
Vapour Pressure Deficit (VPD) Drives the Balance of Hydraulic-Related Anatomical Traits in Lettuce Leaves





Abstract
:1. Introduction
2. Results
2.1. VPD Influence on Leaf Size and Functional Traits
2.2. VPD Influence on Leaf Stomatal and Vein Traits
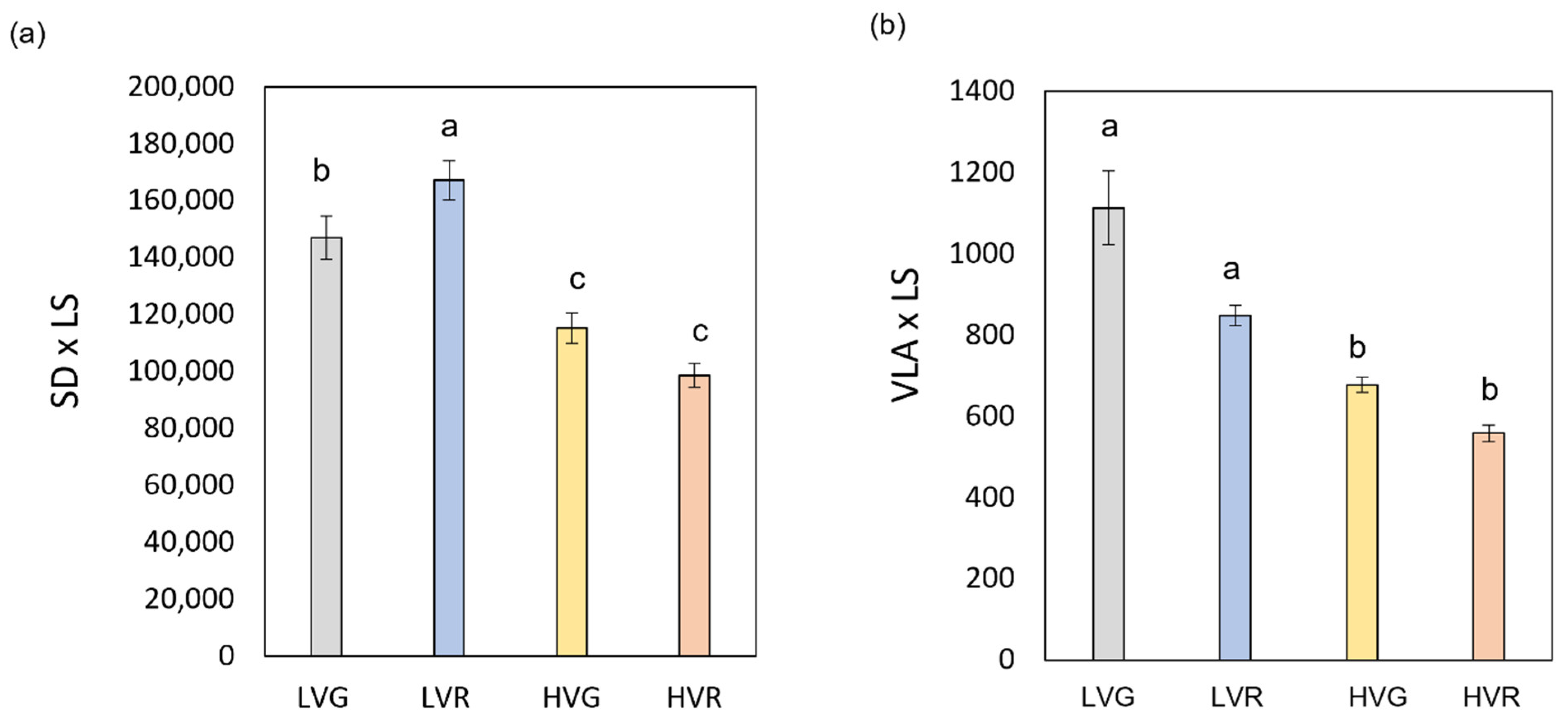
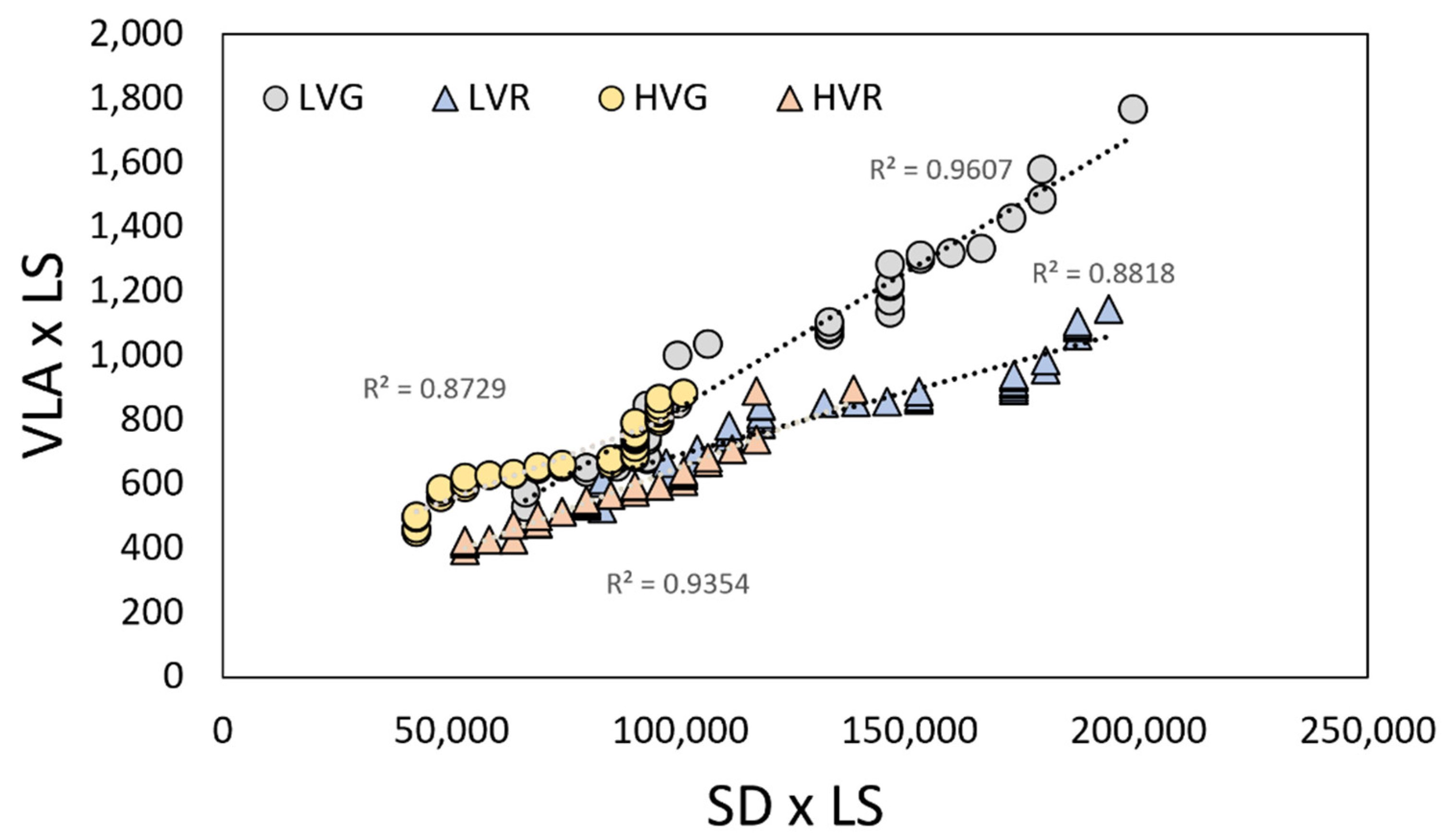
2.3. Relationships between Leaf Morpho-Anatomical Traits
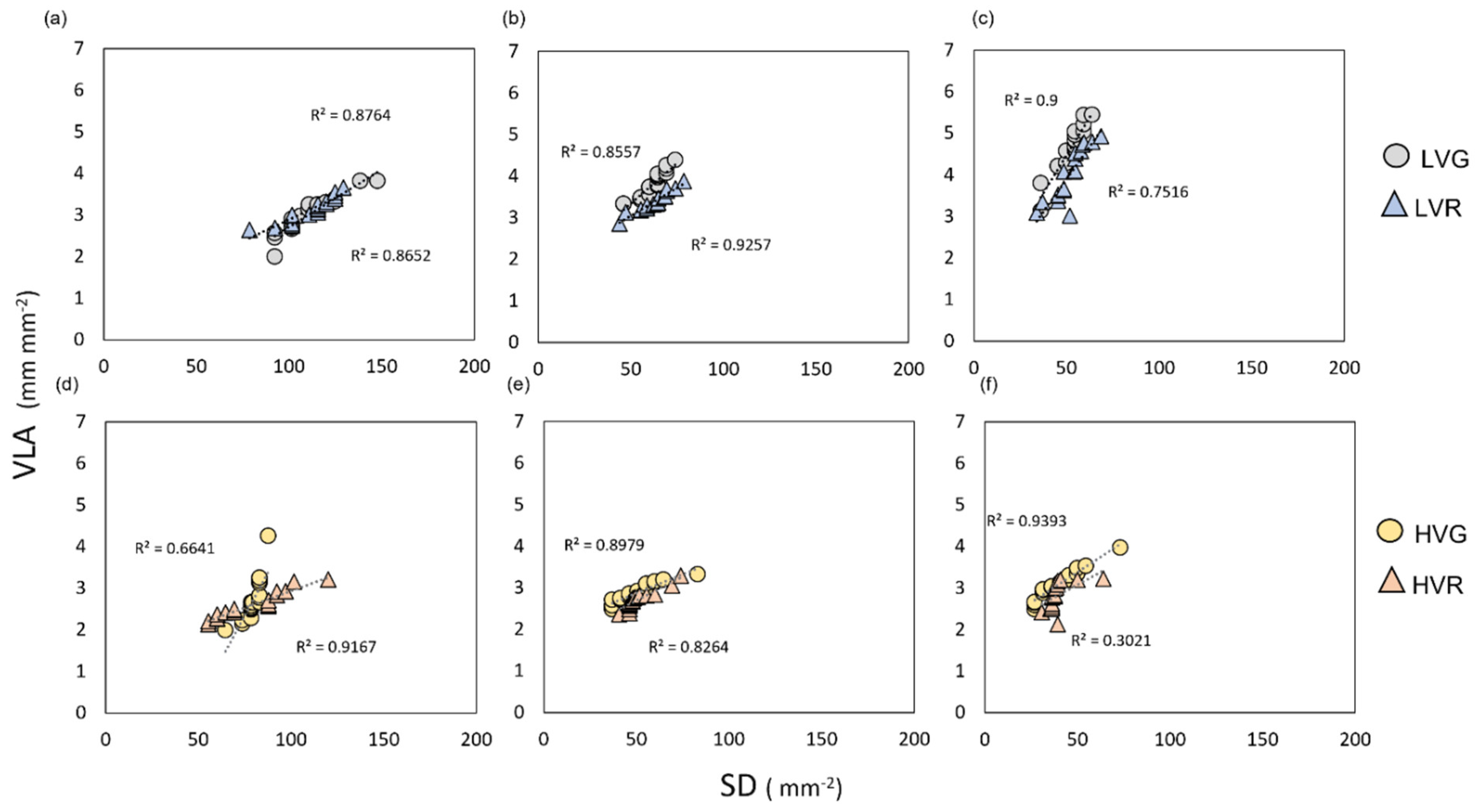
2.4. VPD Influence on Stomatal and Vein Distributions within Leaves
3. Discussion
3.1. Response of Stomatal and Vein Densities and Coordination with Leaf Size under Different VPD
3.2. Acclimation of Anatomy to Sun and Shade within the Same Leaf
4. Materials and Methods
4.1. Plant Material and Growth under Controlled Conditions
4.2. Leaf Size and Other Leaf Functional Traits
4.3. Leaf Vein Traits
4.4. Stomatal Traits
4.5. Relationships between Leaf Size and Leaf Hydraulic-Related Traits
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kattge, J.; Bönisch, G.; Díaz, S.; Lavorel, S.; Prentice, I.C.; Leadley, P.; Tautenhahn, S.; Werner, G.D.; Aakala, T.; Abedi, M. TRY plant trait database–enhanced coverage and open access. Glob. Chang. Biol. 2020, 26, 119–188. [Google Scholar] [CrossRef] [PubMed]
- Valladares, F.; Gianoli, E.; Gómez, J.M. Ecological limits to plant phenotypic plasticity. New Phytol. 2007, 176, 749–763. [Google Scholar] [CrossRef] [PubMed]
- Violle, C.; Navas, M.L.; Vile, D.; Kazakou, E.; Fortunel, C.; Hummel, I.; Garnier, E. Let the concept of trait be functional! Oikos 2007, 116, 882–892. [Google Scholar] [CrossRef]
- Garnier, E.; Navas, M.L. A trait-based approach to comparative functional plant ecology: Concepts, methods and applications for agroecology. A review. Agron. Sustain. Dev. 2012, 32, 365–399. [Google Scholar] [CrossRef]
- Wright, I.J.; Dong, N.; Maire, V.; Prentice, I.C.; Westoby, M.; Díaz, S.; Gallagher, R.V.; Jacobs, B.F.; Kooyman, R.; Law, E.A. Global climatic drivers of leaf size. Science 2017, 357, 917–921. [Google Scholar] [CrossRef]
- Conesa, M.À.; Muir, C.D.; Molins, A.; Galmés, J. Stomatal anatomy coordinates leaf size with Rubisco kinetics in the Balearic Limonium. AoB PLANTS 2020, 12, plz050. [Google Scholar] [CrossRef]
- Ramírez-Valiente, J.A.; Sánchez-Gómez, D.; Aranda, I.; Valladares, F. Phenotypic plasticity and local adaptation in leaf ecophysiological traits of 13 contrasting cork oak populations under different water availabilities. Tree Physiol. 2010, 30, 618–627. [Google Scholar] [CrossRef]
- Yates, M.J.; Verboom, G.A.; Rebelo, A.G.; Cramer, M.D. Ecophysiological significance of leaf size variation in Proteaceae from the Cape Floristic Region. Funct. Ecol. 2010, 24, 485–492. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Portsmuth, A.; Tena, D.; Tobias, M.; Matesanz, S.; Valladares, F. Do we underestimate the importance of leaf size in plant economics? Disproportional scaling of support costs within the spectrum of leaf physiognomy. Ann. Bot. 2007, 100, 283–303. [Google Scholar] [CrossRef]
- Royer, D.L.; Wilf, P.; Janesko, D.A.; Kowalski, E.A.; Dilcher, D.L. Correlations of climate and plant ecology to leaf size and shape: Potential proxies for the fossil record. Am. J. Bot. 2005, 92, 1141–1151. [Google Scholar] [CrossRef] [Green Version]
- Sack, L.; Scoffoni, C. Leaf venation: Structure, function, development, evolution, ecology and applications in the past, present and future. New Phytol. 2013, 198, 983–1000. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-H.; Cai, Y.-F.; Li, S.-F.; Zhang, S.-B. Photosynthetic acclimation of rhododendrons to light intensity in relation to leaf water-related traits. Plant Ecol. 2020, 221, 407–420. [Google Scholar] [CrossRef]
- Scoffoni, C.; Kunkle, J.; Pasquet-Kok, J.; Vuong, C.; Patel, A.J.; Montgomery, R.A.; Givnish, T.J.; Sack, L. Light-induced plasticity in leaf hydraulics, venation, anatomy, and gas exchange in ecologically diverse Hawaiian lobeliads. New Phytol. 2015, 207, 43–58. [Google Scholar] [CrossRef] [PubMed]
- Carins Murphy, M.R.; Jordan, G.J.; Brodribb, T.J. Differential leaf expansion can enable hydraulic acclimation to sun and shade. Plant Cell Environ. 2012, 35, 1407–1418. [Google Scholar] [CrossRef]
- Gupta, B. Correlation of Tissues in Leaves: 1. Absolute Vein-islet Numbers and Absolute Veinlet Termination Numbers. Ann. Bot. 1961, 25, 65–70. [Google Scholar] [CrossRef]
- Gupta, B. Correlation of Tissues in Leaves: 2. Absolute Stomatal Numbers. Ann. Bot. 1961, 25, 71–77. [Google Scholar] [CrossRef]
- Brodribb, T.J.; Feild, T.S.; Jordan, G.J. Leaf maximum photosynthetic rate and venation are linked by hydraulics. Plant Physiol. 2007, 144, 1890–1898. [Google Scholar] [CrossRef]
- Carins Murphy, M.R.; Jordan, G.J.; Brodribb, T.J. Acclimation to humidity modifies the link between leaf size and the density of veins and stomata. Plant Cell Environ. 2014, 37, 124–131. [Google Scholar] [CrossRef]
- Torre, S.; Fjeld, T.; Gislerød, H.R.; Moe, R. Leaf anatomy and stomatal morphology of greenhouse roses grown at moderate or high air humidity. J. Am. Soc. Hortic. Sci. 2003, 128, 598–602. [Google Scholar] [CrossRef]
- Amitrano, C.; Arena, C.; Cirillo, V.; De Pascale, S.; De Micco, V. Leaf morpho-anatomical traits in Vigna radiata L. affect plant photosynthetic acclimation to changing vapor pressure deficit. Environ. Exp. Bot. 2021, 186, 104453. [Google Scholar] [CrossRef]
- Du, Q.; Liu, T.; Jiao, X.; Song, X.; Zhang, J.; Li, J. Leaf anatomical adaptations have central roles in photosynthetic acclimation to humidity. J. Exp. Bot. 2019, 70, 4949–4962. [Google Scholar] [CrossRef] [PubMed]
- Martins, S.C.; Galmes, J.; Cavatte, P.C.; Pereira, L.F.; Ventrella, M.C.; DaMatta, F.M. Understanding the low photosynthetic rates of sun and shade coffee leaves: Bridging the gap on the relative roles of hydraulic, diffusive and biochemical constraints to photosynthesis. PLoS ONE 2014, 9, e95571. [Google Scholar] [CrossRef] [PubMed]
- Mathur, S.; Jain, L.; Jajoo, A. Photosynthetic efficiency in sun and shade plants. Photosynthetica 2018, 56, 354–365. [Google Scholar] [CrossRef]
- Uhl, D.; Mosbrugger, V. Leaf venation density as a climate and environmental proxy: A critical review and new data. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1999, 149, 15–26. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Babani, F. Light adaptation and senescence of the photosynthetic apparatus. Changes in pigment composition, chlorophyll fluorescence parameters and photosynthetic activity. In Chlorophyll a Fluorescence; Springer: Berlin/Heidelberg, Germany, 2004; pp. 713–736. [Google Scholar]
- Lichtenthaler, H.K. Biosynthesis, accumulation and emission of carotenoids, α-tocopherol, plastoquinone, and isoprene in leaves under high photosynthetic irradiance. Photosynth. Res. 2007, 92, 163–179. [Google Scholar] [CrossRef]
- Valladares, F.; Niinemets, Ü. Shade tolerance, a key plant feature of complex nature and consequences. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 237–257. [Google Scholar] [CrossRef]
- Hoshino, R.; Yoshida, Y.; Tsukaya, H. Multiple steps of leaf thickening during sun-leaf formation in Arabidopsis. Plant J. 2019, 100, 738–753. [Google Scholar] [CrossRef]
- Zhang, D.; Jiao, X.; Du, Q.; Song, X.; Li, J. Reducing the excessive evaporative demand improved photosynthesis capacity at low costs of irrigation via regulating water driving force and moderating plant water stress of two tomato cultivars. Agric. Water Manag. 2018, 199, 22–33. [Google Scholar] [CrossRef]
- Knauer, J.; Zaehle, S.; De Kauwe, M.G.; Haverd, V.; Reichstein, M.; Sun, Y. Mesophyll conductance in land surface models: Effects on photosynthesis and transpiration. Plant J. 2020, 101, 858–873. [Google Scholar] [CrossRef]
- Amitrano, C.; Chirico, G.B.; De Pascale, S.; Rouphael, Y.; De Micco, V. Crop Management in Controlled Environment Agriculture (CEA) Systems Using Predictive Mathematical Models. Sensors 2020, 20, 3110. [Google Scholar] [CrossRef]
- Strock, C.F.; Schneider, H.M.; Lynch, J.P. Anatomics: High-throughput phenotyping of plant anatomy. Trends Plant Sci. 2022, 27, 520–523. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Liu, H.; Qiao, Y.; Wang, Y.; Cai, Z.; Dong, B.; Shi, C.; Liu, Y.; Li, X.; Liu, M. Effects of elevated CO2 on the growth, seed yield, and water use efficiency of soybean (Glycine max (L.) Merr.) under drought stress. Agric. Water Manag. 2013, 129, 105–112. [Google Scholar] [CrossRef]
- Amitrano, C.; Rouphael, Y.; De Pascale, S.; De Micco, V. Modulating Vapor Pressure Deficit in the Plant Micro-Environment May Enhance the Bioactive Value of Lettuce. Horticulturae 2021, 7, 32. [Google Scholar] [CrossRef]
- Parkash, V.; Singh, S. A review on potential plant-based water stress indicators for vegetable crops. Sustainability 2020, 12, 3945. [Google Scholar] [CrossRef]
- Jiao, X.C.; Song, X.M.; Zhang, D.L.; Du, Q.J.; Li, J.M. Coordination between vapor pressure deficit and CO2 on the regulation of photosynthesis and productivity in greenhouse tomato production. Sci. Rep. 2019, 9, 8700. [Google Scholar] [CrossRef]
- Lu, N.; Nukaya, T.; Kamimura, T.; Zhang, D.; Kurimoto, I.; Takagaki, M.; Maruo, T.; Kozai, T.; Yamori, W. Control of vapor pressure deficit (VPD) in greenhouse enhanced tomato growth and productivity during the winter season. Sci. Hortic. 2015, 197, 17–23. [Google Scholar] [CrossRef]
- Scoffoni, C.; Rawls, M.; McKown, A.; Cochard, H.; Sack, L. Decline of leaf hydraulic conductance with dehydration: Relationship to leaf size and venation architecture. Plant Physiol. 2011, 156, 832–843. [Google Scholar] [CrossRef]
- Sack, L.; Scoffoni, C. Measurement of leaf hydraulic conductance and stomatal conductance and their responses to irradiance and dehydration using the Evaporative Flux Method (EFM). J. Vis. Exp. 2012, 70, 4179. [Google Scholar] [CrossRef]
- Schneider, J.V.; Habersetzer, J.; Rabenstein, R.; Wesenberg, J.; Wesche, K.; Zizka, G. Water supply and demand remain coordinated during breakdown of the global scaling relationship between leaf size and major vein density. New Phytol. 2017, 214, 473–486. [Google Scholar] [CrossRef]
- Franks, P.J.; Beerling, D.J. Maximum leaf conductance driven by CO2 effects on stomatal size and density over geologic time. Proc. Natl. Acad. Sci. USA 2009, 106, 10343–10347. [Google Scholar] [CrossRef] [Green Version]
- Giday, H.; Kjaer, K.H.; Fanourakis, D.; Ottosen, C.O. Smaller stomata require less severe leaf drying to close: A case study in Rosa hydrida. J. Plant Physiol. 2013, 170, 1309–1316. [Google Scholar] [CrossRef] [PubMed]
- Arve, L.E.; Kruse, O.M.; Tanino, K.K.; Olsen, J.E.; Futsaether, C.; Torre, S. Daily changes in VPD during leaf development in high air humidity increase the stomatal responsiveness to darkness and dry air. J. Plant Physiol. 2017, 211, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Drake, P.L.; Froend, R.H.; Franks, P.J. Smaller, faster stomata: Scaling of stomatal size, rate of response, and stomatal conductance. J. Exp. Bot. 2013, 64, 495–505. [Google Scholar] [CrossRef] [PubMed]
- Poorter, H.; Niinemets, U.; Poorter, L.; Wright, I.J.; Villar, R. Causes and consequences of variation in leaf mass per area (LMA): A meta-analysis. New Phytol. 2009, 182, 565–588. [Google Scholar] [CrossRef]
- Vile, D.; Garnier, E.; Shipley, B.; Laurent, G.; Navas, M.-L.; Roumet, C.; Lavorel, S.; Díaz, S.; Hodgson, J.G.; Lloret, F. Specific leaf area and dry matter content estimate thickness in laminar leaves. Ann. Bot. 2005, 96, 1129–1136. [Google Scholar] [CrossRef]
- Griffith, D.M.; Quigley, K.M.; Anderson, T.M. Leaf thickness controls variation in leaf mass per area (LMA) among grazing-adapted grasses in Serengeti. Oecologia 2016, 181, 1035–1040. [Google Scholar] [CrossRef]
- Brodribb, T.J.; Jordan, G.J. Water supply and demand remain balanced during leaf acclimation of Nothofagus cunninghamii trees. New Phytol. 2011, 192, 437–448. [Google Scholar] [CrossRef]
- Sellin, A.; Taneda, H.; Alber, M. Leaf structural and hydraulic adjustment with respect to air humidity and canopy position in silver birch (Betula pendula). J. Plant Res. 2019, 132, 369–381. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Keenan, T.F.; Hallik, L. A worldwide analysis of within-canopy variations in leaf structural, chemical and physiological traits across plant functional types. New Phytol. 2015, 205, 973–993. [Google Scholar] [CrossRef]
- Gay, A.; Hurd, R. The influence of light on stomatal density in the tomato. New Phytol. 1975, 75, 37–46. [Google Scholar] [CrossRef]
- Xu, Z.; Jiang, Y.; Zhou, G. Response and adaptation of photosynthesis, respiration, and antioxidant systems to elevated CO2 with environmental stress in plants. Front. Plant Sci. 2015, 6, 701. [Google Scholar] [CrossRef] [PubMed]
- Taneda, H.; Tateno, M. Leaf-lamina conductance contributes to an equal distribution of water delivery in current-year shoots of kudzu-vine shoot, Pueraria lobata. Tree Physiol. 2011, 31, 782–794. [Google Scholar] [CrossRef] [PubMed]
- Amitrano, C.; Rouphael, Y.; Pannico, A.; De Pascale, S.; De Micco, V. Reducing the Evaporative Demand Improves Photosynthesis and Water Use Efficiency of Indoor Cultivated Lettuce. Agronomy 2021, 11, 1396. [Google Scholar] [CrossRef]
- Cornelissen, J.; Lavorel, S.; Garnier, E.; Diaz, S.; Buchmann, N.; Gurvich, D.; Reich, P.B.; Ter Steege, H.; Morgan, H.; Van Der Heijden, M. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust. J. Bot. 2003, 51, 335–380. [Google Scholar] [CrossRef]
- Ryser, P.; Urbas, P. Ecological significance of leaf life span among Central European grass species. Oikos 2000, 91, 41–50. [Google Scholar] [CrossRef]
- Witkowski, E.; Lamont, B.B. Leaf specific mass confounds leaf density and thickness. Oecologia 1991, 88, 486–493. [Google Scholar] [CrossRef]
- Elsherif, A.; Gaulton, R.; Shenkin, A.; Malhi, Y.; Mills, J. Three dimensional mapping of forest canopy equivalent water thickness using dual-wavelength terrestrial laser scanning. Agric. For. Meteorol. 2019, 276, 107627. [Google Scholar] [CrossRef]
- Miksche, J.P.; Berlyn, G.P. Botanical Microtechnique and Cytochemistry; Iowa State University Press: Iowa City, IA, USA, 1976. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LS (mm2) | LMA (mg mm−2) | LDMC (g g−1) | RWC (%) | EWT (mg mm−2) | |
|---|---|---|---|---|---|
| VPD (V) | |||||
| LV | 316.8 ± 7.7 a | 0.51 ± 0.03 a | 0.13 ± 0.008 a | 95.1 ± 1.4 a | 3.06 ± 0.13 a |
| HV | 247.5 ± 2.5 b | 0.25 ± 0.01 b | 0.06 ± 0.002 b | 87.7 ± 3.4 b | 3.55 ± 0.09 a |
| Cultivar (C) | |||||
| G | 279.0 ± 6.1 a | 0.38 ± 0.02 a | 0.09 ± 0.006 a | 90.1 ± 2.6 b | 3.26 ± 0.10 a |
| R | 285.2 ± 4.1 a | 0.37 ± 0.02 a | 0.09 ± 0.006 a | 92.9 ± 2.7 a | 3. 35 ± 0.12 a |
| Interaction | |||||
| LVG | 310.1 ± 8.9 a | 0.52 ± 0.03 a | 0.13 ± 0.007 a | 94.3 ± 1.6 b | 3.15 ± 0.13 a |
| LVR | 323.5 ± 6.6 a | 0.51 ± 0.03 a | 0.13 ± 0.009 a | 95.1 ± 1.1 a | 2.98 ± 0.12 a |
| HVG | 247.9 ± 3.3 b | 0.25 ± 0.01 b | 0.06 ± 0.002 b | 89.2 ± 5.1 c | 3.38 ± 0.07 a |
| HVR | 247.0 ± 1.7 b | 0.25 ± 0.01 b | 0.06 ± 0.002 b | 86.2 ± 4.8 c | 3.72 ± 0.11 a |
| Significance | |||||
| VPD | *** | *** | *** | *** | NS |
| C | NS | NS | NS | * | NS |
| VPD × C | NS | NS | NS | ** | NS |
| SL (µm) | SW (µm) | ED (n mm−2) | FEV (n mm−2) | |
|---|---|---|---|---|
| VPD (V) | ||||
| LV | 20.28 ± 0.63 b | 15.91 ± 0.40 a | 1.93 × 108 ± 9.84 × 106 a | 5933 ± 46 a |
| HV | 22.55 ± 0.59 a | 15.93 ± 0.17 a | 1.71 × 108 ± 5.15 × 106 b | 2913 ± 16 b |
| Cultivar (C) | ||||
| G | 22.11 ± 0.42 a | 16.03 ± 0.37 a | 1.71 × 108 ± 5.87 × 106 a | 5048 ± 35 a |
| R | 20.72 ± 0.38 b | 15.92 ± 0.21 a | 1.82 × 108 ± 5.77 × 106 a | 3799 ± 27 b |
| Interaction | ||||
| LVG | 20.31 ± 0.43 b | 16.15 ± 0.56 a | 1.94 × 108 ± 6.69 × 106 a | 6753 ± 3 a |
| LVR | 19.76 ± 0.40 c | 15.73 ± 0.24 a | 1.92 × 108 ± 6.30 × 106 a | 5113 ± 40 b |
| HVG | 22.06 ± 0.41 a | 15.91 ± 0.18 a | 1.71 × 108 ± 5.06 × 106 b | 3343 ± 17 c |
| HVR | 21.27 ± 0.36 a | 15.91 ± 0.17a | 1.71 × 108± 5.25 × 106 b | 2484 ± 15 c |
| Significance | ||||
| VPD | *** | NS | *** | *** |
| C | * | NS | NS | * |
| VPD × C | * | NS | NS | * |
| SD | SL | |||||
|---|---|---|---|---|---|---|
| (n mm2) | (µm) | |||||
| VPD (V) | ||||||
| LV | 76.1 ± 2.43 a | 20.7 ± 0.35 b | ||||
| HV | 56.1 ± 2.71 b | 22.1 ± 0.33 a | ||||
| Cultivar (C) | ||||||
| G | 64.9 ± 2.38 a | 21.4 ± 0.30 a | ||||
| R | 66.4 ± 2.75 b | 21.4 ± 0.37 a | ||||
| Position (P) | ||||||
| apex | 95.0 ± 3.10 a | 22.0 ± 0.32 a | ||||
| bottom | 56.1 ± 2.48 b | 21.7 ± 0.34 b | ||||
| medium | 45.6 ± 2.12 c | 20.5 ± 0.35 c | ||||
| Interaction (V × C × P) | b | m | a | b | m | a |
| LVG | 50.8 ± 1.75 e | 62.1 ± 1.76 c | 110.6 ± 3.78 a | 20.2 ± 0.30 c | 20.5 ± 0.40 c | 18.8 ± 0.12 d |
| LVR | 49.0 ± 2.11 e | 62.4 ± 2.09 c | 111.9 ± 3.09 a | 20.8 ± 0.45 c | 20.0 ± 0.28 c | 17.4 ± 0.52 d |
| HVG | 39.0 ± 2.90 f | 51.7 ± 2.89 d | 78.1 ± 1.23 b | 28.4 ± 0.32 a | 23.5 ± 0.33 b | 24.1 ± 0.32 b |
| HVR | 39.4 ± 1.73 f | 52.4 ± 3.19 de | 79.4 ± 4.32 b | 27.4 ± 0.32 a | 23.5 ± 0.35 b | 23.9 ± 0.32 b |
| Significance | ||||||
| VPD | *** | *** | ||||
| C | NS | NS | ||||
| P | *** | *** | ||||
| V × C × P | NS | * | ||||
| FEV | VLA | |||||
|---|---|---|---|---|---|---|
| (n mm−2) | (mm mm−2) | |||||
| VPD (V) | ||||||
| LV | 18.8 ± 1.07 a | 3.64 ± 0.11 a | ||||
| HV | 10.2 ± 0.58 b | 2.67 ± 0.21 b | ||||
| Cultivar (C) | ||||||
| G | 16.0 ± 0.84 a | 3.31 ± 0.24 a | ||||
| R | 16.8 ± 0.81 a | 3.13 ± 0.09 b | ||||
| Position (P) | ||||||
| apex | 9.77 ± 0.95 c | 2.84 ± 0.11 c | ||||
| bottom | 12.5 ± 0.73 b | 3.19 ± 0.07 b | ||||
| medium | 19.2 ± 0.79 a | 3.63 ± 0.30 a | ||||
| Interaction (V × C × P) | b | m | a | b | m | a |
| LVG | 12.9 ± 0.71 c | 21.8 ± 0.98 b | 30.6 ± 0.54 a | 4.64 ± 0.13 a | 3.77 ± 0.11 b | 2.91 ± 0.15 c |
| LVR | 13.0 ± 0.93 c | 21.8 ± 0.92 b | 31.5 ± 0.32 a | 4.01± 0.14 a | 3.38 ± 0.06 b | 3.13 ± 0.06 c |
| HVG | 8.06 ± 0.87 d | 10.3 ± 0.55 cd | 12.6 ± 0.36 c | 3.06 ± 0.08 c | 2.85 ± 0.06 cd | 2.65 ± 0.88 d |
| HVR | 8.10 ± 0.66 d | 10.0 ± 0.46 cd | 12.0 ± 0.56 c | 2.92 ± 0.07 c | 2.76 ± 0.06 cd | 2.71 ± 0.12 d |
| Significance | ||||||
| VPD | *** | *** | ||||
| C | ** | ** | ||||
| P | *** | *** | ||||
| V × C × P | * | ** | ||||
| Trait | Acronym | Measurement Unit |
|---|---|---|
| Leaf size | LS | mm2 |
| Leaf mass per area | LMA | g mm−2 |
| Leaf dry matter content | LDMC | g g−1 |
| Relative water content | RWC | % |
| Equivalent water thickness | EWT | g mm−2 |
| Stomatal density | SD | n mm−2 |
| Stomatal density per leaf size | SD × LS | n mm−2 LS |
| Stomatal guard cells length | SL | µm |
| Stomatal guard cells width | SW | µm |
| Epidermal cell density | ED | n mm−2 |
| Epidermal cell density per leaf size | ED × LS | n mm−2 LS |
| Vein density | VLA | mm mm−2 |
| Vein density per leaf size | VLA × LS | mm mm−2 LS |
| Free Vein Endings | FEV | n mm−2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amitrano, C.; Rouphael, Y.; De Pascale, S.; De Micco, V. Vapour Pressure Deficit (VPD) Drives the Balance of Hydraulic-Related Anatomical Traits in Lettuce Leaves. Plants 2022, 11, 2369. https://doi.org/10.3390/plants11182369
Amitrano C, Rouphael Y, De Pascale S, De Micco V. Vapour Pressure Deficit (VPD) Drives the Balance of Hydraulic-Related Anatomical Traits in Lettuce Leaves. Plants. 2022; 11(18):2369. https://doi.org/10.3390/plants11182369
Chicago/Turabian StyleAmitrano, Chiara, Youssef Rouphael, Stefania De Pascale, and Veronica De Micco. 2022. "Vapour Pressure Deficit (VPD) Drives the Balance of Hydraulic-Related Anatomical Traits in Lettuce Leaves" Plants 11, no. 18: 2369. https://doi.org/10.3390/plants11182369
APA StyleAmitrano, C., Rouphael, Y., De Pascale, S., & De Micco, V. (2022). Vapour Pressure Deficit (VPD) Drives the Balance of Hydraulic-Related Anatomical Traits in Lettuce Leaves. Plants, 11(18), 2369. https://doi.org/10.3390/plants11182369