

Domestication Syndrome in Dacryodes edulis (Burseraceae): Comparison of Morphological and Biochemical Traits between Wild and Cultivated Populations

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Sex Distribution in D. edulis
2.2. Morphometric Characteristics of Flowers according to the Sexual Type, Status of the Individuals and Sites
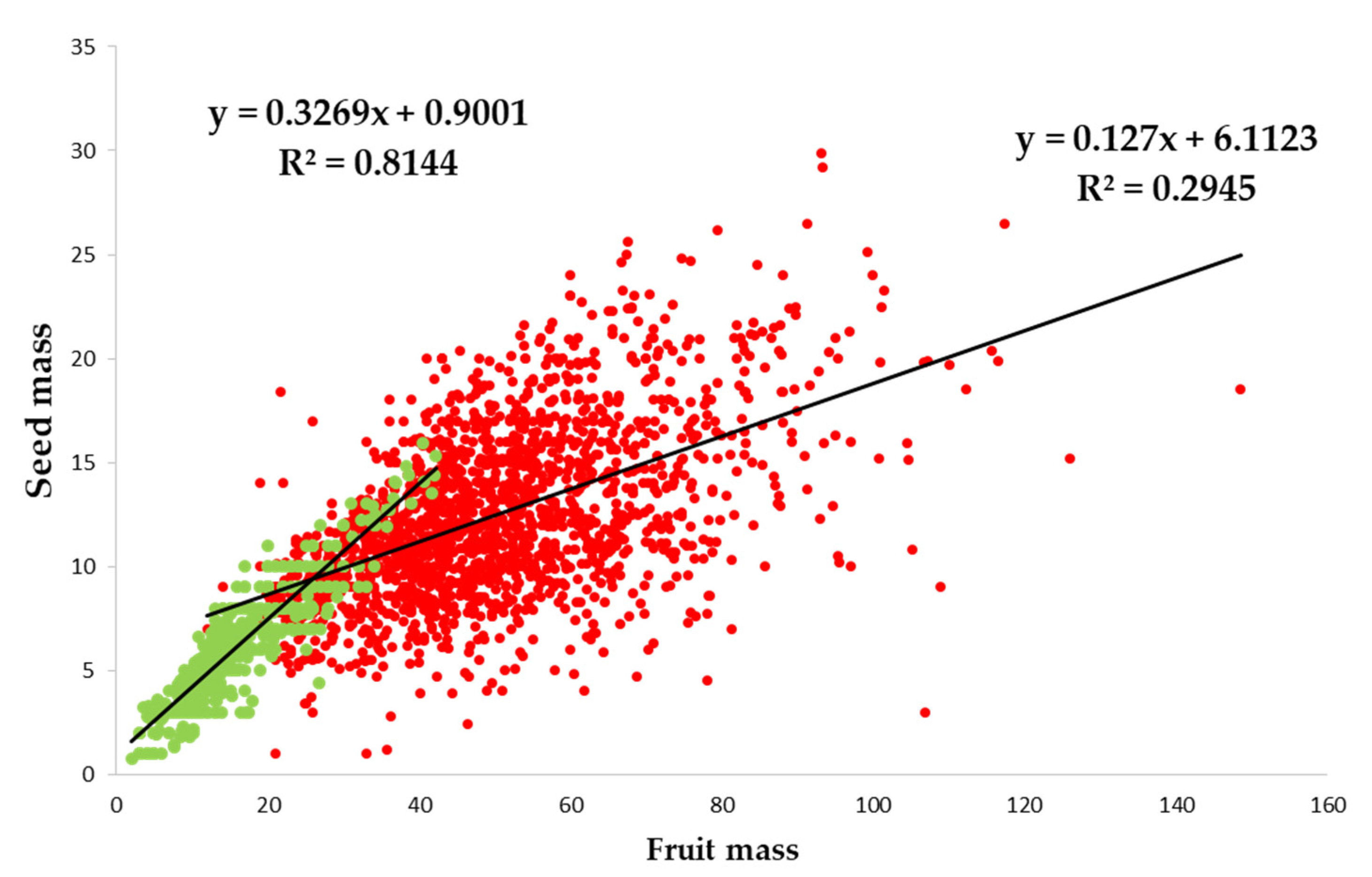
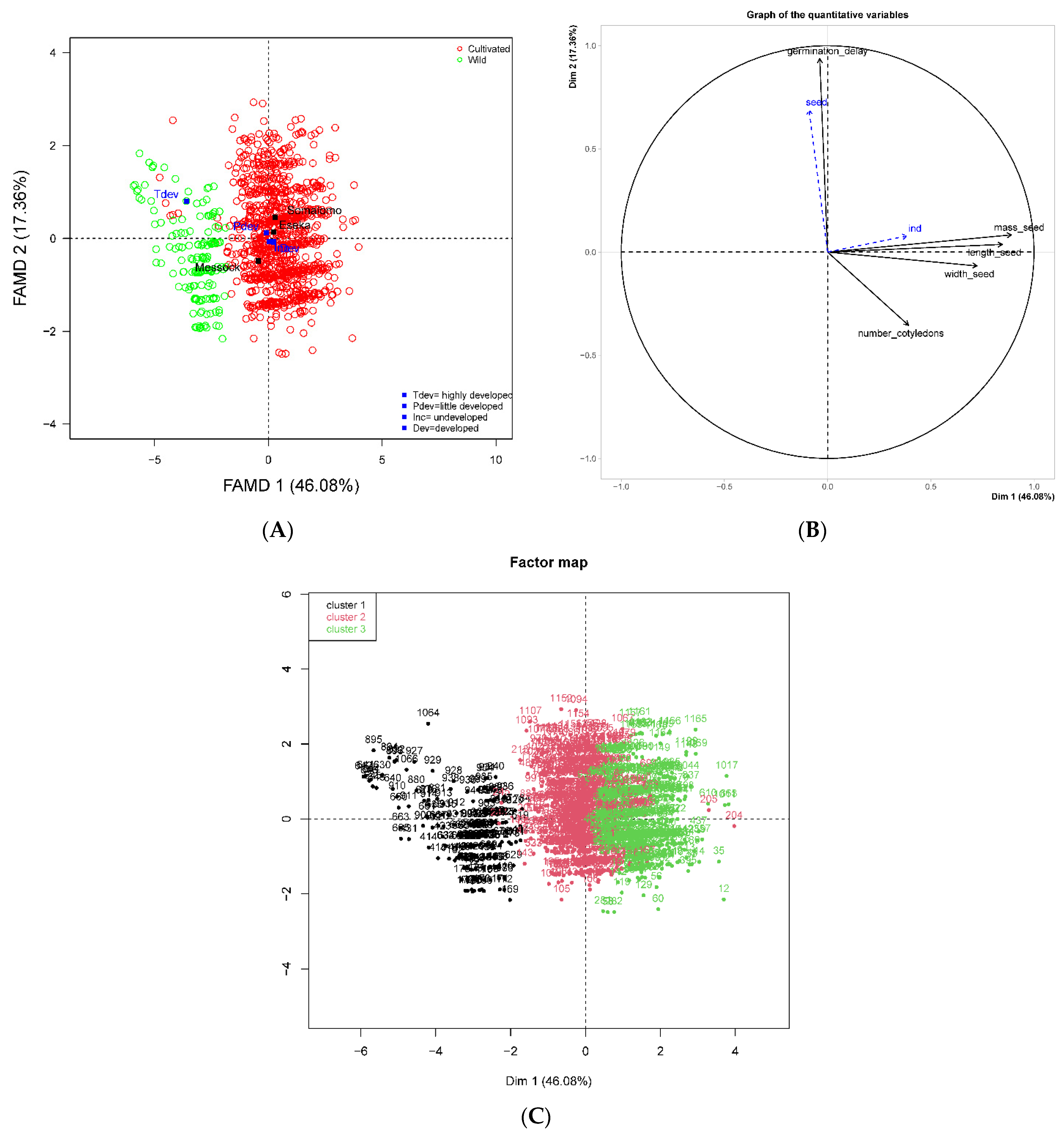
2.3. Morphometric Characteristics of Fruits and Seeds
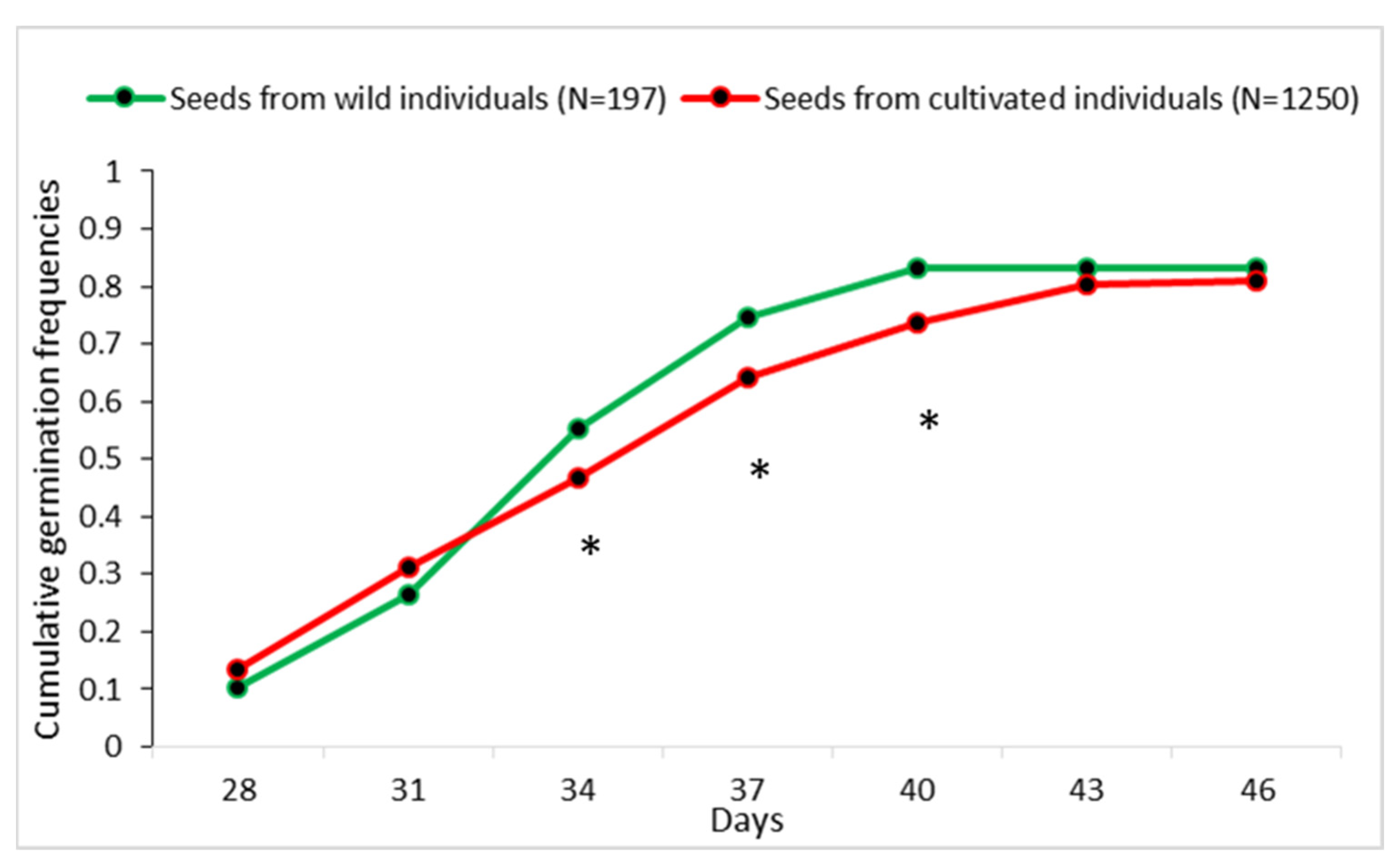
2.4. Seed Germination
2.5. Mesocarp Lipid Content and Fatty Acid Composition
3. Discussion
3.1. Effect of Selection on Sex Distribution and Flower Morphometric Characteristics
3.2. Effect of Selection on the Morphometric Parameters of Fruits and Seeds and on the Germination Dynamic
3.3. Effect of Selection on the Lipid Composition of D. edulis Fruits
4. Material and Methods
4.1. Study Sites
4.2. Sex Distribution and Flower Morphometric Characteristics
4.3. Fruit and Seed’s Morphometric Characteristics
4.4. Seed Germination Tests
4.5. Oil Content and Fatty Acid Composition of D. edulis Fruits
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Begna, T. Importance and Impact of Ecological Approaches to Crop Domestication. J. Biol. Agric. Healthc. 2020, 10, 32–37. [Google Scholar]
- Smith, B.D. The Cultural Context of Plant Domestication in Eastern North America. Curr. Anthropol. 2011, 52 (Suppl. S4), S471–S484. [Google Scholar] [CrossRef]
- Gepts, P. Crop Domestication as a Long-Term Selection Experiment. Plant Breed. 2004, 24, 1–44. [Google Scholar]
- Clement, C.R. 1492 and the Loss of Amazonian Crop Genetic Resources. I. The Relation between Domestication and Human Population Decline. Econ. Bot. 1999, 53, 188–202. [Google Scholar] [CrossRef]
- Lavachery, P. The Holocene Archaeological Sequence of Shum Laka Rock Shelter (Grassfields, Western Cameroon). Afr. Archaeol. Rev. 2001, 18, 213–247. [Google Scholar] [CrossRef]
- Oslisly, R.; White, L. Human impact and environmental exploitation in Gabon and Cameroon during the Holocene. In Rethinking Agriculture; Archaeological and Ethnoarchaeological Perspectives; Denham, T.P., Iriarte, J., Vrydaghs, L., Eds.; Left Coast Press Inc.: Walnut Creek, CA, USA, 2007; pp. 345–358. [Google Scholar]
- Morin-Rivat, J.; Fayolle, A.; Gillet, J.-F.; Bourland, N.; Gourlet-Fleury, S.; Oslisly, R.; Bremond, L.; Bentaleb, I.; Beeckman, H.; Doucet, J.-L. New Evidence of Human Activities during the Holocene in the Lowland Forests of the Northern Congo Basin. Radiocarbon 2014, 56, 209–220. [Google Scholar] [CrossRef]
- Vleminckx, J.; Morin-Rivat, J.; Biwolé, A.B.; Daïnou, K.; Gillet, J.F.; Doucet, J.L.; Drouet, T.; Hardy, O.J. Soil Charcoal to Assess the Impacts of Past Human Disturbances on Tropical Forests. PLoS ONE 2014, 9, e108121. [Google Scholar] [CrossRef]
- Miller, A.J.; Gross, B.L. From Forest to Field: Perennial Fruit Crop Domestication. Am. J. Bot. 2011, 98, 1389–1414. [Google Scholar] [CrossRef]
- Meyer, R.S.; Duval, A.E.; Jensen, H.R. Tansley Review Patterns and Processes in Crop Domestication: An Historical Review and Quantitative Analysis of 203 Global Food Crops. New Phyt. 2012, 196, 29–48. [Google Scholar] [CrossRef]
- Pedrosa, H.C.; Clement, C.R.; Schietti, J. The Domestication of the Amazon Tree Grape (Pourouma Cecropiifolia) under an Ecological Lens. Front. Plant Sci. 2018, 9, 203. [Google Scholar] [CrossRef]
- Meyer, R.S.; Purugganan, M.D. Evolution of Crop Species: Genetics of Domestication and Diversification. Nat. Rev. Genet. 2013, 14, 840–852. [Google Scholar] [CrossRef] [PubMed]
- Parker, I.M.; López, I.; Petersen, J.J.; Anaya, N.; Cubilla-Rios, L.; Potter, D. Domestication Syndrome in Caimito (Chrysophyllum Cainito L.): Fruit and Seed Characteristics. Econ. Bot. 2010, 64, 161–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iñiguez, J.C.; Hernández, M.S.; Galarza, M.D.L.A.; Arrazate, C.H.A.; Medina, J.F.A.; Posadas, L.D.M.R. Caracterización Bioquímica de Variedades Domesticadas de Chayote Sechium edule (Jacq.) Sw. Comparadas Con Parientes Silvestres. Rev. Chapingo. Ser. Hortic. 2011, 17, 45–55. [Google Scholar]
- Dolley, N.; Lyngdoh, N.; Singh, S.; Singh, M.C.; Devi, M.B.; Hazarika, B.N. Domestication of Phoebe cooperiana in the Eastern Himalayas: Population Variation in Morphological and Biochemical Fruit Parameters. Plant Genet. Resour. 2020, 18, 259–269. [Google Scholar] [CrossRef]
- Awono, A.; Ndoye, O.; Schreckenberg, K.; Tabuna, H.; Isseri, F.; Temple, L. Production and Marketing of Safou (Dacryodes Edulis) in Cameroon and Internationally: Market Development Issues. For. Trees Livelihoods 2002, 12, 125–147. [Google Scholar] [CrossRef]
- Biloso, A.; Tsobeng, A.; Akalakou, C.; Mwengi, I.; Kapalay, O. Revue Africaine d ’ Environnement et d ’ Agriculture Variation Phénotypique Des Fruits Des Écotypes de Dacryodes Edulis (G. Dom) H. J. Lam Récoltés Dans Trois Zones Agro-Écologiques de La République Démocratique Du Congo. Rev. Afric. d’Envr. d’Agric. 2018, 1, 25–29. [Google Scholar]
- Hancock, J.F. Plant Evolution and the Origin of Crop Species, 2nd ed.; CABI Publishing: Wallingford, UK, 2004. [Google Scholar]
- Todou, G.; Coppens, G.; Joly, H.I.; Akoa, A. Climatic Niche of Dacryodes edulis (G. Don) H. J. Lam (Burseraceae), a Semi-Domesticated Fruit Tree Native to Central Africa. J. Ecol. Nat. Environ. 2013, 5, 231–240. [Google Scholar] [CrossRef]
- Onana, J.M. A Synoptic Revision of Dacryodes (Burseraceae) in Africa, with a New Species from Central Africa. Kew Bull. 2008, 63, 385–400. [Google Scholar] [CrossRef]
- Kengue, J. Le Safoutier (Dacryodes edulis (G.Don) H. J. Lam) Premières Données Sur la Morphologie et la Biologie. Ph.D. Thesis, Université de Yaoundé, Yaoundé, Cameroun, 1990. [Google Scholar]
- Kengue, J.; Fohouo, F.N.T.; Adewusi, H.G. Towards the improvement of Safou (Dacryodes edulis (G.Don) H. J. Lam) population variation and reproductive biology. For. Trees Livelihoods 2012, 12, 73–84. [Google Scholar] [CrossRef]
- Makueti, J.T.; Tchoundjeu, Z.; Kalinganire, A.; Nkongmeneck, B.A.; Kouodiekong, L.; Asaah, E.; Tsobeng, A. Morphological Traits of Control-Pollinated Fruits in African Plum (Dacryodes Edulis (G.Don).Lam.) Using Multivariate Statistical Techniques. Int. J. Agron. Agric. Res. 2012, 2, 1–17. [Google Scholar]
- Waruhiu, A.N.; Kengue, J.; Atangana, A.R.; Tchoundjeu, Z.; Leakey, R.R.B. Domestication of Dacryodes edulis: 2. Phenotypic Variation of Fruit Traits in 200 Trees from Four Populations in the Humid Lowlands of Cameroon. J. Food Agric. Environ. 2004, 2, 340–346. [Google Scholar]
- Rimlinger, A.; Duminil, J.; Lemoine, T.; Avana, M.L.; Chakocha, A.; Gakwavu, A.; Mboujda, F.; Tsogo, M.; Elias, M.; Carrière, S.M. Shifting Perceptions, Preferences and Practices in the African Fruit Trade: The Case of African Plum (Dacryodes edulis) in Different Cultural and Urbanization Contexts in Cameroon. J. Ethnobiol. Ethnomed. 2021, 17, 65. [Google Scholar] [CrossRef] [PubMed]
- Ondo-Azi, A.S.; Missang, C.E.; Nsikabaka, S.; Silou, T.; Chalchat, J.C. Variation in Physicochemical and Morphological Characteristics of Safou (Dacryodes edulis (G. Don) H.J.Lam) Fruits: Classification and Identification of Elite Trees for Industrial Exploitation. J. Food Agric. Environ. 2014, 12, 212–217. [Google Scholar]
- Rimlinger, A.; Carrière, S.M.; Avana, M.L.; Nguegang, A.; Duminil, J. The Influence of Farmers’ Strategies on Local Practices, Knowledge, and Varietal Diversity of the Safou Tree (Dacryodes edulis) in Western Cameroon. Econ. Bot. 2019, 73, 249–264. [Google Scholar] [CrossRef]
- Leakey, R.R.B.; Tchoundjeu, Z.; Smith, R.I.; Munro, R.C.; Fondoun, J.; Kengue, J.; Anegbeh, P.O.; Atangana, A.R.; Waruhiu, N.; Asaah, E.; et al. Evidence Ttat Subsistence Farmers Have Domesticated Indigenous Fruits (Dacryodes edulis and Irvingia gabonensis) in Cameroon and Nigeria. Agrofor. Syst. 2004, 60, 101–111. [Google Scholar] [CrossRef]
- Ferrer, M.M.; Tapia-Gómez, C.A.; Estrada-Medina, H.; del Ruenes-Morales, M.R.; Montañez-Escalante, P.I.; Jiménez-Osornio, J.J. Growing Out of the Tropical Forests: Gene Flow of Native Mesoamerican Trees Among Forest and Mayan Homegardens. Front. Ecol. Evol. 2021, 9, 628765. [Google Scholar] [CrossRef]
- Gros-Balthazard, M.; Newton, C.; Ivorra, S.; Pierre, M.H.; Pintaud, J.C.; Terral, J.F. The Domestication Syndrome in Phoenix dactylifera Seeds: Toward the Identification of Wild Date Palm Populations. PLoS ONE 2016, 11, e0152394. [Google Scholar] [CrossRef]
- Arellano, E.; Casas, A. Morphological Variation and Domestication of Escontria chiotilla (Cactaceae) under Silvicultural Management in the Tehuacán Valley, Central Mexico. Genet. Resour. Crop Evol. 2003, 50, 439–453. [Google Scholar] [CrossRef]
- Chávez-Pesqueira, M.; Núñez-Farfán, J. Domestication and Genetics of Papaya: A Review. Front. Ecol. Evol. 2017, 5, 155. [Google Scholar] [CrossRef]
- Cornille, A.; Giraud, T.; Smulders, M.J.M.; Roldán-Ruiz, I.; Gladieux, P. The Domestication and Evolutionary Ecology of Apples. Trends Genet. 2014, 30, 57. [Google Scholar] [CrossRef]
- Hannachi, H.; Breton, C.; Msallem, M.; Ben El Hadj, S.; El Gazzah, M.; Bervillé, A. Differences between Native and Introduced Olive Cultivars as Revealed by Morphology of Drupes, Oil Composition and SSR Polymorphisms: A Case Study in Tunisia. Sci. Hortic. 2008, 116, 280–290. [Google Scholar] [CrossRef]
- Purugganan, M.D.; Fuller, D.Q. Archaeological Data Reveal Slow Rates of Evolution during Plant Domestication. Evolution 2011, 65, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Purugganan, M.D.; Fuller, D.Q. The Nature of Selection during Plant Domestication. Nature 2009, 457, 843–848. [Google Scholar] [CrossRef] [PubMed]
- Guillén, S.; Benítez, J.; Martínez-Ramos, M.; Casas, A. Seed Germination of Wild, in Situ-Managed, and Cultivated Populations of Columnar Cacti in the Tehuacán-Cuicatlán Valley, Mexico. J. Arid Environ. 2009, 73, 407–413. [Google Scholar] [CrossRef]
- Kadji, B.R.L.; Kone, F.M.T.; Sika, A.E.; Dabonne, S. Physico-Chemical Properties of Safou (Dacryodes edulis) Fruits Grown in Côte d’Ivoire. J. Appl. Biosci. 2016, 105, 10103–10110. [Google Scholar] [CrossRef]
- Kapseu, C.; Avouampo, E.; Djeumako, B. Oil Extraction from Dacryodes edulis (G.Don) H J. Lam Fruit. For. Trees Livelihoods 2002, 12, 97–104. [Google Scholar] [CrossRef]
- Mbofung, C.M.F.; Silou, T.; Mouragadja, I. Chemical Characterization of Safou (Dacryodes edulis) and Evaluation of Its Potential as an Ingredient in Nutritious Biscuits. For. Trees Livelihoods 2002, 12, 105–117. [Google Scholar] [CrossRef]
- Ondo-Azi, A.S.; Missang, C.E.; Silou, T. Classification of Dacryodes edulis (G. Don) H.J. Lam by Using Morphological and Physical Characteristics of the Fruits: A Statistical Approach. For. Trees Livelihoods 2009, 19, 99–109. [Google Scholar] [CrossRef]
- Silou, T. Characterization of Safou pulp Oil: Extraction, Assay and Analysis. In Oils and Fats for the Future: A Case Study: Safou (Dacryodes edulis) from the Congo Basin Countries in Africa; Nova Science Publishers, Inc.: New York, NY, USA, 2012; pp. 43–74. [Google Scholar]
- Tshiombe, M.; Masimango, N.; Silou, T.; Diawidi, D. Influence of Taste on Safou Oil Productivity. Int. J. Agric. Food Sci. 2014, 4, 50–53. [Google Scholar]
- Zohary, D.; Spiegel-Roy, P.; Science, S.; Series, N.; Jan, N. Beginnings of Fruit Growing in the Old World. Olive, grape, date, and fig emerge as important Bronze Age additions to grain agriculture in the Near East. Science 1975, 187, 319–327. [Google Scholar] [CrossRef]
- Besnard, G.; Terral, J.F.; Cornille, A. On the Origins and Domestication of the Olive: A Review and Perspectives. Ann. Bot. 2018, 121, 385–403. [Google Scholar] [CrossRef] [PubMed]
- Omondi, S.F.; Odee, D.W.; Ongamo, G.O.; Kanya, J.I.; Khasa, D.P. Synchrony in Leafing, Flowering, and Fruiting Phenology of Senegalia Senegal within Lake Baringo Woodland, Kenya: Implication for Conservation and Tree Improvement. Int. J. of For. Res. 2016, 2016, 6904834. [Google Scholar]
- Leakey, R.R.B.; Atangana, A.A.; Kengni, E.; Waruhiu, A.N.; Usoro, C.; Anegbeh, P.O.; Tchoundjeu, Z. Domestication of Dacryodes edulis in West and Central Africa: Characterization of Genetic Variation. For. Trees Livelihoods 2002, 12, 57–71. [Google Scholar] [CrossRef]
- Anegbeh, P.O.; Ukafor, V.; Usoro, C.; Tchoundjeu, Z. Domestication of Dacryodes edulis: 1. Phenotypic Variation of Fruit Traits from 100 Trees in Southeast Nigeria. New For. 2005, 29, 149–160. [Google Scholar] [CrossRef]
- Ndindeng, S.A.; Bella-Manga, K.J.; Talle, L.D. Quality Standards for Dacryodes edulis (Safou). Res. Rep. Int. Cent. Underutil. Crops 2008, 5, 4–7. [Google Scholar]
- Firestone, D.; Horwitz, W. IUPAC Gas Chromatographic Method for Determination of Fatty Acid Composition: Collaborative Study. J. Assoc. Off. Anal. Chem. 1979, 62, 709–721. [Google Scholar] [CrossRef]
- Laffargue, A.; de Kochko, A.; Dussert, S. Development of Solid-Phase Extraction and Methylation Procedures to Analyze Free Fatty Acids in Lipid-Rich Seeds. Plant Physiol. Biochem. 2007, 45, 250–257. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing; R Development Core Team: Vienna, Austria, 2008; ISBN 3-900051-07-0. Available online: Http://www.R-Project.Org/ (accessed on 20 January 2021).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivated Individuals | Wild Individuals | |||||
|---|---|---|---|---|---|---|
| Sexual Type | Eseka (N = 30) | Makak (N = 16) | Total (N = 46) | Messock (N = 28) | Somalomo (N = 10) | Total (N = 38) |
| Male | 5 (16.7%) | 3 (18.7%) | 8 (17.4%) | 11 (39.3%) | 7 (70.0%) | 18 (47.4%) |
| Female | 12 (40.0%) | 4 (25.0%) | 16 (34.8%) | 7 (25.0%) | 1 (10.0%) | 8 (21%) |
| Male-hermaphrodite | 5 (16.7%) | 7 (43.8%) | 12 (26.1%) | 10 (35.7%) | 2 (20.0%) | 12 (31.6%) |
| Hermaphrodite | 8 (26.6%) | 2 (12.5%) | 10 (21.7%) | 0 | 0 | 0 |
| Factors | Modalities | N | Flower Length (mm) | Stamen Length (mm) | Peduncle Length (mm) | Ovary Length (mm) |
|---|---|---|---|---|---|---|
| Male flowers | ||||||
| Status of individuals | Cultivated | 240 | 5.36 ± 1.34 a | 2.05 ± 0.59 a | 1.41 ± 0.68 a | 0.05 ± 0.19 a |
| Wild | 540 | 5.52 ± 1.42 a | 1.90 ± 0.58 b | 1.68 ± 0.78 b | 0.05 ± 0.18 a | |
| Sites | Eseka (C) | 150 | 5.27 ± 1.12 a | 2.06 ± 0.57 a | 1.25 ± 0.51 c | 0.07 ± 0.24 a |
| Makak (C) | 90 | 5.51 ± 1.63 a | 2.03 ± 0.62 a | 1.68 ± 0.83 ab | 0.00 ± 0.0 b | |
| Messock (W) | 330 | 5.21 ± 1.14 a | 2.03 ± 0.51 a | 1.59 ± 0.71 b | 0.08 ± 0.22 a | |
| Somalomo (W) | 210 | 6.01 ± 1.66 b | 1.69 ± 0.62 b | 1.82 ± 0.87 a | 0.00 ± 0.00 b | |
| Female flowers | ||||||
| Status of individuals | Cultivated | 530 | 7.40 ± 1.42 a | 1.47 ± 0.77 a | 2.86 ± 1.02 a | 2.36 ± 0.70 a |
| Wild | 270 | 7.43 ± 1.42 a | 1.36 ± 0.5 b | 2.81 ± 0.96 a | 2.23 ± 0.63 b | |
| Sites | Eseka (C) | 350 | 7.54 ± 1.51 a | 1.53 ± 0.86 a | 3.01 ± 1.01 a | 2.46 ± 0.63 a |
| Makak (C) | 120 | 7.25 ± 1.83 ab | 1.23 ± 0.58 b | 2.78 ± 1.13 a | 2.11 ± 0.91 b | |
| Messock (C) | 60 | 6.81 ± 1.04 b | 1.57 ± 0.35 a | 2.23 ± 0.54 b | 2.31 ± 0.40 ab | |
| Messock (W) | 240 | 7.47 ± 1.75 a | 1.43 ± 0.5 ab | 2.85 ± 0.96 a | 2.34 ± 0.56 a | |
| Somalomo (W) | 30 | 7.13 ± 1.22 ab | 0.85 ± 0.45 c | 2.50 ± 0.90 ab | 1.38 ± 0.44 c | |
| Status of Individuals | Morphometric Parameters | ||||||
|---|---|---|---|---|---|---|---|
| Fruits | N | Mass (g) | Length (mm) | Width (mm) | Thickness (mm) | Mass of pulp (g) | |
| Cultivated | 1896 | 49.10 ± 17.52 a | 68.24 ± 14.0 a | 36.20 ± 7.74 a | 5.70 ± 1.42 a | 36.60 ± 15.70 a | |
| Wild | 650 | 15.60 ± 7.30 b | 42.04 ± 8.41 b | 23.00 ± 4.41 b | 2.72 ± 1.30 b | 9.72 ± 5.01b | |
| Seeds | N | Mass (g) | Length (mm) | Width (mm) | N cotyledon lobes | ||
| Cultivated | 1896 | 12.32 ± 4.07 a | 40.60 ± 12.80 a | 21.56 ± 5.95 a | 9.6 ± 1.04 a | ||
| Wild | 650 | 6.05 ± 2.63 b | 32.30 ± 5.83 b | 15.40 ± 3.45 b | 9.34 ± 1.46 b | ||
| Status of Individuals | |||
|---|---|---|---|
| Compounds | Cultivated | Wild | F Value |
| %Oil | 67.10 ± 11.80 | 64.30 ± 11.30 | 0.8 |
| %Palmitic acid | 39.62 ± 4.05 | 41.96 ± 2.64 | 6.5 * |
| %Palmitoleic acid | 0.17 ± 0.07 | 0.15 ± 0.05 | 0.9 |
| %Stearic acid | 2.64 ± 0.75 | 2.46 ± 0.75 | 0.6 |
| %Oleic acid | 32.52 ± 6.15 | 32.62 ± 6.04 | 0.01 |
| %Vaccenic acid | 0.95 ± 0.25 | 0.98 ± 0.34 | 0.2 |
| %Linoleic acid | 23.11 ± 20.76 | 20.76 ± 5.42 | 3.1 |
| %Linolenic acid | 0.82 ± 0.25 | 0.91 ± 0.17 | 1.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mboujda, F.M.M.; Avana-Tientcheu, M.-L.; Momo, S.T.; Ntongme, A.M.; Vaissayre, V.; Azandi, L.N.; Dussert, S.; Womeni, H.; Onana, J.-M.; Sonké, B.; et al. Domestication Syndrome in Dacryodes edulis (Burseraceae): Comparison of Morphological and Biochemical Traits between Wild and Cultivated Populations. Plants 2022, 11, 2496. https://doi.org/10.3390/plants11192496
Mboujda FMM, Avana-Tientcheu M-L, Momo ST, Ntongme AM, Vaissayre V, Azandi LN, Dussert S, Womeni H, Onana J-M, Sonké B, et al. Domestication Syndrome in Dacryodes edulis (Burseraceae): Comparison of Morphological and Biochemical Traits between Wild and Cultivated Populations. Plants. 2022; 11(19):2496. https://doi.org/10.3390/plants11192496
Chicago/Turabian StyleMboujda, Franca Marcelle Meguem, Marie-Louise Avana-Tientcheu, Stéphane Takoudjou Momo, Alix Mboukap Ntongme, Virginie Vaissayre, Laura N. Azandi, Stéphane Dussert, Hilaire Womeni, Jean-Michel Onana, Bonaventure Sonké, and et al. 2022. "Domestication Syndrome in Dacryodes edulis (Burseraceae): Comparison of Morphological and Biochemical Traits between Wild and Cultivated Populations" Plants 11, no. 19: 2496. https://doi.org/10.3390/plants11192496
APA StyleMboujda, F. M. M., Avana-Tientcheu, M. -L., Momo, S. T., Ntongme, A. M., Vaissayre, V., Azandi, L. N., Dussert, S., Womeni, H., Onana, J. -M., Sonké, B., Tankou, C., & Duminil, J. (2022). Domestication Syndrome in Dacryodes edulis (Burseraceae): Comparison of Morphological and Biochemical Traits between Wild and Cultivated Populations. Plants, 11(19), 2496. https://doi.org/10.3390/plants11192496