
Nitric Oxide Participates in Aluminum-Stress-Induced Pollen Tube Growth Inhibition in Tea (Camelliasinensis) by Regulating CsALMTs

 , ,
, ,
Abstract
:1. Introduction
2. Results
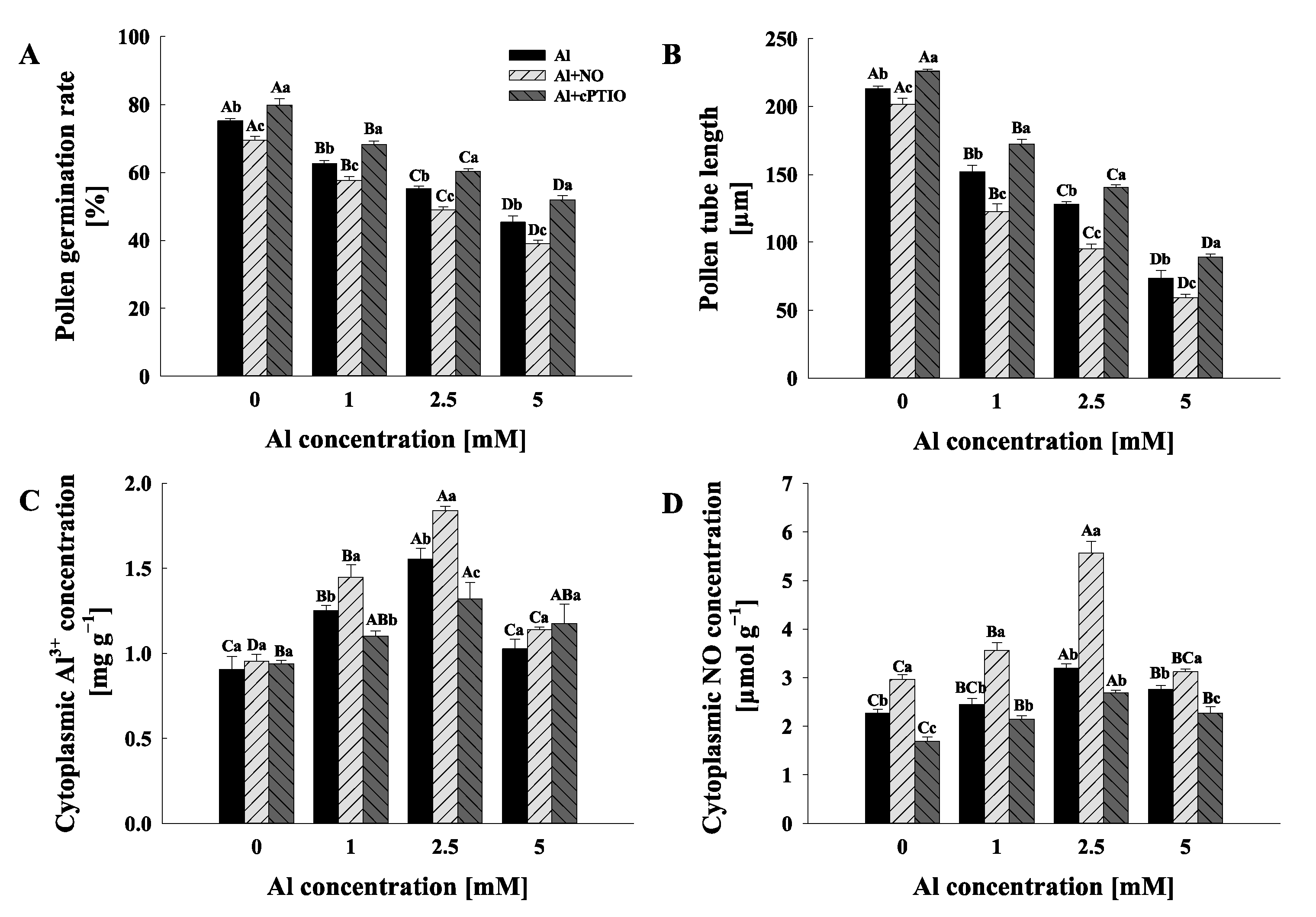
2.1. Pollen Germination and Pollen Tube Elongation
2.2. Cytoplasmic Al3+ and NO
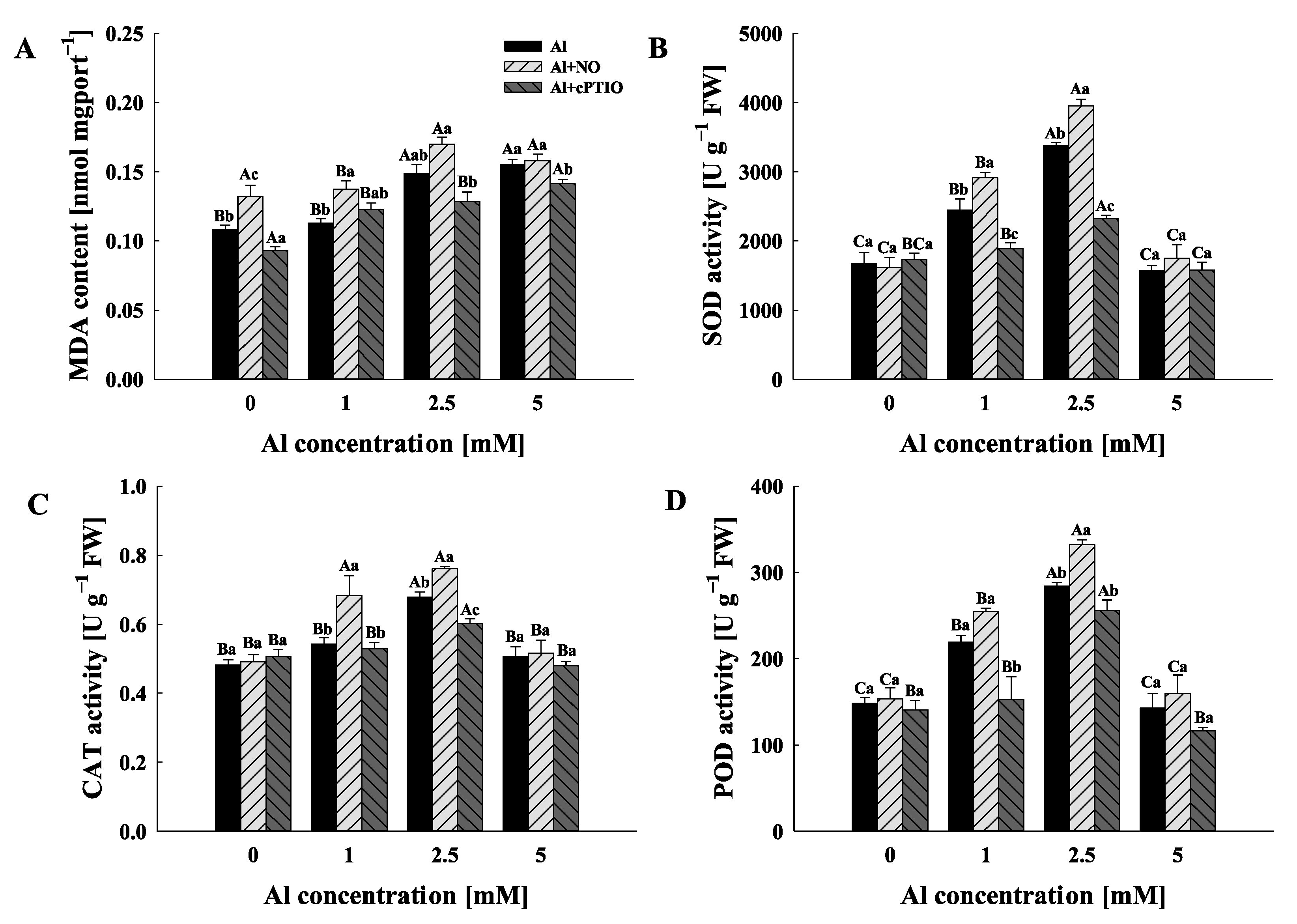
2.3. Physiochemical Responses
2.4. Bioinformatics Analysis for ALMT Gene Family in C. sinensis
2.4.1. Identification, Classification and Naming of ALMT Gene Family in C. sinensis
2.4.2. Basic Physicochemical Properties, Subcellular Localization and Protein Secondary Structure Prediction of CsALMTs
2.4.3. Analysis of Gene Structure and Amino Acid Structure of CsALMTs
2.5. Expression Analysis of CsALMTs in Different C. sinensis Tissues
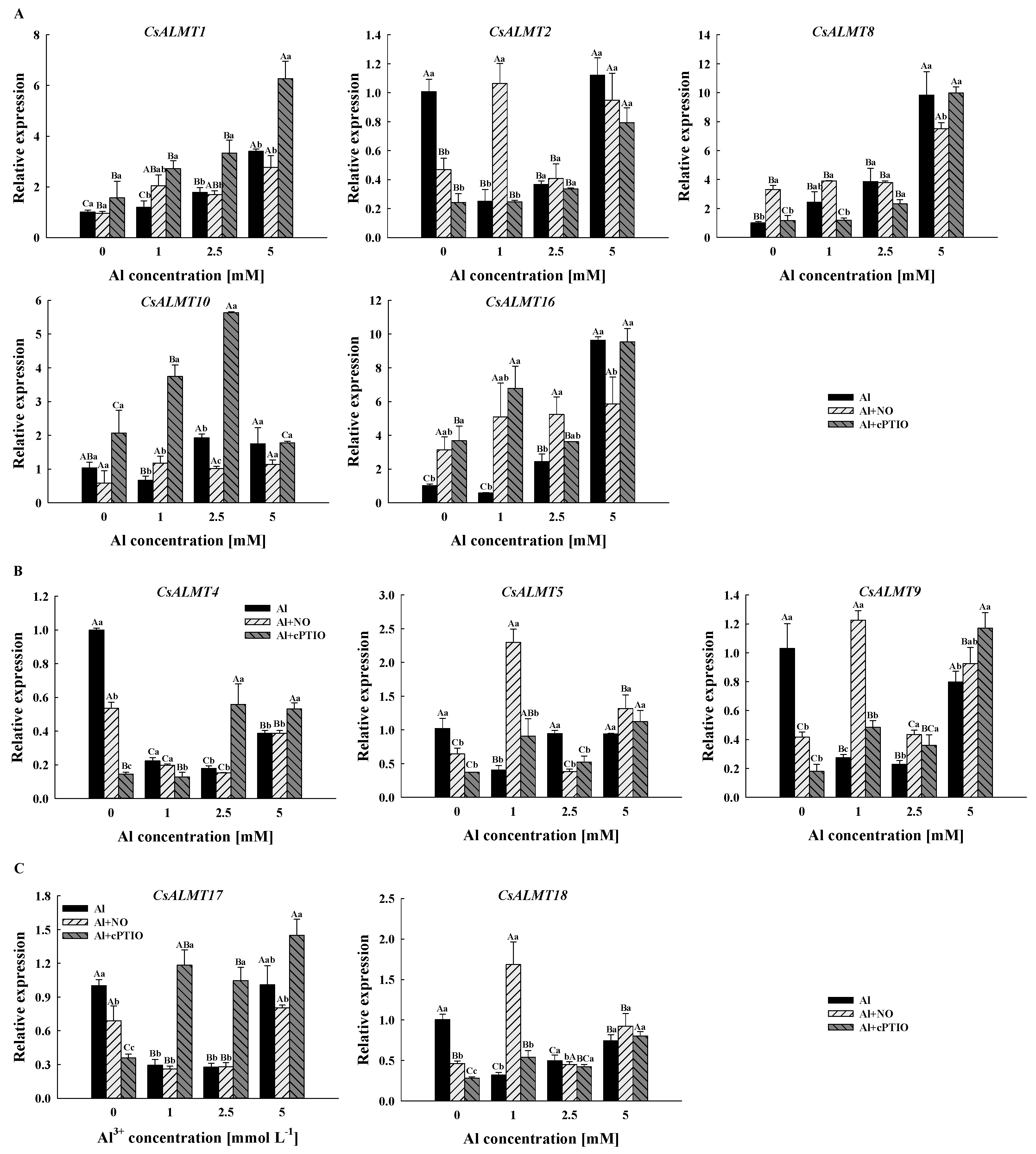
2.6. Effect of Al3+ and NO on CsALMTs’ Expression in Pollen Tube
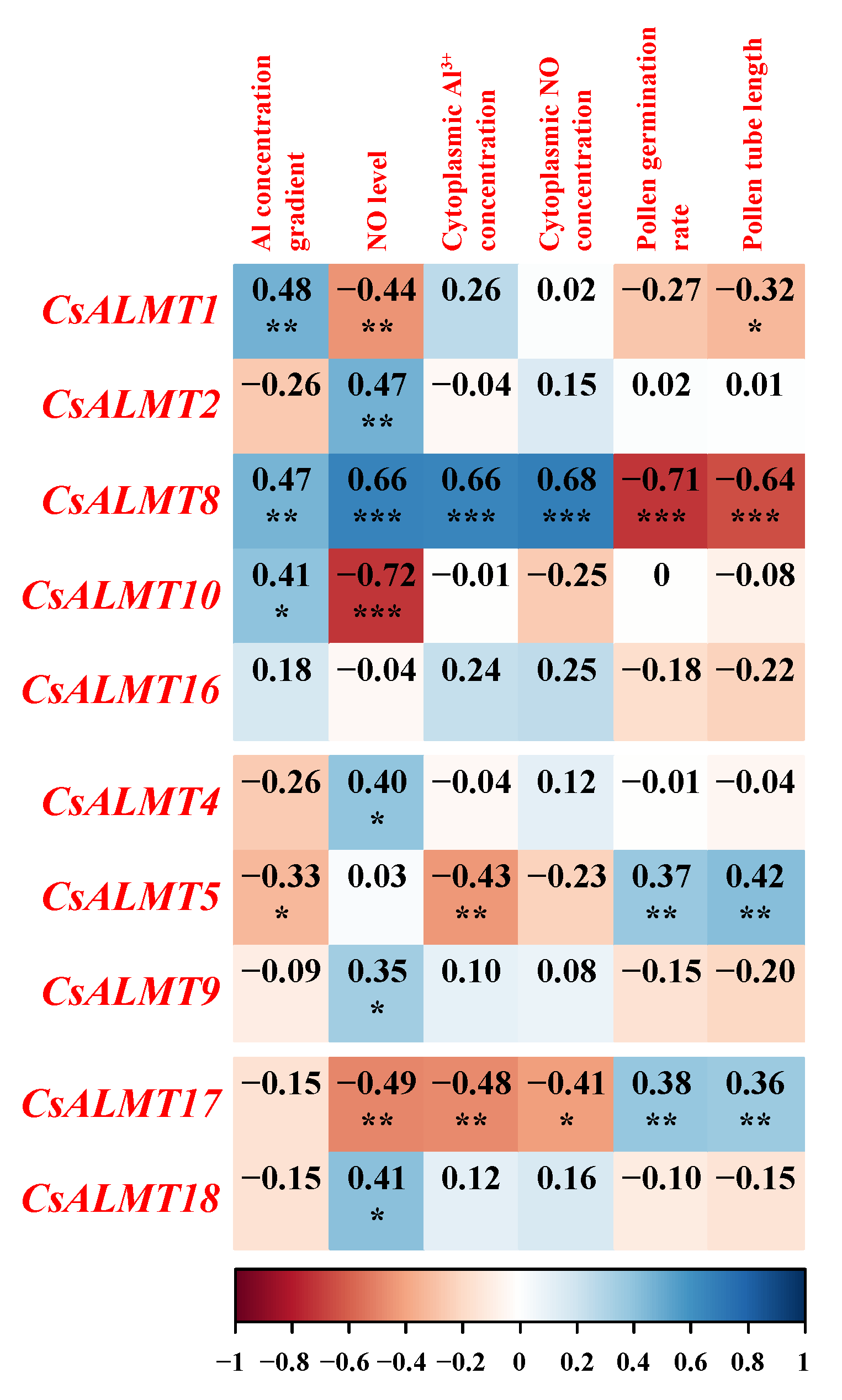
2.7. Two-Way ANOVA Analysis and Pearson Correlation Coefficients
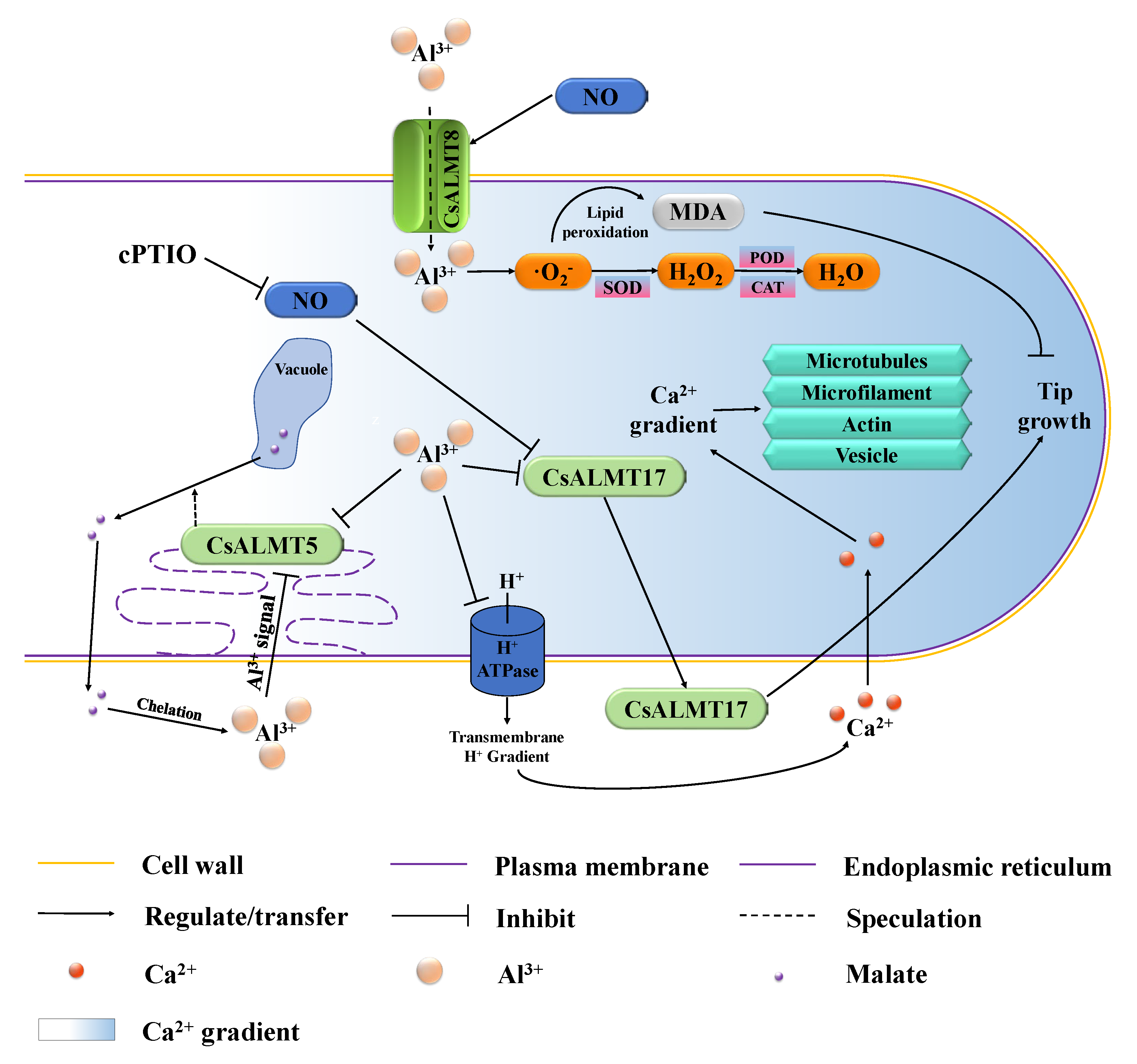
3. Discussion
4. Materials and Methods
4.1. Pollen Source and Culture
4.2. Observation of Pollen Germination Rate and Pollen Tube Elongation
4.3. Measurement of Cytoplasmic Al3+ and NO
4.4. Measurement of Cytoplasmic MDA and Antioxidant Enzyme Activity
4.5. Identification of ALMT Family Genes in C. sinensis
4.6. Bioinformatics Analysis for CsALMTs
4.7. Total RNA Extraction and RT-qPCR Analysis
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dai, B.; Chen, C.; Liu, Y.; Liu, L.; Qaseem, M.F.; Wang, J.; Li, H.; Wu, A.-M. Physiological, biochemical, and transcriptomic responses of neolamarckia cadamba to aluminum stress. Int. J. Mol. Sci. 2020, 21, 9624. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Ai, H.; Wei, M.; Qin, C.; Feng, Y.; Ran, S.; Wei, Z.; Niu, H.; Zhu, Q.; Zhu, H.; et al. Distribution and phytotoxicity of soil labile aluminum fractions and aluminum species in soil water extracts and their effects on tall fescue. Ecotoxicol. Environ. Saf. 2018, 163, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Kochian, L.V. Cellular mechanisms of aluminum toxicity and resistance in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1995, 46, 237–260. [Google Scholar] [CrossRef]
- Barceló, J.; Poschenrieder, C. Fast root growth responses, root exudates, and internal detoxification as clues to the mechanisms of aluminium toxicity and resistance: A review. Environ. Exp. Bot. 2002, 48, 75–92. [Google Scholar] [CrossRef]
- Blamey, F.P.C.; Wheeler, D.M.; Christie, R.A.; Edmeades, D.C. Variation in aluminum tolerance among and within lotus lines. J. Plant Nutr. 1990, 13, 745–755. [Google Scholar] [CrossRef]
- Blancaflor, E.B.; Jones, D.L.; Gilroy, S. Alterations in the cytoskeleton accompany aluminum-induced growth inhibition and morphological changes in primary roots of maize. Plant Physiol. 1998, 118, 159–172. [Google Scholar] [CrossRef]
- Hao, H.; Chen, T.; Fan, L.; Li, R.; Wang, X. 2, 6-dichlorobenzonitrile causes multiple effects on pollen tube growth beyond altering cellulose synthesis in Pinus bungeana Zucc. PLOS ONE 2013, 8, e76660. [Google Scholar] [CrossRef]
- Wang, W.D.; Sheng, X.Y.; Shu, Z.F.; Li, D.Q.; Pan, J.T.; Ye, X.L.; Chang, P.P.; Li, X.H.; Wang, Y.H. Combined cytological and transcriptomic analysis reveals a nitric oxide signaling pathway involved in cold-inhibited Camellia sinensis pollen tube growth. Front. Plant Sci 2016, 7, 456. [Google Scholar] [CrossRef]
- Searcy, K.B.; Mulcahy, D.L. Comparison of the response to aluminum toxicity in gametophyte and sporophyte of four tomato (Lycopersicon esculentum Mill.) cultivars. Theor. Appl. Genet. 1990, 80, 289–295. [Google Scholar] [CrossRef]
- Sawidis, T.; Reiss, H.D. Effects of heavy metals on pollen tube growth and ultrastructure. Protoplasma 1995, 185, 113–122. [Google Scholar] [CrossRef]
- Konishi, S.; Ferguson, I.B.; Putterill, J. Effect of acidic polypeptides on aluminium toxicity in tube growth of pollen from tea (Camellia sinensis L.). Plant Sci. 1988, 56, 55–59. [Google Scholar] [CrossRef]
- Fang, K.; Xie, P.; Zhang, Q.; Xing, Y.; Cao, Q.; Qin, L. Aluminum toxicity-induced pollen tube growth inhibition in apple (Malus domestica) is mediated by interrupting calcium dynamics and modification of cell wall components. Environ. Exp. Bot. 2020, 171, 103928. [Google Scholar] [CrossRef]
- Beligni, M.V.; Lamattina, L. Nitric oxide stimulates seed germination and de-etiolation, and inhibits hypocotyl elongation, three light-inducible responses in plants. Planta 2000, 210, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Shen, W.B.; Xu, L.L. Effects of nitric oxide on the germination of wheat seeds and its reactive oxygen species metabolisms under osmotic stress. Acta Bot. Sinica 2003, 45, 901–905. [Google Scholar]
- Tian, X.; Lei, Y. Nitric oxide treatment alleviates drought stress in wheat seedlings. Biol. Plant. 2006, 50, 775–778. [Google Scholar] [CrossRef]
- Shao, R.; Wang, K.; Shangguan, Z. Cytokinin-induced photosynthetic adaptability of Zea mays L. to drought stress associated with nitric oxide signal: Probed by ESR spectroscopy and fast OJIP fluorescence rise. J. Plant Physiol. 2010, 167, 472–479. [Google Scholar] [CrossRef]
- Wang, H.H.; Huang, J.J.; Bi, Y.R. Nitrate reductase-dependent nitric oxide production is involved in aluminum tolerance in red kidney bean roots. Plant Sci. 2010, 179, 281–288. [Google Scholar] [CrossRef]
- Zhang, Z.Y.; Wang, H.H.; Wang, X.M.; Bi, Y.R. Nitric oxide enhances aluminum tolerance by affecting cell wall polysaccharides in rice roots. Plant Cell Rep. 2011, 30, 1701–1711. [Google Scholar] [CrossRef]
- Bose, J.; Babourina, O.; Rengel, Z. Role of magnesium in alleviation of aluminium toxicity in plants. J. Exp. Bot. 2011, 62, 2251–2264. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, T.; Yamamoto, Y.; Ezaki, B.; Katsuhara, M.; Ahn, S.J.; Ryan, P.R.; Delhaize, E.; Matsumoto, H. A wheat gene encoding an aluminum-activated malate transporter. PlJ 2004, 37, 645–653. [Google Scholar] [CrossRef]
- Liu, J.; Zhou, M.; Delhaize, E.; Ryan, R.P. Altered expression of a Malate-Permeable anion channel, OsALMT4, disrupts mineral nutrition. Plant Physiol. 2017, 175, 1745–1759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoekenga, O.A.; Maron, L.G.; Piñeros, M.A.; Cançado, G.M.A.; Shaff, J.; Kobayashi, Y.; Ryan, P.R.; Dong, B.; Delhaize, E.; Sasaki, T.; et al. AtALMT1, which encodes a malate transporter, is identified as one of several genes critical for aluminum tolerance in Arabidopsis. Proc. Natl. Acad. Sci. USA 2006, 103, 9738–9743. [Google Scholar] [CrossRef] [PubMed]
- Ligaba, A.; Katsuhara, M.; Ryan, P.R.; Shibasaka, M.; Matsumoto, H. The BnALMT1 and BnALMT2 genes from rape encode aluminum-activated malate transporters that enhance the aluminum resistance of plant cells. Plant Physiol. 2006, 142, 1294–1303. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.Y.; Piñeros, M.A.; Tian, J.; Yao, Z.; Sun, L.; Liu, J.; Shaff, J.; Coluccio, A.; Kochian, L.V.; Liao, H. Low pH, aluminum, and phosphorus coordinately regulate malate exudation through GmALMT1 to improve soybean adaptation to acid soils. Plant Physiol. 2013, 161, 1347–1361. [Google Scholar] [CrossRef]
- Chen, Q.; Wu, K.H.; Wang, P.; Yi, J.; Li, K.Z.; Yu, Y.X.; Chen, L.M. Overexpression of MsALMT1, from the aluminum-sensitive medicago sativa, enhances malate exudation and aluminum resistance in Tobacco. Plant Mol. Biol. Report. 2013, 31, 769–774. [Google Scholar] [CrossRef]
- Xu, L.; Qiao, X.; Zhang, M.; Zhang, S. Genome-Wide analysis of aluminum-activated malate transporter family genes in six rosaceae species, and expression analysis and functional characterization on malate accumulation in Chinese white pear. Plant Sci. 2018, 274, 451–465. [Google Scholar] [CrossRef]
- Puyaubert, J.; Baudouin, E. New clues for a cold case: Nitric oxide response to low temperature. Plant Cell Environ. 2014, 37, 2623–2630. [Google Scholar] [CrossRef]
- Sun, C.; Liu, L.; Yu, Y.; Liu, W.; Lu, L.; Jin, C.; Lin, X. Nitric oxide alleviates aluminum-induced oxidative damage through regulating the ascorbate-glutathione cycle in roots of wheat. J. Integr. Plant Biol. 2015, 57, 550–561. [Google Scholar] [CrossRef]
- Besson-Bard, A.; Pugin, A.; Wendehenne, D. New insights into nitric oxide signaling in plants. Annu. Rev. Plant Biol. 2008, 59, 21–39. [Google Scholar] [CrossRef]
- Leterrier, M.; Airaki, M.; Palma, J.M.; Chaki, M.; Barroso, J.B.; Corpas, F.J. Arsenic triggers the nitric oxide (NO) and S-nitrosoglutathione (GSNO) metabolism in Arabidopsis. Environ. Pollut. 2012, 166, 136–143. [Google Scholar] [CrossRef]
- Chen, M.; Cui, W.; Zhu, K.; Xie, Y.; Zhang, C.; Shen, W. Hydrogen-rich water alleviates aluminum-induced inhibition of root elongation in alfalfa via decreasing nitric oxide production. J. Hazard. Mater. 2014, 267, 40–47. [Google Scholar] [CrossRef]
- Sun, C.; Lu, L.; Yu, Y.; Liu, L.; Hu, Y.; Ye, Y.; Jin, C.; Lin, X. Decreasing methylation of pectin caused by nitric oxide leads to higher aluminium binding in cell walls and greater aluminium sensitivity of wheat roots. J. Exp. Bot. 2016, 67, 979–989. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Xu, X.Y.; Chen, L.Q.; Yang, J.L.; Zheng, S.J. Nitric oxide exacerbates Al-induced inhibition of root elongation in rice bean by affecting cell wall and plasma membrane properties. Phytochemistry 2012, 76, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.W.; Zheng, L.P.; Wu, J.Y.; Tan, R.X. Involvement of nitric oxide in oxidative burst, phenylalanine ammonia-lyase activation and Taxol production induced by low-energy ultrasound in Taxus yunnanensis cell suspension cultures. Nitric Oxide 2006, 15, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; An, F.; Wang, L.; Guo, D.; Xie, G.; Liu, Z. Genome-wide identification of Aluminum-Activated Malate Transporter (ALMT) gene family in rubber trees (Hevea brasiliensis) highlights their involvement in aluminum detoxification. Forests 2020, 11, 142. [Google Scholar] [CrossRef]
- Zhang, W.H.; Rengel, Z. Aluminium induces an increase in cytoplasmic calcium in intact wheat root apical cells. Funct. Plant Biol. 1999, 26, 401–409. [Google Scholar] [CrossRef]
- Havlin, J.L.; Soltanpour, P.N. A nitric acid plant tissue digest method for use with inductively coupled plasma spectrometry. Commun. Soil Sci. Plant Anal. 1980, 11, 969–980. [Google Scholar] [CrossRef]
- Lei, Y.; Korpelainen, H.; Li, C. Physiological and biochemical responses to high Mn concentrations in two contrasting Populus cathayana populations. Chemosphere 2007, 68, 686–694. [Google Scholar] [CrossRef]
- Xu, X.; Yang, B.; Qin, G.; Wang, H.; Zhu, Y.; Zhang, K.; Yang, H. Growth, accumulation, and antioxidative responses of two Salix genotypes exposed to cadmium and lead in hydroponic culture. Environ. Sci. Pollu. Res. 2019, 26, 19770–19784. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Song, Y.; Cui, H.; Shi, Y.; Xue, J.; Ji, C.; Zhang, C.; Yuan, L.; Li, R. Genome-wide identification and functional characterization of the Camelina sativa WRKY gene family in response to abiotic stress. BMC Genom. 2020, 21, 786. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Zhu, X.; Shen, J.; Xing, H.; Zou, Z.; Ma, Y.; Wang, Y.; Fang, W. Genome-wide identification, characterization and expression analysis of the amino acid permease gene family in tea plants (Camellia sinensis). Genomics 2020, 112, 2866–2874. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Indicator | Al Concentration Gradient | NO Level | Al Concentration Gradient × NO Level |
|---|---|---|---|
| PGR | 360.73 *** | 98.54 *** | 0.33 ns |
| PTL | 808.36 *** | 110.99 *** | 3.57 * |
| CAl | 59.20 *** | 12.95 *** | 5.01 ** |
| CNO | 93.57 *** | 202.58 *** | 19.51 *** |
| MDA | 45.33 *** | 31.79 *** | 3.25 * |
| SOD | 120.27 *** | 32.78 *** | 11.95 *** |
| CAT | 35.32 *** | 11.38 *** | 3.41 * |
| POD | 85.13 *** | 19.97 *** | 2.43 ns |
| CsALMT1 | 31.707 *** | 23.293 *** | 3.477 * |
| CsALMT2 | 19.830 *** | 13.046 *** | 10.070 *** |
| CsALMT4 | 32.305 *** | 7.324 ** | 23.334 *** |
| CsALMT5 | 50.007 *** | 11.831 *** | 46.449 *** |
| CsALMT8 | 86.568 *** | 2.478 ns | 4.492 ** |
| CsALMT9 | 14.940 *** | 11.086 *** | 18.032 *** |
| CsALMT10 | 16.988 *** | 71.364 *** | 11.985 *** |
| CsALMT16 | 19.636 *** | 6.496 ** | 4.385 ** |
| CsALMT17 | 20.919 *** | 29.545 *** | 15.241 *** |
| CsALMT18 | 9.824 *** | 12.570 *** | 16.588 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, X.; Tian, Z.; Xing, A.; Wu, Z.; Li, X.; Dai, L.; Yang, Y.; Yin, J.; Wang, Y. Nitric Oxide Participates in Aluminum-Stress-Induced Pollen Tube Growth Inhibition in Tea (Camelliasinensis) by Regulating CsALMTs. Plants 2022, 11, 2233. https://doi.org/10.3390/plants11172233
Xu X, Tian Z, Xing A, Wu Z, Li X, Dai L, Yang Y, Yin J, Wang Y. Nitric Oxide Participates in Aluminum-Stress-Induced Pollen Tube Growth Inhibition in Tea (Camelliasinensis) by Regulating CsALMTs. Plants. 2022; 11(17):2233. https://doi.org/10.3390/plants11172233
Chicago/Turabian StyleXu, Xiaohan, Zhiqiang Tian, Anqi Xing, Zichen Wu, Xuyan Li, Lingcong Dai, Yiyang Yang, Juan Yin, and Yuhua Wang. 2022. "Nitric Oxide Participates in Aluminum-Stress-Induced Pollen Tube Growth Inhibition in Tea (Camelliasinensis) by Regulating CsALMTs" Plants 11, no. 17: 2233. https://doi.org/10.3390/plants11172233
APA StyleXu, X., Tian, Z., Xing, A., Wu, Z., Li, X., Dai, L., Yang, Y., Yin, J., & Wang, Y. (2022). Nitric Oxide Participates in Aluminum-Stress-Induced Pollen Tube Growth Inhibition in Tea (Camelliasinensis) by Regulating CsALMTs. Plants, 11(17), 2233. https://doi.org/10.3390/plants11172233