappa, Rajesh Kumar, and Anant Bahadur. 2022. "Seed Priming with ZnO and Fe3O4 Nanoparticles Alleviate the Lead Toxicity in Basella alba L. through Reduced Lead Uptake and Regulation of ROS" Plants 11, no. 17: 2227. https://doi.org/10.3390/plants11172227
APA StyleGupta, N., Singh, P. M., Sagar, V., Pandya, A., Chinnappa, M., Kumar, R., & Bahadur, A. (2022). Seed Priming with ZnO and Fe3O4 Nanoparticles Alleviate the Lead Toxicity in Basella alba L. through Reduced Lead Uptake and Regulation of ROS. Plants, 11(17), 2227. https://doi.org/10.3390/plants11172227