
Evolutionary Aspects of Hypericin Productivity and Endogenous Phytohormone Pools Evidenced in Hypericum Species In Vitro Culture Model

 ,
,  ,
,  , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
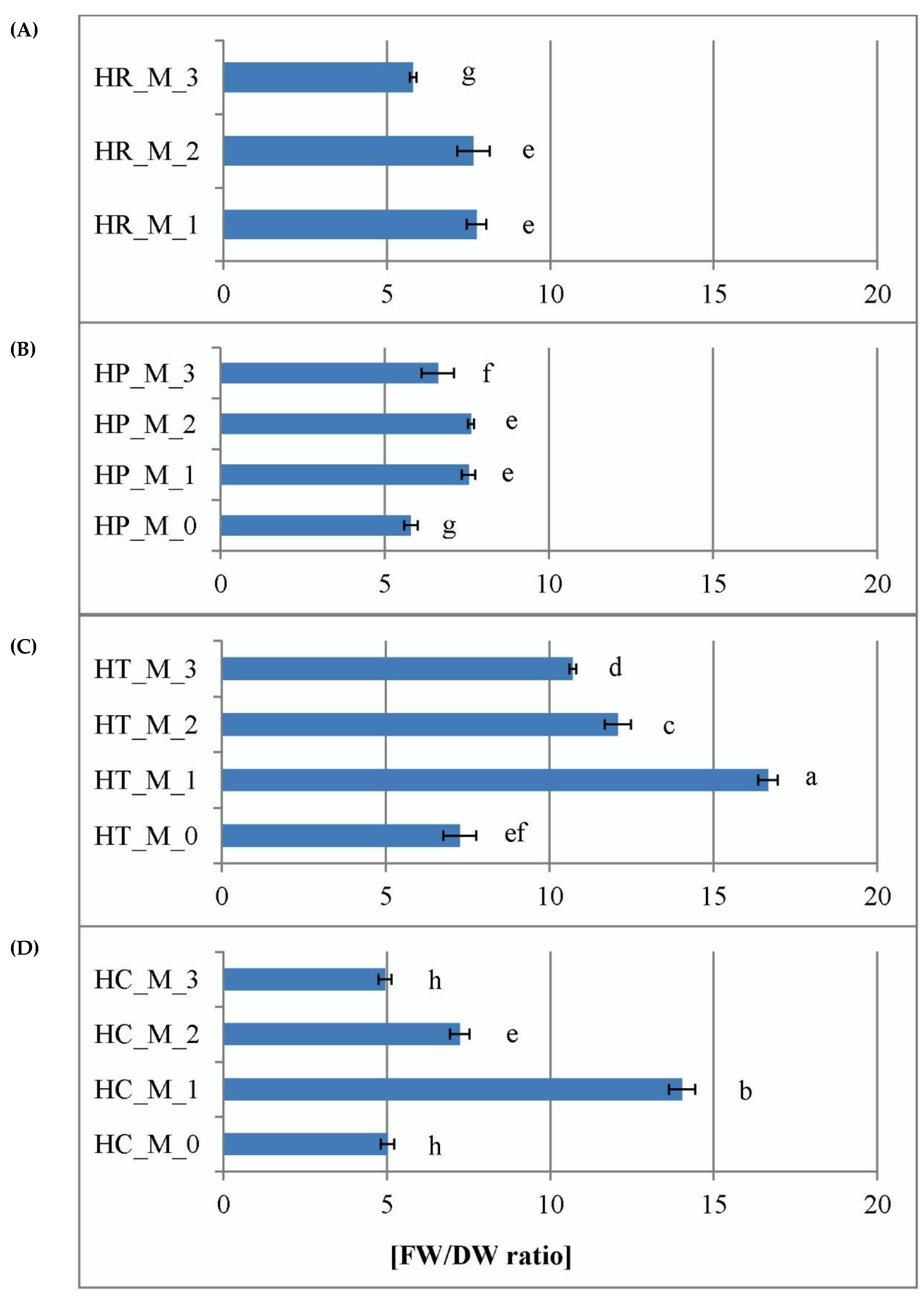
2.1. Effect of Plant Growth Regulators on In Vitro Culture Development
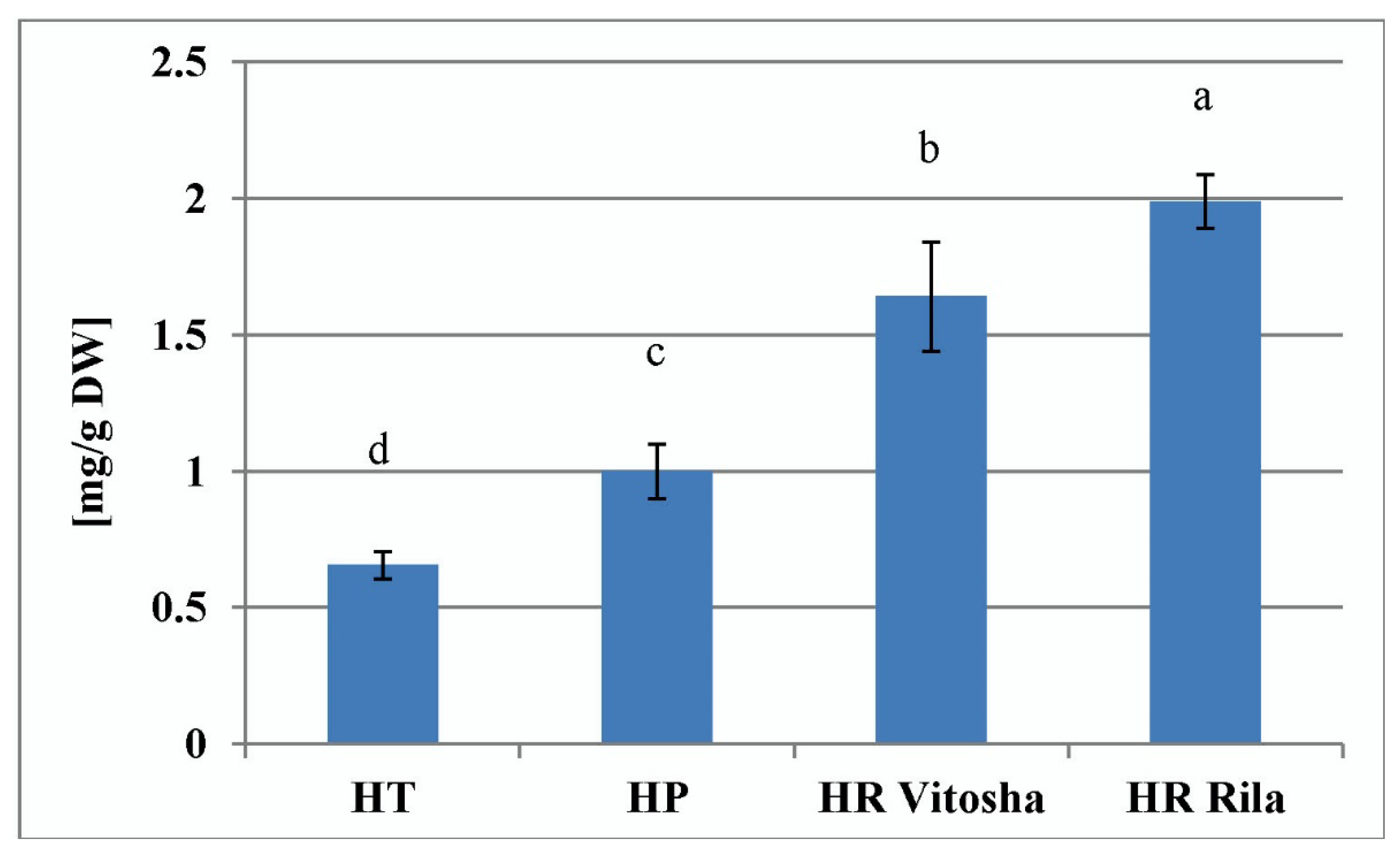
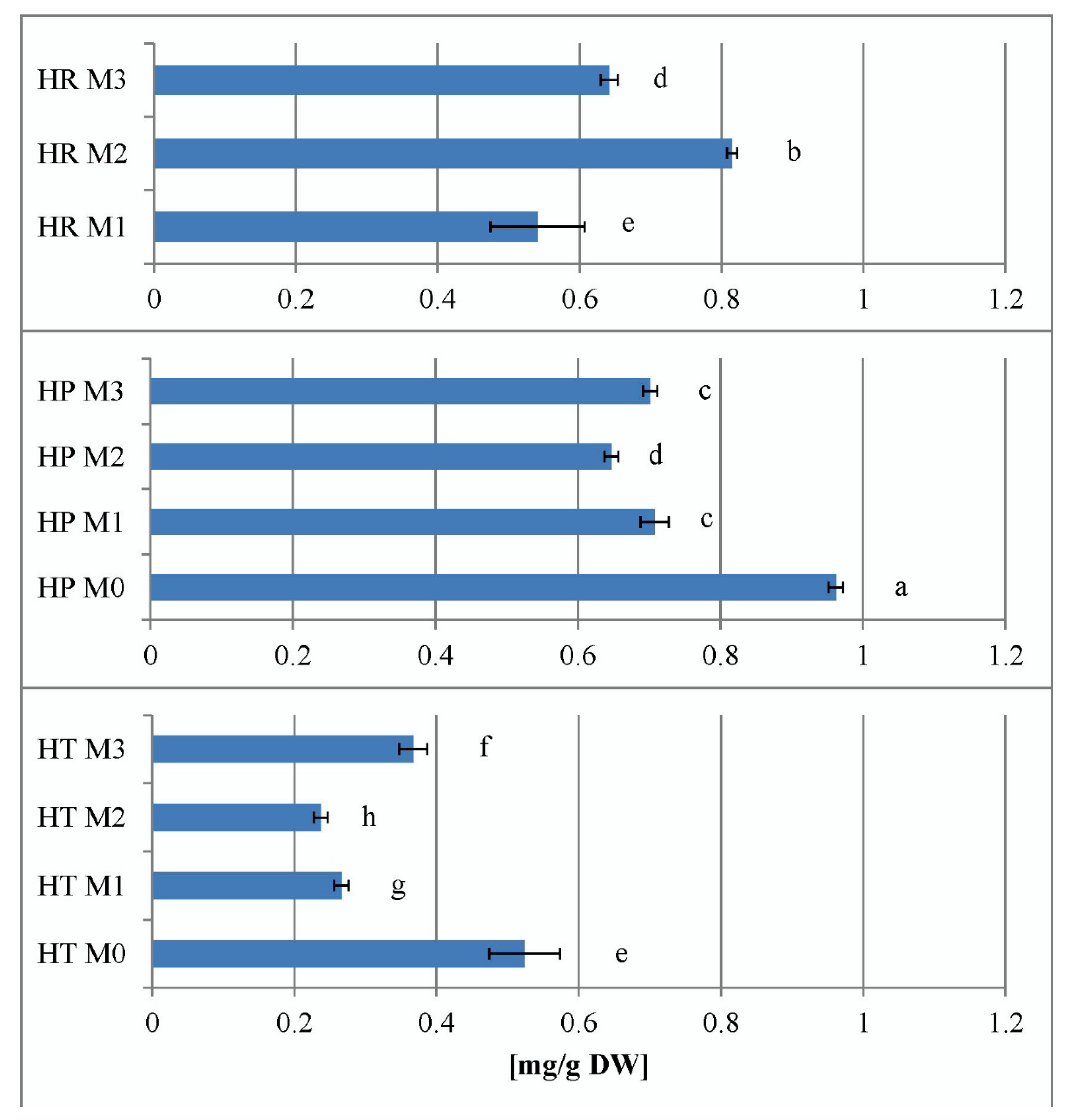
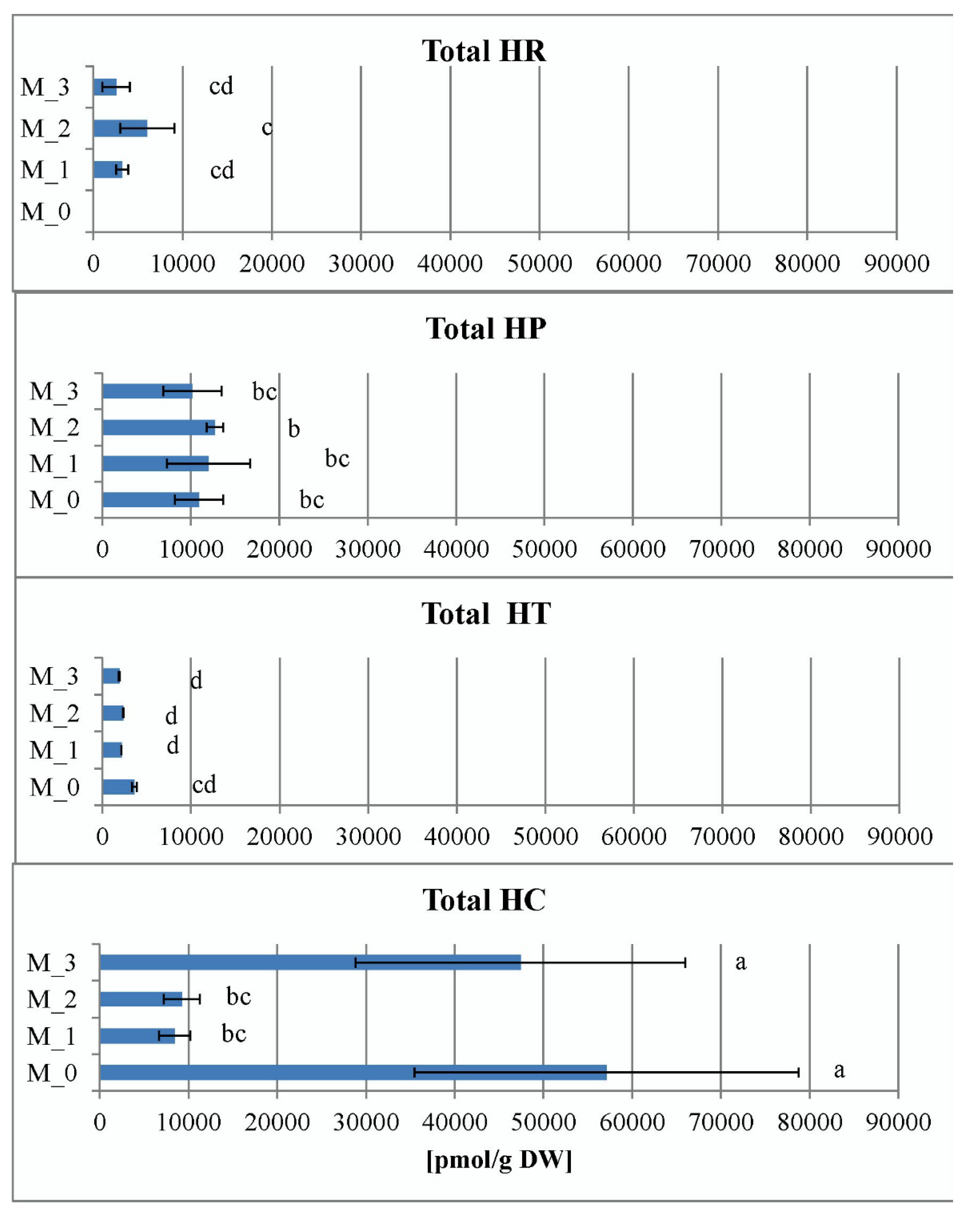
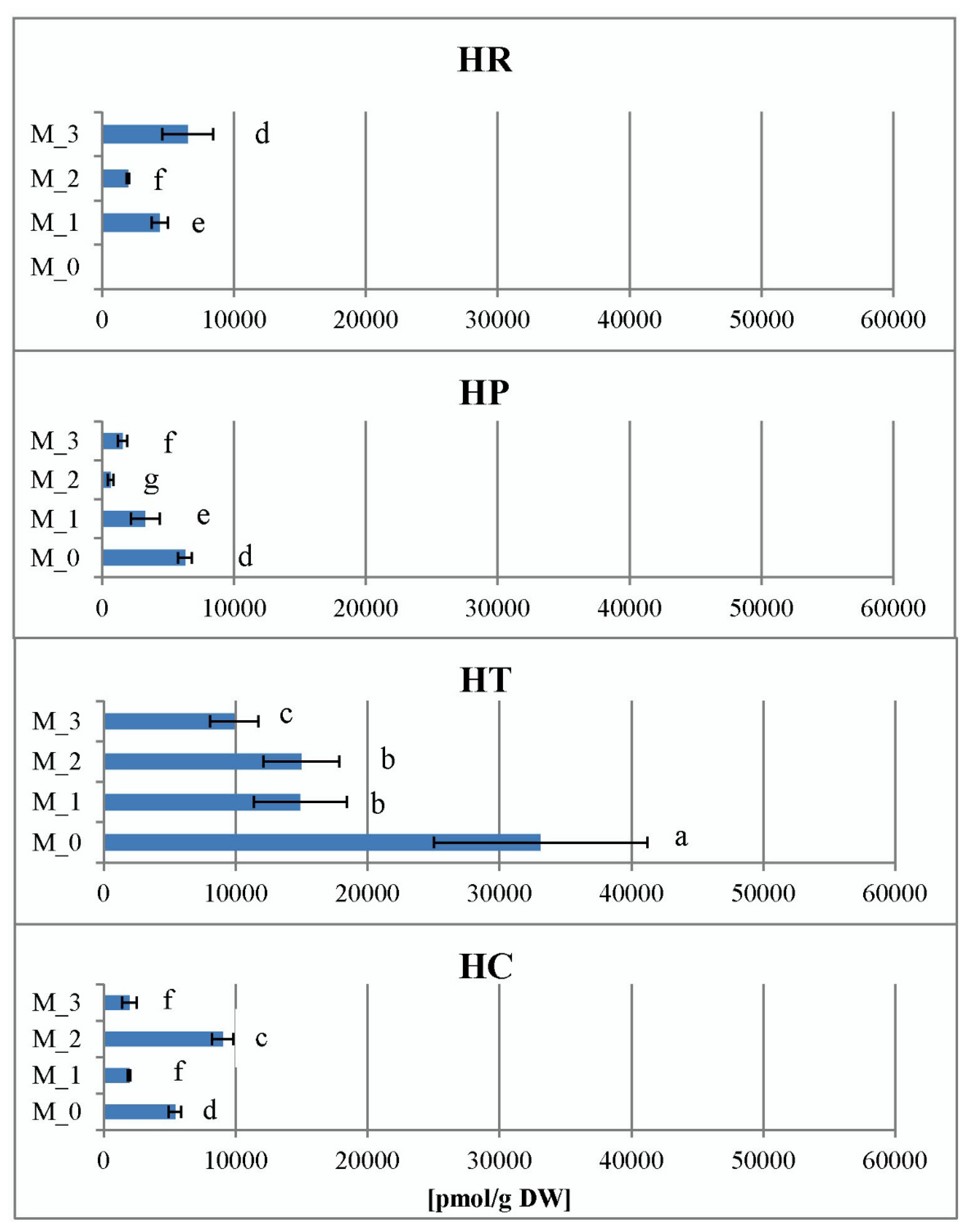
2.2. Total Hypericins Content of Field and In Vitro Collected Samples
2.3. Effect of Plant Growth Regulators Treatments on Endogenous Phytohormone Levels In Vitro
2.3.1. Levels of Abscisic Acid and Its Conjugates In Vitro
2.3.2. Levels of Benzoic and Salicylic Acid In Vitro
2.3.3. Endogenous Levels of Jasmonates In Vitro
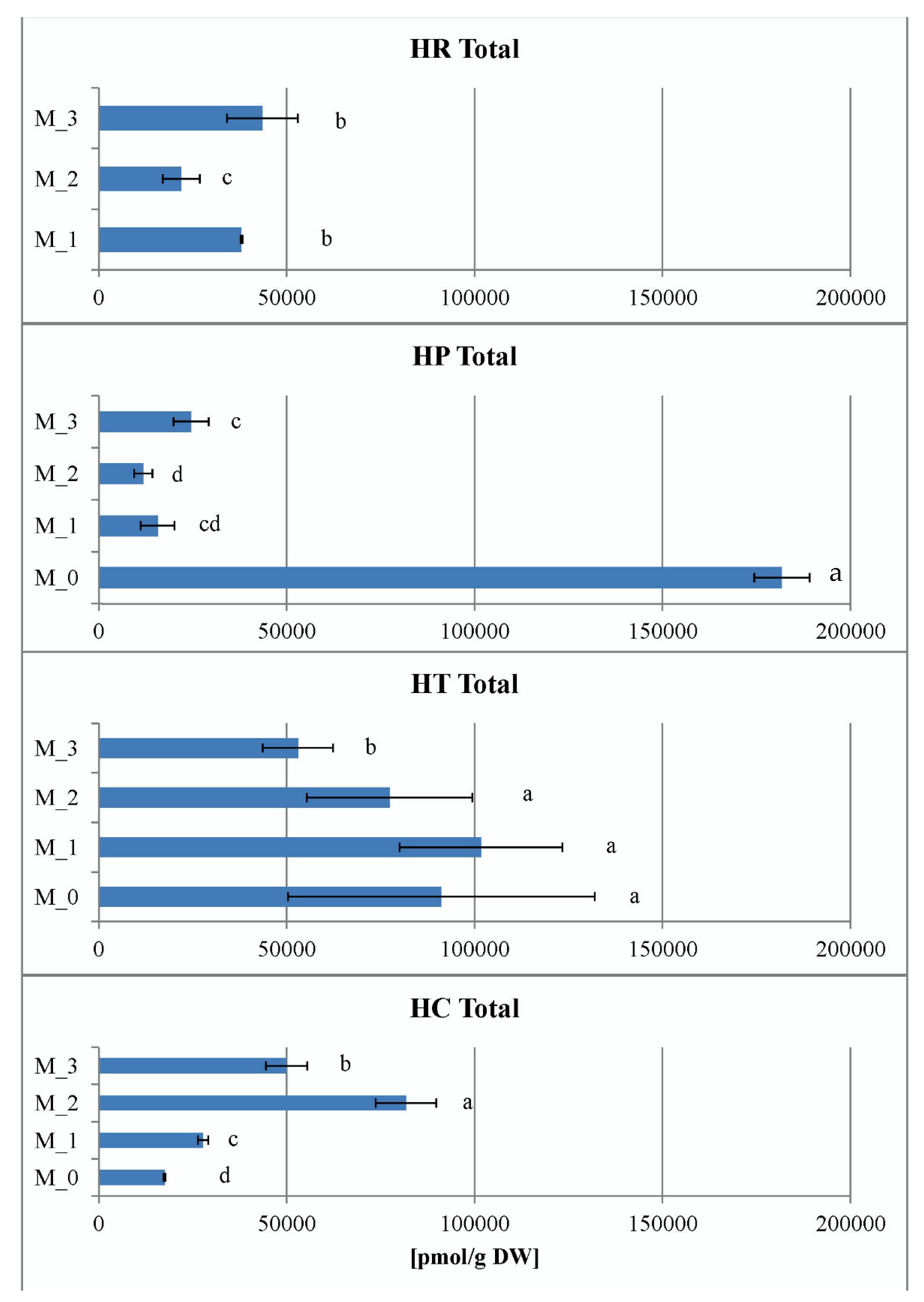
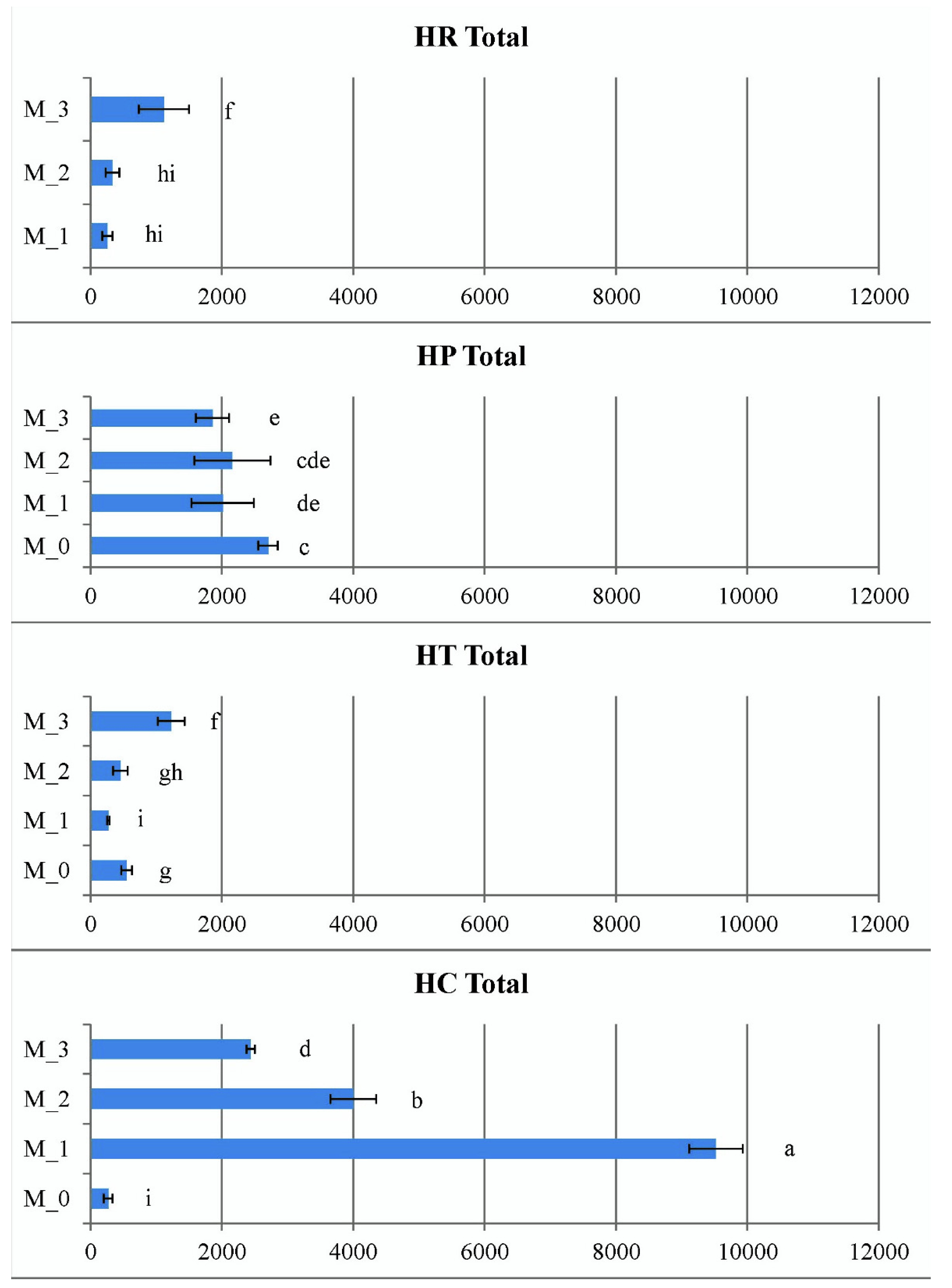
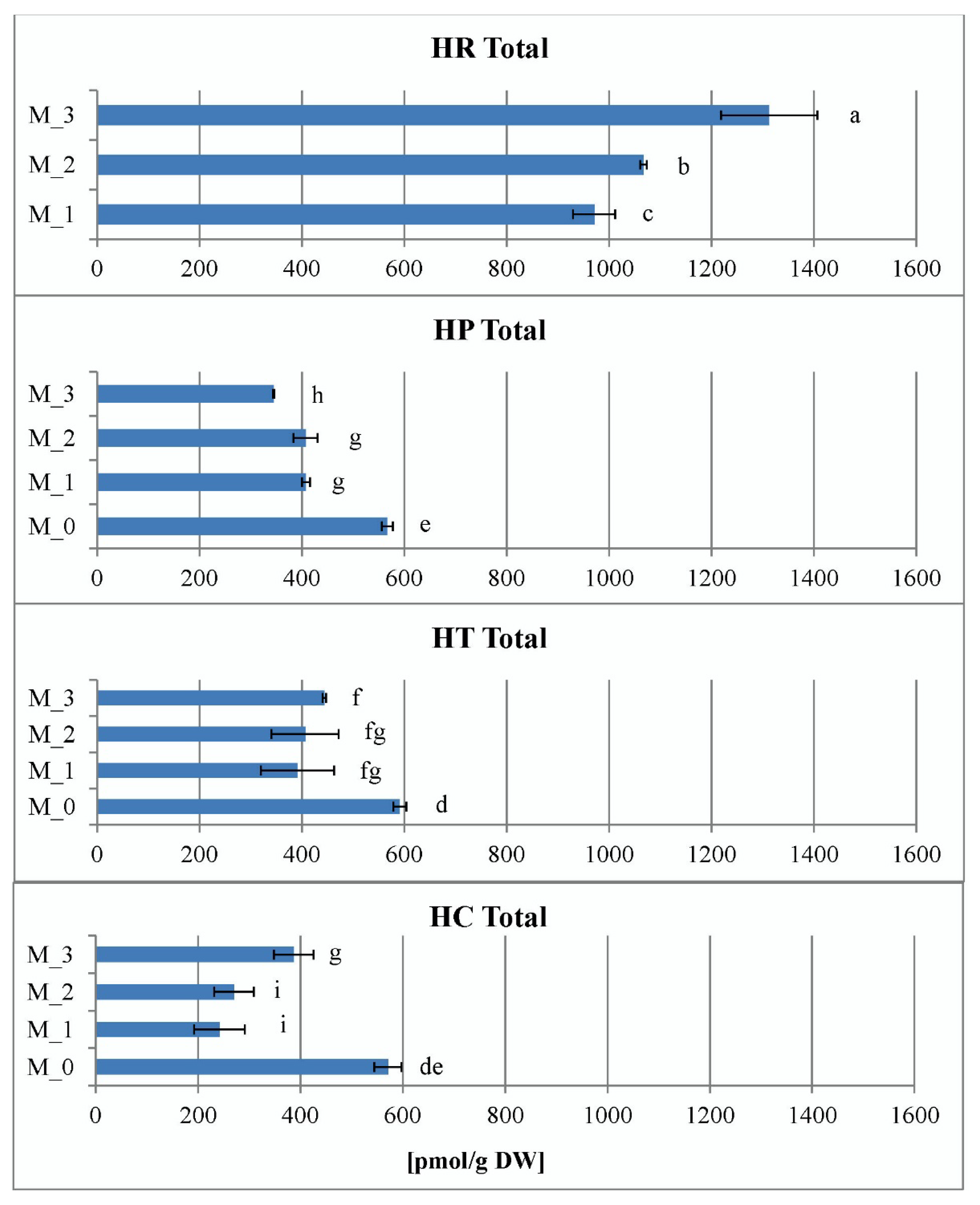
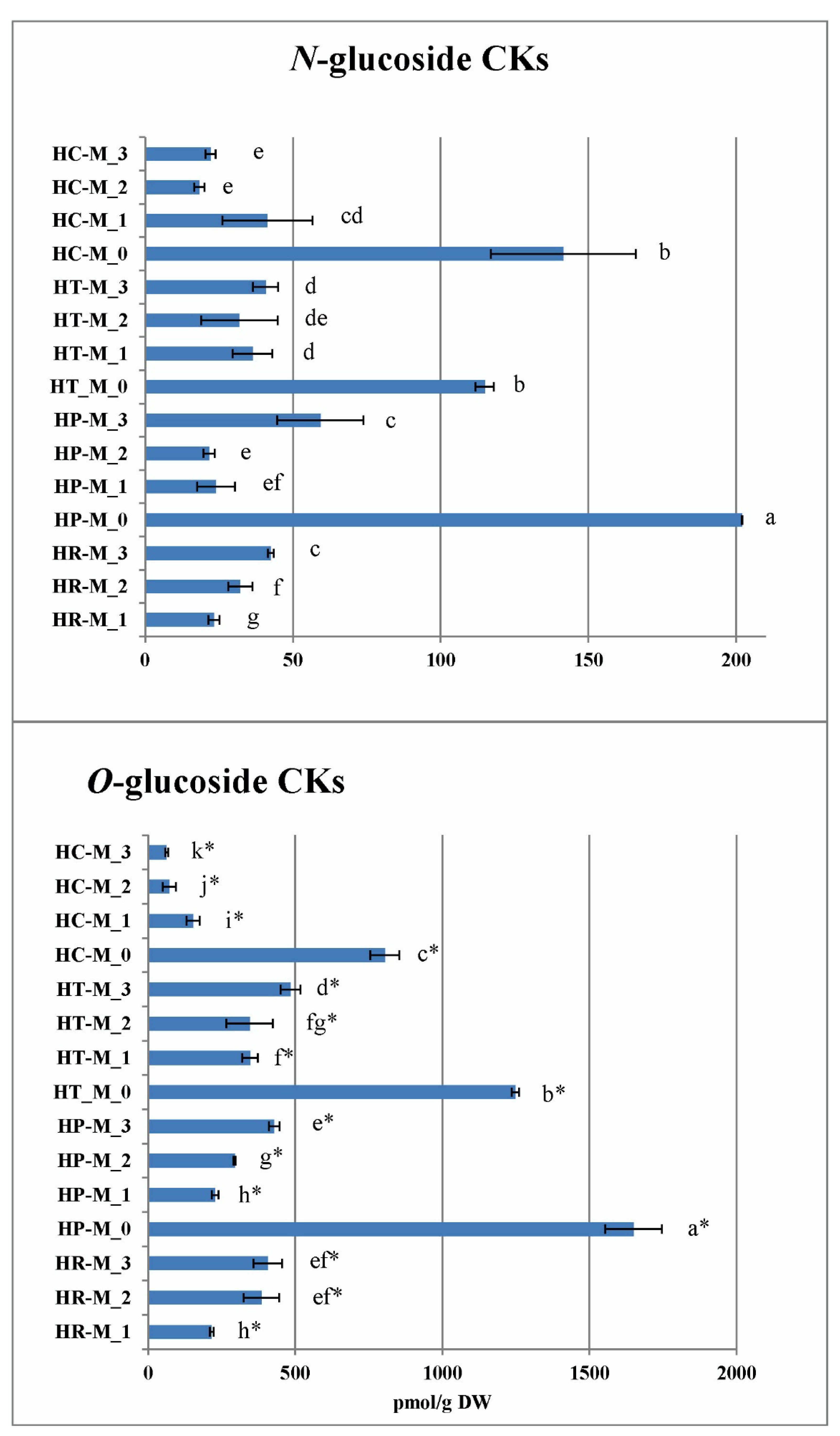
2.3.4. Contents of Endogenous Cytokinins In Vitro
2.3.5. Contents of Phenylacetic Acid In Vitro
3. Materials and Methods
3.1. Plant Material
3.2. Tissue Culture Initiation
3.3. Treatments with Plant Growth Regulators
3.4. Estimation of Hydricity of the Plant Tissues In Vitro
3.5. Analysis of the Total Amount of Hypericin
3.5.1. Extraction of Plant Material
3.5.2. Determination of Total Hypericins
3.6. Endogenous Phytohormone Levels Analyses
3.7. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Danova, K. Potential of the Balkan flora as a source of prospective Hypericum genotypes for the conventional and biotechnological delivery of phytopharmaceuticals. In Hypericum: Botanical Sources, Medical Properties and Health Effects; Davis, H.R., Ed.; Series Plant Science Research and Practices; Nova Science Publishers: New York, NY, USA, 2015; Chapter 2; pp. 19–52. ISBN 978-163482750-8/978-163482701-0. [Google Scholar]
- Nahrstedt, A.; Butterweck, V. Biologically active and other chemical constituents of the herb of Hypericum perforatum L. Pharmacopsychiatry 1997, 30, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Vuko, E.; Dunkić, V.; Ruščić, M.; Nazlić, M.; Mandić, N.; Soldo, B.; Šprung, M.; Fredotović, Ž. Chemical Composition and New Biological Activities of Essential Oil and Hydrosol of Hypericum perforatum L. ssp. veronense (Schrank) H. Lindb. Plants 2021, 10, 1014. [Google Scholar] [CrossRef] [PubMed]
- Nahrstedt, A.; Butterweck, V. Lessons learned from herbal medicinal products: The example of St. John’s wort. J. Nat. Prod. 2010, 73, 1015–1021. [Google Scholar] [CrossRef] [PubMed]
- Barnes, J.; Anderson, L.A.; Phillipson, J.D. St John’s wort (Hypericum perforatum L.): A review of its chemistry, pharmacology and clinical properties. J. Pharm. Pharmacol. 2001, 53, 583–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saddiqe, Z.; Naeem, I.; Maimoona, A. A review of the antibacterial activity of Hypericum perforatum L. J. Ethnopharmacol. 2010, 131, 511–521. [Google Scholar] [CrossRef] [PubMed]
- Süntar, I.P.; Akkol, E.K.; Yilmazer, D.; Baykal, T.; Kirmizibekmez, H.; Alper, M.; Yeşilada, E. Investigations on the in vivo wound healing potential of Hypericum perforatum L. J. Ethnopharmacol. 2010, 127, 468–477. [Google Scholar] [CrossRef] [PubMed]
- Crockett, S.L.; Robson, N.K.B. Taxonomy and chemotaxonomy of the genus Hypericum. In Medicinal and Aromatic Plant Science and Biotechnology; Global Science Books: Bexhill, UK, 2011; Volume 5, pp. 1–13. [Google Scholar] [PubMed] [Green Version]
- Karioti, A.; Bilia, A.R. Hypericins as potential leads for new therapeutics. Int. J. Mol. Sci. 2010, 11, 562–594. [Google Scholar] [CrossRef] [Green Version]
- Vandenbogaerde, A.L.; Kamuhabwa, A.; Delaey, E.; Himpens, B.E.; Merlevede, W.J.; de Witte, P.A. Photocytotoxic effect of pseudohypericin versus hypericin. J. Photochem. Photobiol. B 1998, 45, 87–94. [Google Scholar] [CrossRef]
- Martinez-Poveda, B.; Quesada, A.R.; Medina, M.A. Hypericin in the dark inhibits key steps of angiogenesis in vitro. Eur. J. Pharmacol. 2005, 516, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Blank, M.; Lavie, G.; Mandel, M.; Hazan, S.; Orenstein, A.; Meruelo, D.; Keisari, Y. Antimetastatic activity of the photodynamic agent hypericin in the dark. Int. J. Cancer 2004, 111, 596–603. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, H.; Zou, M.; Oerlemans, R.; Shao, C.; Ren, Y.; Zhang, R.; Huang, X.; Li, G.; Cong, Y. Hypericin inhibit alpha-coronavirus replication by targeting 3CL protease. Viruses 2021, 13, 1825. [Google Scholar] [CrossRef] [PubMed]
- Shivanika, C.; Deepak, K.S.; Venkataraghavan, R.; Pawan, T.; Sumitha, A.; Brindha, D.P. Molecular docking, validation, dynamics simulations, and pharmacokinetic prediction of natural compounds against the SARS-CoV-2 main-protease. J. Biomol. Struct. Dyn. 2022, 40, 585–611. [Google Scholar] [CrossRef]
- Islam, R.; Parves, R.; Paul, A.S.; Uddin, N.; Rahman, S.; Al Mamun, A.; Hossain, N.; Ali, A.; Halim, M.A. A molecular modeling approach to identify effective antiviral phytochemicals against the main protease of SARS-CoV-2. J. Biomol. Struct. Dyn. 2020, 39, 3213–3224. [Google Scholar] [CrossRef] [PubMed]
- Saravanan, K.M.; Zhang, H.; Senthil, R.; Vijayakumar, K.K.; Sounderrajan, V.; Wei, Y.; Shakila, H. Structural basis for the inhibition of SARS-CoV2 main protease by Indian medicinal plant-derived antiviral compounds. J. Biomol. Struct. Dyn. 2022, 40, 1970–1978. [Google Scholar] [CrossRef] [PubMed]
- Carrubba, A.; Lazzara, S.; Giovino, A.; Ruberto, G.; Napoli, E. Content variability of bioactive secondary metabolites in Hypericum perforatum L. Phytochem. Lett. 2021, 46, 71–78. [Google Scholar] [CrossRef]
- Kitanov, G.M. Hypericin and pseudohypericin in some Hypericum species. Biochem. Syst. Ecol. 2001, 29, 171–178. [Google Scholar] [CrossRef]
- Smelcerovic, A.; Verma, V.; Spiteller, M.; Ahmad, S.M.; Puri, S.C.; Qazi, G.N. Phytochemical analysis and genetic characterization of six Hypericum species from Serbia. Phytochemistry 2006, 67, 171–177. [Google Scholar] [CrossRef]
- Bruni, R.; Sacchetti, G. Factors affecting polyphenol biosynthesis in wild and field grown St. John’s Wort (Hypericum perforatum L. Hypericaceae/Guttiferae). Molecules 2009, 14, 682–725. [Google Scholar] [CrossRef] [Green Version]
- Yordanov, D.; Kojuharov, S. Guttiferae Family. In Flora of the PR of Bulgaria; Bulgarian Academy of Sciences: Sofia, Bulgaria, 1970; Volume 4, p. 227. [Google Scholar]
- Robson, N.K.B. Studies in the genus Hypericum L. (Guttiferae). 1. Infrageneric classification. Bull. Br. Mus. (Nat. Hist.) 1977, 5, 325, ISSN: 0068-2292. [Google Scholar]
- Anchev, M.; Apostolova, I.; Assyov, B.; Bancheva, S.; Denchev, C.M.; Dimitrov, D.; Dimitrova, D.; Evstatieva, L.; Genova, E.; Georgiev, V.; et al. Red List of Bulgarian Vascular Plants. Phytol. Balc. 2009, 15, 63–94. [Google Scholar]
- Danova, K.; Motyka, V.; Dobrev, P. Could evolutionary factors affect endogenous cytokinin pools in Hypericum species in vitro? Annual of Sofia Universiti “St. Kliment Ohridski”; Faculty of Biology, Sofia Universiti “St. Kliment Ohridski”: Sofia, Bulgaria, 2021; Volume 106, pp. 12–19. [Google Scholar]
- Huang, W.; Ratkowsky, D.A.; Hui, C.; Wang, P.; Su, J.; Shi, P. Leaf Fresh Weight Versus Dry Weight: Which is Better for Describing the Scaling Relationship between Leaf Biomass and Leaf Area for Broad-Leaved Plants? Forests 2019, 10, 256. [Google Scholar] [CrossRef]
- Ciccarelli, D.; Andreucci, A.C.; Pagni, A.M. Translucent glands and secretory canals in Hypericum perforatum L. (Hyperiacaceae): Morphological, anatomical and histochemical studies during the course of ontogenesis. Ann. Bot. 2001, 88, 637–644. [Google Scholar] [CrossRef] [Green Version]
- Adam, P.; Arigoni, D.; Bacher, A.; Eisenreich, W. Biosynthesis of hyperforin in Hypericum perforatum. J. Med. Chem. 2002, 45, 4786–4793. [Google Scholar] [CrossRef] [PubMed]
- Soelberg, J.; Jørgensen, L.B.; Jäger, A.K. Hyperforin accumulates in the translucent glands of Hypericum perforatum. Ann. Bot. 2007, 99, 1097–1100. [Google Scholar] [CrossRef] [Green Version]
- Pradeep, M.; Franklin, G. Understanding the hypericin biosynthesis via reversible inhibition of dark gland development in Hypericum perforatum L. Ind. Crop. Prod. 2022, 182, 114876. [Google Scholar] [CrossRef]
- Zdunek, K.; Alfermann, A.W. Introduction of shoot organ cultures of Hypericum perforatum and formation of hypericin derivatives. Planta Med. 1992, 58, 621–622. [Google Scholar] [CrossRef]
- Čellárová, E.; Kimáková, K.; Brutovská, R. Multiple shoot formation and phenotypic changes of R0 regenerants in Hypericum perforatum L. Acta Biotechnol. 1992, 12, 445–452. [Google Scholar] [CrossRef]
- Shakya, P.; Marslin, G.; Siram, K.; Beerhues, L.; Franklin, G. Elicitation as a tool to improve the profiles of high-value secondary metabolites and pharmacological properties of Hypericum perforatum. J. Pharm. Pharmacol. 2019, 71, 70–82. [Google Scholar] [CrossRef] [Green Version]
- Coste, A.; Vlase, L.; Halmagyi, A.; Deliu, C.; Coldea, G. Effects of plant growth regulators and elicitors on production of secondary metabolites in shoot cultures of Hypericum hirsutum and Hypericum maculatum. Plant Cell Tissue Organ Cult. 2011, 106, 279–288. [Google Scholar] [CrossRef]
- Tusevski, O.; Stanoeva, J.P.; Markoska, E.; Brndevska, N.; Stefova, M.; Gadzovska Simic, S. Callus cultures of Hypericum perforatum L. a novel and efficient source for xanthone production. Plant Cell Tissue Organ Cult. 2016, 125, 309–319. [Google Scholar] [CrossRef]
- Božin, B.; Kladar, N.; Grujić, N.; Anačkov, G.; Samojlik, I.; Gavarić, N.; Conić, B.S. Impact of origin and biological source on chemical composition, anticholinesterase and antioxidant properties of some St. John’s wort species (Hypericum spp., Hypericaceae) from the Central Balkans. Molecules 2013, 18, 11733–11750. [Google Scholar] [CrossRef] [PubMed]
- Christmann, A.; Hoffmann, T.; Teplova, I.; Grill, E.; Müller, A. Generation of Active Pools of Abscisic Acid Revealed by In Vivo Imaging of Water-Stressed Arabidopsis. Plant Physiol. 2005, 137, 209–219. [Google Scholar] [CrossRef] [Green Version]
- Tallman, G. Are diurnal patterns of stomatal movement the result of alternating metabolism of endogenous guard cell ABA and accumulation of ABA delivered to the apoplast around guard cells by transpiration? J. Exp. Bot. 2004, 55, 1963–1976. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Cao, J.; He, J.; Chen, Q.; Li, X.; Yang, Y. Molecular Mechanism for the Regulation of ABA Homeostasis During Plant Development and Stress Responses. Int. J. Mol. Sci. 2018, 19, 3643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, D.M.; Liu, Z.; Chen, H.; Xue, J.T.; Yang, Y.; Chen, C.; Ye, L.M. Profiling the dynamics of abscisic acid and ABA-glucose ester after using the glucosyltransferase UGT71C5 to mediate abscisic acid homeostasis in Arabidopsis thaliana by HPLC-ESI-MS/MS. J. Pharm. Anal. 2014, 4, 190–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehmann, H.; Schutte, H.R. Abscisic acid metabolism in intact wheat seedlings under normal and stress conditions. J. Plant Physiol. 1984, 117, 201–209. [Google Scholar] [CrossRef]
- Sauter, G.; Simon, R. Perspective-predictive molecular pathology. N. Engl. J. Med. 2002, 347, 1995–1996. [Google Scholar] [CrossRef] [PubMed]
- Widhalm, J.R.; Dudareva, N. A familiar ring to it: Biosynthesis of plant benzoic acids. Mol. Plant. 2015, 8, 83–97. [Google Scholar] [CrossRef] [Green Version]
- Creelman, R.A.; Mullet, J.E. Jasmonic acid distribution and action in plants: Regulation during development and response to biotic and abiotic stress. Proc. Natl. Acad. Sci. USA 1995, 92, 4114–4119. [Google Scholar] [CrossRef] [Green Version]
- Bertini, L.; Palazzi, L.; Proietti, S.; Pollastri, S.; Arrigoni, G.; de Laureto, P.; Carusco, C. Proteomic analysis of MeJa-induced defense responses in rice against wounding. Int. J. Mol. Sci. 2019, 2020, 2525. [Google Scholar] [CrossRef] [Green Version]
- Giri, C.C.; Zaheer, M. Chemical elicitors versus secondary metabolite production in vitro using plant cell, tissue and organ cultures: Recent trends and a sky eye view appraisal. Plant Cell Tissue Organ Cult 2016, 126, 1–18. [Google Scholar] [CrossRef]
- Ho, T.-T.; Murthy, H.N.; Park, S.-Y. Methyl jasmonate induced oxidative stress and accumulation of secondary metabolites in plant cell and organ cultures. Int. J. Mol. Sci. 2020, 21, 716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Böttcher, C.; Burbidge, C.A.; di Rienzo, V.; Boss, P.K.; Davies, C. Jasmonic acid-isoleucine formation in grapevine (Vitis vinifera L.) by two enzymes with distinct transcription profiles. J. Integr. Plant Biol. 2015, 57, 618–627. [Google Scholar] [CrossRef]
- Gibb, M.; Kisiala, A.B.; Morrison, E.N.; Emery, R.J.N. The origins and roles of methylthiolated cytokinins: Evidence from among life kingdoms. Front. Cell Dev. Biol. 2020, 8, 605672. [Google Scholar] [CrossRef]
- Žižková, E.; Kubeš, M.; Dobrev, P.I.; Přibyl, P.; Šimura, J.; Zahajská, L.; Záveská Drábková, L.; Novák, O.; Motyka, V. Control of cytokinin and auxin homeostasis in cyanobacteria and algae. Ann. Bot. 2017, 119, 151–166. [Google Scholar] [CrossRef] [Green Version]
- Záveská Drábková, L.; Dobrev, P.I.; Motyka, V. Phytohormone profiling across the bryophytes. PLoS ONE 2015, 10, e0125411. [Google Scholar]
- Zemanová, V.; Pavlíková, D.; Dobrev, P.I.; Motyka, V.; Pavlík, M. Endogenous phytohormone profiles in Pteris fern species differing in arsenic accumulating ability. Environ. Exp. Bot. 2019, 166, 103822. [Google Scholar] [CrossRef]
- Gajdošová, S.; Spíchal, L.; Kamínek, M.; Hoyerová, K.; Novák, O.; Dobrev, P.I.; Galuszka, P.; Klíma, P.; Gaudinová, A.; Žižková, E.; et al. Distribution, biological activities, metabolism, and the conceivable function of cis-zeatin-type cytokinins in plants. J. Exp. Bot. 2011, 62, 2827–2840. [Google Scholar] [CrossRef] [Green Version]
- Sugawara, S.; Mashiguchi, K.; Tanaka, K.; Hishiyama, S.; Sakai, T.; Hanada, K.; Kinoshita-Tsujimura, K.; Yu, H.; Dai, X.; Takebayashi, Y.; et al. Distinct Characteristics of Indole-3-Acetic Acid and Phenylacetic Acid, Two Common Auxins in Plants. Plant Cell Physiol. 2015, 56, 1641–1654. [Google Scholar] [CrossRef] [Green Version]
- Sauer, M.; Robert, S.; Kleine-Vehn, J. Auxin: Simply complicated. J. Exp. Bot. 2013, 64, 2565–2577. [Google Scholar] [CrossRef] [Green Version]
- Bondev, I. Map of the Florostic Regions of Bulgaria. In Flora of the Peoples Republic of BulgariaI; Yordanov, D., Ed.; Publishing House of the Bulgarian Academy of Sciences: Sofia, Bulgaria, 1966; Volume 3, p. 638. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Gamborg, O.L.; Miller, R.A.; Ojima, K. Nutreint requerments of suspension culture of soybean root cells. Exp. Cell Res. 1968, 50, 151–158. [Google Scholar] [CrossRef]
- Danova, K.; Čellárová, E.; Macková, A.; Daxnerová, Z.; Kapchina-Toteva, V. In vitro culture of Hypericum rumeliacum Boiss. and production of phenolics and flavonoids. In Vitr. Cell. Dev. Biol.-Plant 2010, 46, 422–429. [Google Scholar] [CrossRef]
- Treneva, G.; Markovska, Y.; Wolfram, E.; Danova, K. Effect of plant growth regulators on growth patterns and enzymatic antioxidant activities in Hypericum calycinum shoot cultures. Bulg. J. Agric. Sci. 2014, 20 (Suppl. S1), 46–50. [Google Scholar]
- EDQM. European Pharmacopoeia, 8th ed.; Deutscher Apotheker Verlag: Stuttgart, Germany, 2013; Volume 7, p. 1438. [Google Scholar]
- Dobrev, P.I.; Kaminek, M. Fast and efficient separation of cytokinins from auxin and abscisic acid and their purification using mixed-mode solid-phase extraction. J. Chromatogr. A 2002, 950, 21–29. [Google Scholar] [CrossRef]
- Djilianov, D.L.; Dobrev, P.I.; Moyankova, D.P.; Vaňková, R.; Georgieva, D.T.; Gajdošová, S.; Motyka, V. Dynamics of endogenous phytohormones during dessication and recovery of the resurrection plant species Haberlea rhodopensis. J. Plant Growth Regul. 2013, 32, 564–574. [Google Scholar] [CrossRef]
- Žižková, E.; Dobrev, P.I.; Muhovski, Y.; Hošek, P.; Hoyerová, K.; Haisel, D.; Procházková, D.; Lutts, S.; Motyka, V.; Hichri, I. Tomato (Solanum lycopersicum L.) SlIPT3 and SlIPT4 isopentenyltransferases mediate salt stress response in tomato. BMC Plant Biol. 2015, 15, 85. [Google Scholar] [CrossRef]
- Kamínek, M.; Březinová, A.; Gaudinová, A.; Motyka, V.; Vaňková, R.; Zažímalová, E. Purine cytokinins: A proposal for abbreviations. Plant Growth Regul. 2000, 32, 253–256. [Google Scholar] [CrossRef]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Danova, K.; Motyka, V.; Trendafilova, A.; Dobrev, P.I.; Ivanova, V.; Aneva, I. Evolutionary Aspects of Hypericin Productivity and Endogenous Phytohormone Pools Evidenced in Hypericum Species In Vitro Culture Model. Plants 2022, 11, 2753. https://doi.org/10.3390/plants11202753
Danova K, Motyka V, Trendafilova A, Dobrev PI, Ivanova V, Aneva I. Evolutionary Aspects of Hypericin Productivity and Endogenous Phytohormone Pools Evidenced in Hypericum Species In Vitro Culture Model. Plants. 2022; 11(20):2753. https://doi.org/10.3390/plants11202753
Chicago/Turabian StyleDanova, Kalina, Vaclav Motyka, Antoaneta Trendafilova, Petre I. Dobrev, Viktorya Ivanova, and Ina Aneva. 2022. "Evolutionary Aspects of Hypericin Productivity and Endogenous Phytohormone Pools Evidenced in Hypericum Species In Vitro Culture Model" Plants 11, no. 20: 2753. https://doi.org/10.3390/plants11202753
APA StyleDanova, K., Motyka, V., Trendafilova, A., Dobrev, P. I., Ivanova, V., & Aneva, I. (2022). Evolutionary Aspects of Hypericin Productivity and Endogenous Phytohormone Pools Evidenced in Hypericum Species In Vitro Culture Model. Plants, 11(20), 2753. https://doi.org/10.3390/plants11202753