


A New Biocontrol Tool to Fight Potato Late Blight Based on Willaertia magna C2c Maky Lysate



Abstract
:1. Introduction
2. Results
2.1. Elicitor Effect on Potato Genes
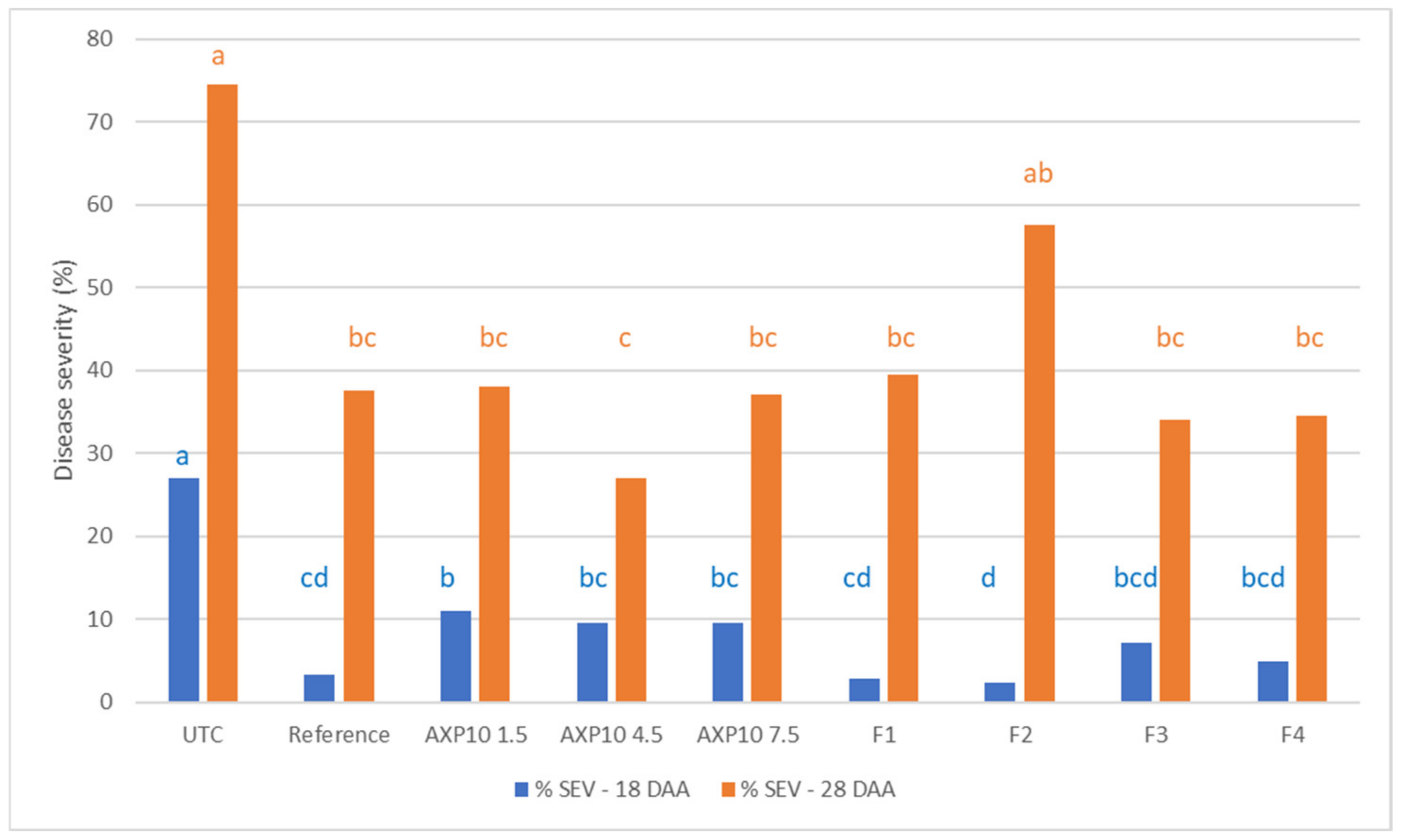
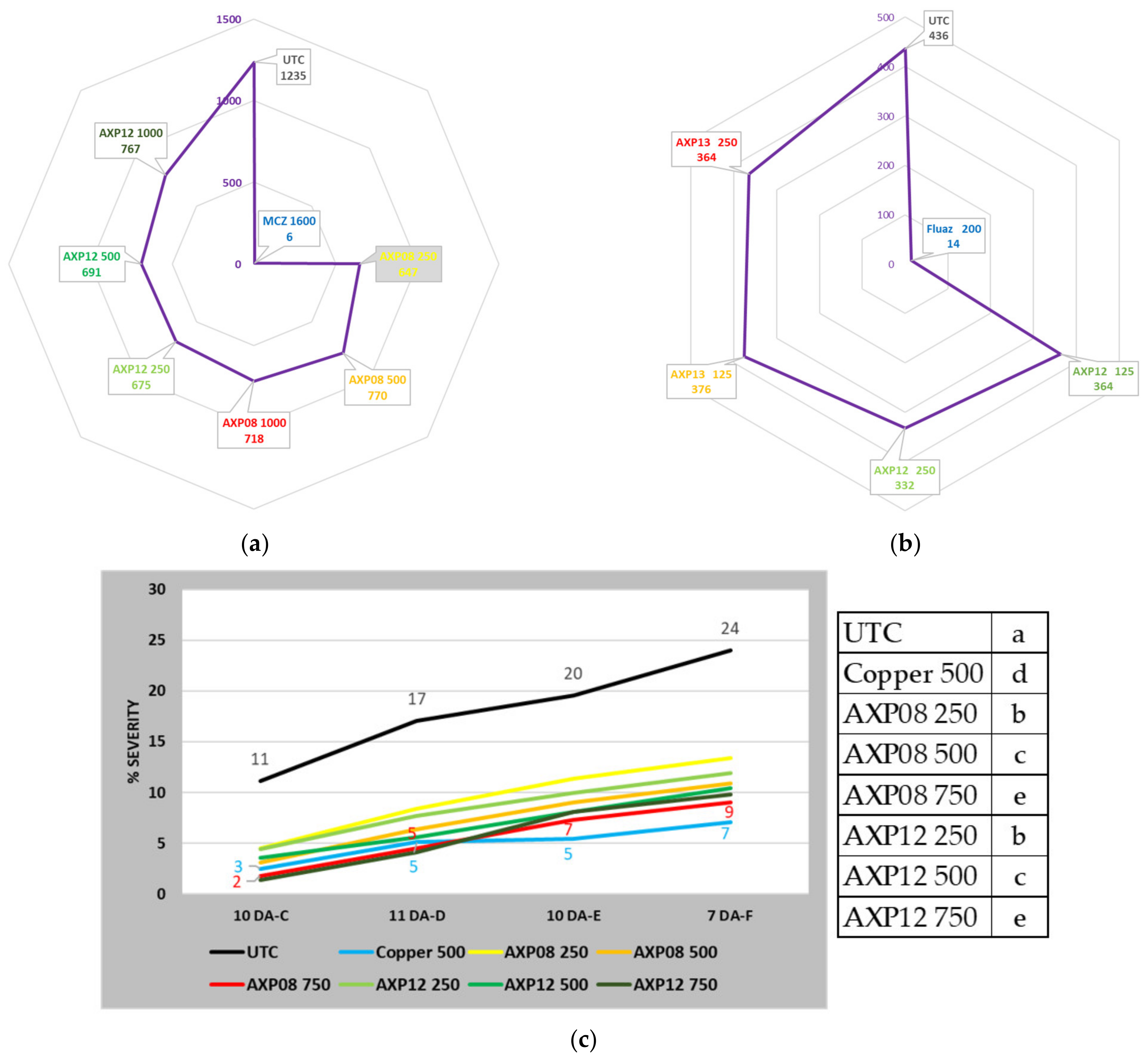
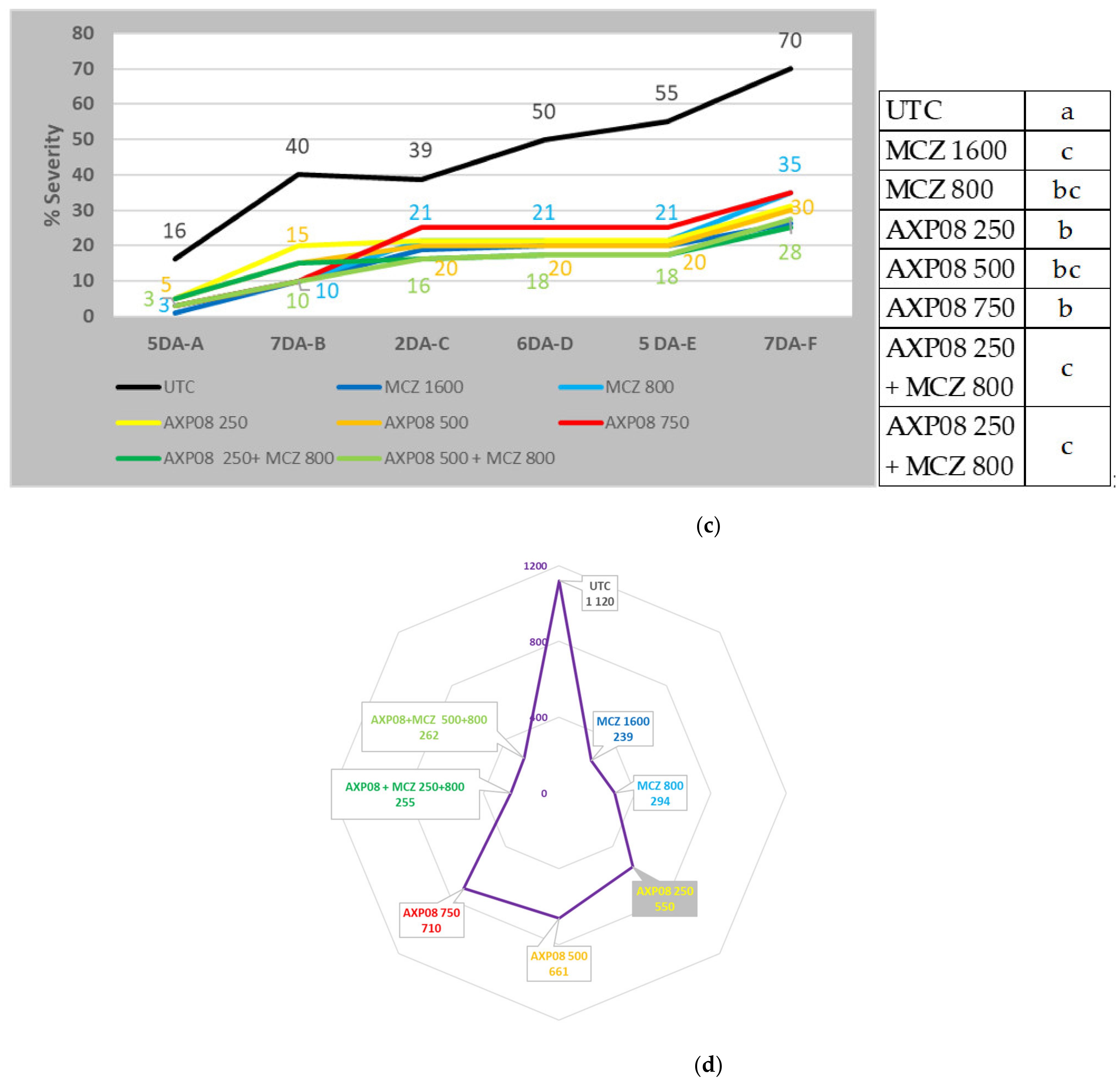
2.2. Greenhouse Efficacy Tests
2.2.1. 2019 Greenhouse Trial
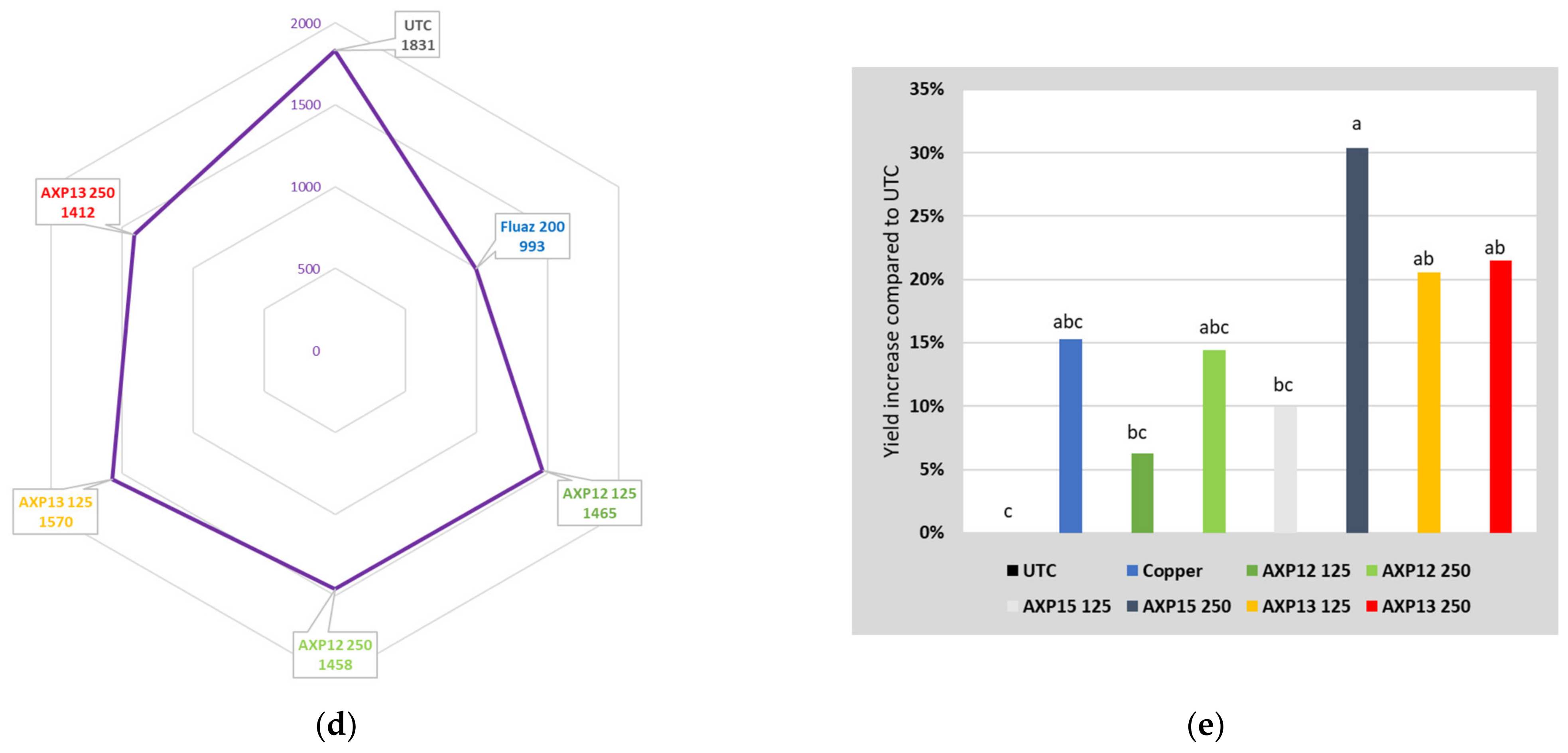
2.2.2. 2021 Greenhouse Trial
2.3. Field Trials
3. Discussion
4. Materials and Methods
4.1. Biocontrol Products
4.2. Anti-Oomycetal Tests under Greenhouse Conditions
4.2.1. 2019 Greenhouse Trial
4.2.2. 2021 Greenhouse Trial
4.3. Plant Defense Gene Study by qPFD®
4.3.1. Biological Material
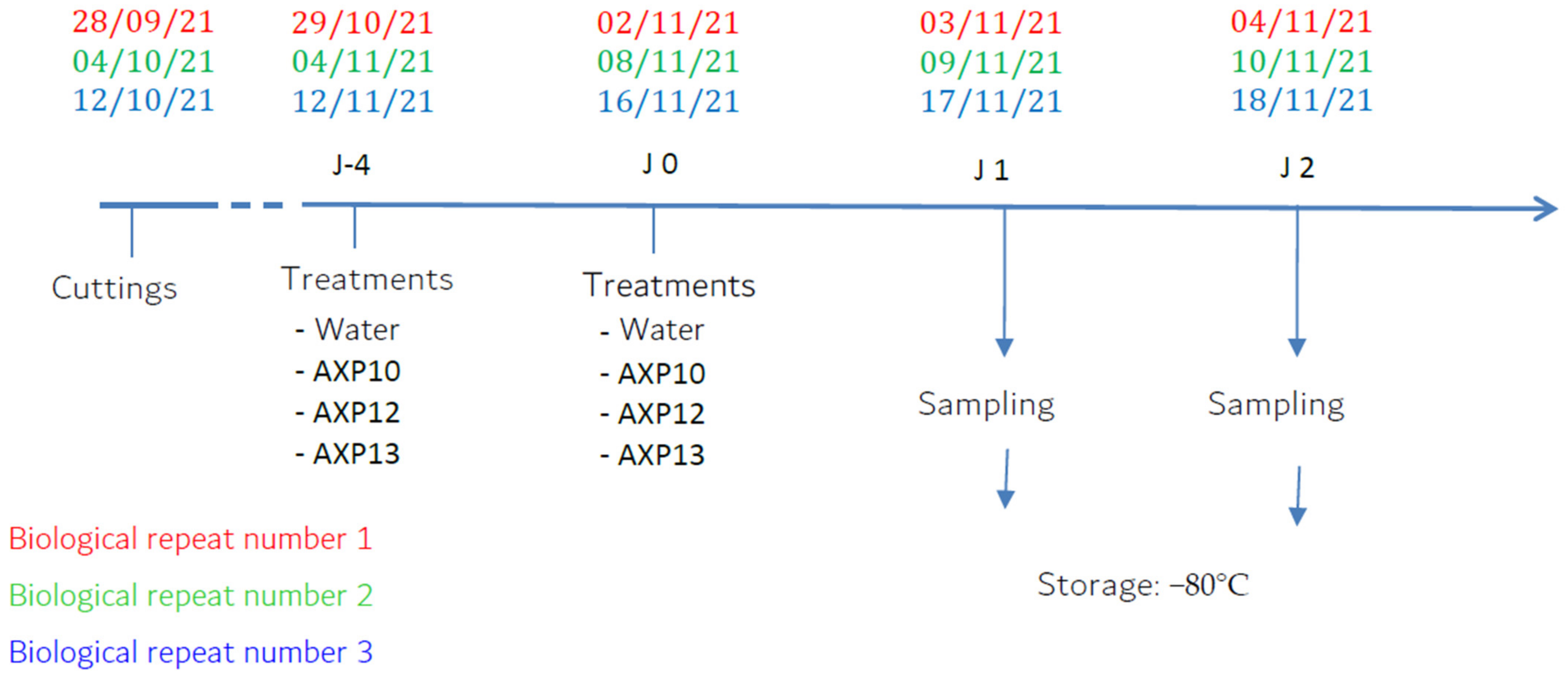
4.3.2. Plant Treatment and Samplings
- -
- Water (negative control);
- -
- AXP10: AS not formulated (1 g/L);
- -
- AXP12: formulation (1 or 5 g/L); and
- -
- AXP13: formulation (1 or 5 g/L).
4.3.3. Gene Expression Monitoring
4.4. Field Trials
4.5. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 13 July 2022).
- Potatoes 2020 Summary 28 September 2021. Available online: https://usda.library.cornell.edu/concern/publications/fx719m44h (accessed on 29 July 2022).
- Wild Potatoes Tapped for Late Blight Guard Duty: USDA ARS. Available online: https://www.ars.usda.gov/news-events/news/research-news/2021/wild-potatoes-tapped-for-late-blight-guard-duty/ (accessed on 11 April 2022).
- Fry, W. Phytophthora Infestans: The Plant (and R Gene) Destroyer. Mol. Plant Pathol. 2008, 9, 385–402. [Google Scholar] [CrossRef] [PubMed]
- Bødker, L.; Pedersen, H.; Kristensen, K.; Møller, L.; Lehtinen, A.; Hannukkala, A. Influence of Crop History of Potato on Early Occurrence and Disease Severity of Potato Late Blight Caused by Phytophthora Infestans. PPO Spec. Rep. 2006, 11, 53. [Google Scholar]
- Schepers, H.T.A.M.; Kessel, G.J.T.; Lucca, F.; Förch, M.G.; van den Bosch, G.B.M.; Topper, C.G.; Evenhuis, A. Reduced Efficacy of Fluazinam against Phytophthora Infestans in the Netherlands. Eur. J. Plant Pathol. 2018, 151, 947–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, B.J.; Kamoun, S.; Zody, M.C.; Jiang, R.H.Y.; Handsaker, R.E.; Cano, L.M.; Grabherr, M.; Kodira, C.D.; Raffaele, S.; Torto-Alalibo, T.; et al. Genome Sequence and Analysis of the Irish Potato Famine Pathogen Phytophthora Infestans. Nature 2009, 461, 393–398. [Google Scholar] [CrossRef] [Green Version]
- A European Green Deal. Available online: https://ec.europa.eu/info/strategy/priorities-2019-2024/european-green-deal_en (accessed on 13 July 2022).
- Vilvert, E.; Stridh, L.; Andersson, B.; Olson, Å.; Aldén, L.; Berlin, A. Evidence Based Disease Control Methods in Potato Production: A Systematic Map Protocol. Environ. Evid. 2022, 11, 6. [Google Scholar] [CrossRef]
- Commission Implementing Regulation (EU). 2020/2087 of 14 December 2020 Concerning the Non-Renewal of the Approval of the Active Substance Mancozeb, in Accordance with Regulation (EC) No 1107/2009 of the European Parliament and of the Council Concerning the Placing of Plant Protection Products on the Market, and Amending the Annex to Commission Implementing Regulation (EU) No 540/2011 (Text with EEA Relevance). Available online: http://data.europa.eu/eli/reg_impl/2020/2087/oj/eng (accessed on 24 August 2022).
- Demanèche, S.; Mirabel, L.; Abbe, O.; Eberst, J.-B.; Souche, J.-L. A New Active Substance Derived from Lyzed Willaertia Magna C2c Maky Cells to Fight Grapevine Downy Mildew. Plants 2020, 9, 1013. [Google Scholar] [CrossRef]
- Brisset, M.-N.; De Bernonville, T.D. Device for Determining or Studying the State of Stimulation of the Natural Defences of Plants or Portions of Plants 2016. U.S. Patent No 9,290,788, 22 March 2016. [Google Scholar]
- Leesutthiphonchai, W.; Vu, A.L.; Ah-Fong, A.M.V.; Judelson, H.S. How Does Phytophthora Infestans Evade Control Efforts? Modern Insight Into the Late Blight Disease. Phytopathology 2018, 108, 916–924. [Google Scholar] [CrossRef] [Green Version]
- Bangemann, L.-W.; Westphal, A.; Zwerger, P.; Sieling, K.; Kage, H. Copper Reducing Strategies for Late Blight (Phytophthora infestons) Control in Organic Potato (Solanum tuberosum) Production. J. Plant Dis. Prot. 2014, 121, 105–116. [Google Scholar] [CrossRef]
- Aamir, M.; Singh, V.K.; Dubey, M.K.; Kashyap, S.P.; Zehra, A.; Upadhyay, R.S.; Singh, S. Structural and Functional Dissection of Differentially Expressed Tomato WRKY Transcripts in Host Defense Response against the Vascular Wilt Pathogen (Fusarium oxysporum f. sp. Lycopersici). PLoS ONE 2018, 13, e0193922. [Google Scholar] [CrossRef] [Green Version]
- Eulgem, T.; Somssich, I.E. Networks of WRKY Transcription Factors in Defense Signaling. Curr. Opin. Plant Biol. 2007, 10, 366–371. [Google Scholar] [CrossRef] [Green Version]
- Almagro, L.; Gómez Ros, L.V.; Belchi-Navarro, S.; Bru, R.; Ros Barceló, A.; Pedreño, M.A. Class III Peroxidases in Plant Defence Reactions. J. Exp. Bot. 2009, 60, 377–390. [Google Scholar] [CrossRef] [PubMed]
- Dao, T.T.H.; Linthorst, H.J.M.; Verpoorte, R. Chalcone Synthase and Its Functions in Plant Resistance. Phytochem. Rev. 2011, 10, 397–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinderer, W.; Seitz, H.U. Chalcone Synthase from Cell Suspension Cultures of Daucus carota L. Arch. Biochem. Biophys. 1985, 240, 265–272. [Google Scholar] [CrossRef]
- Hess, F.D.; Foy, C.L. Interaction of Surfactants with Plant Cuticles. Weed Technol. 2000, 14, 807–813. [Google Scholar] [CrossRef]
- El-Hasan, A.; Ngatia, G.; Link, T.I.; Voegele, R.T. Isolation, Identification, and Biocontrol Potential of Root Fungal Endophytes Associated with Solanaceous Plants against Potato Late Blight (Phytophthora infestans). Plants 2022, 11, 1605. [Google Scholar] [CrossRef] [PubMed]
- Léger, G.; Novinscak, A.; Biessy, A.; Lamarre, S.; Filion, M. In Tuber Biocontrol of Potato Late Blight by a Collection of Phenazine-1-Carboxylic Acid-Producing Pseudomonas spp. Microorganisms 2021, 9, 2525. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.H.; Ma, Q.; Sun, Z.N.; Cui, H.C.; Liu, H.R. Biocontrol Mechanism of Myxococcus Fulvus B25-I-3 against Phytophthora Infestans and Its Control Efficiency on Potato Late Blight. Folia Microbiol. (Praha) 2021, 66, 555–567. [Google Scholar] [CrossRef]
- Wu, Z.; Cui, H.; Sun, Z.; Liu, H. Biocontrol Mechanism of Myxococcus Xanthus B25-I-1 against Phytophthora Infestans. Pestic. Biochem. Physiol. 2021, 175, 104832. [Google Scholar] [CrossRef]
- Linkies, A.; Jacob, S.; Zink, P.; Maschemer, M.; Maier, W.; Koch, E. Characterization of Cultural Traits and Fungicidal Activity of Strains Belonging to the Fungal Genus Chaetomium. J. Appl. Microbiol. 2021, 131, 375–391. [Google Scholar] [CrossRef]
- Feng, S.; Jian, Y.; Jin, L.; Tang, S.; Li, Z. Complete Genome Sequence Data of Rare Actinomycetes Strain Saccharothrix Texasensis 6-C, a Biological Control Agent for Potato Late Blight. Mol. Plant Microbe Interact. 2021, 34, 571–574. [Google Scholar] [CrossRef]
- Feng, S.; Jin, L.; Tang, S.; Jian, Y.; Li, Z. Combination of Rhizosphere Bacteria Isolated from Resistant Potato Plants for Biocontrol of Potato Late Blight. Pest Manag. Sci. 2022, 78, 166–176. [Google Scholar] [CrossRef] [PubMed]
- Ghadamgahi, F.; Tarighi, S.; Taheri, P.; Saripella, G.V.; Anzalone, A.; Kalyandurg, P.B.; Catara, V.; Ortiz, R.; Vetukuri, R.R. Plant Growth-Promoting Activity of Pseudomonas Aeruginosa FG106 and Its Ability to Act as a Biocontrol Agent against Potato, Tomato and Taro Pathogens. Biology 2022, 11, 140. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Shim, C.K.; Park, J.-H. Control Efficacy of Bacillus Velezensis AFB2-2 against Potato Late Blight Caused by Phytophthora Infestans in Organic Potato Cultivation. Plant Pathol. J. 2021, 37, 580–595. [Google Scholar] [CrossRef]
- Modesto, O.O.; Anwar, M.; He, Z.; Larkin, R.P.; Honeycutt, C.W. Survival Potential of Phytophthora Infestans Sporangia in Relation to Environmental Factors and Late Blight Occurrence. J. Plant Prot. Res. 2016, 56, 73–81. [Google Scholar] [CrossRef]
- Hadders, J. An Example of Integrated Forecasting System for Phytophthora Infestans on Potato. In Integrated Management of Diseases Caused by Fungi, Phytoplasma and Bacteria; Ciancio, A., Mukerji, K.G., Eds.; Integrated Management of Plant Pests and Diseases; Springer: Dordrecht, The Netherlands, 2008; pp. 179–189. ISBN 978-1-4020-8571-0. [Google Scholar]
- Wander, J.G.N.; Spits, H.G.; Kessel, G.J.T. Exploiting Potato Late Blight Cultivar Resistance Using DSS’s: 4 Years of Field Experiments. PPO-Special Report. 2006. Available online: https://edepot.wur.nl/19021 (accessed on 15 July 2022).
- Pandit, M.A.; Kumar, J.; Gulati, S.; Bhandari, N.; Mehta, P.; Katyal, R.; Rawat, C.D.; Mishra, V.; Kaur, J. Major Biological Control Strategies for Plant Pathogens. Pathogens 2022, 11, 273. [Google Scholar] [CrossRef] [PubMed]
- Asimakis, E.; Shehata, A.A.; Eisenreich, W.; Acheuk, F.; Lasram, S.; Basiouni, S.; Emekci, M.; Ntougias, S.; Taner, G.; May-Simera, H.; et al. Algae and Their Metabolites as Potential Bio-Pesticides. Microorganisms 2022, 10, 307. [Google Scholar] [CrossRef] [PubMed]
- Dugé de Bernonville, T.; Marolleau, B.; Staub, J.; Gaucher, M.; Brisset, M.-N. Using Molecular Tools To Decipher the Complex World of Plant Resistance Inducers: An Apple Case Study. J. Agric. Food Chem. 2014, 62, 11403–11411. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment kg/ha | Spray Time | 4 dpi | 5 dpi | 7 dpi |
|---|---|---|---|---|
| AXP12 0.625 | T-1 | 10.9 ijk | 28.5 jk | 51.1 hijk |
| T-3 | 14.6 k | 28.2 ijk | 54.6 k | |
| T-7 | 11.8 ijk | 30.3 k | 41.0 efghijk | |
| AXP12 1.25 | T-1 | 10.8 ijk | 28.5 jk | 40.8 efghijk |
| T-3&T-1 | 6.1 efg | 19.0 efgh | 38.1 defghi | |
| T-3 | 14.1 jk | 26.3 hijk | 53.0 jk | |
| T-7 | 13.3 jk | 28.5 jk | 49.0 fghijk | |
| AXP12 2.5 | T-1 | 4.5 def | 17.2 def | 35.9 def |
| T-3 | 8.9 ghij | 23.4 fghijk | 39.2 defghij | |
| T-7 | 16.9 k | 29.8 k | 49.9 ghijk | |
| AXP13 0.625 | T-1 | 4.5 def | 20.6 fghij | 37.1 defgh |
| T-3 | 6.2 efg | 22.8 fghijk | 35.9 def | |
| T-7 | 10.7 hijk | 28.7 jk | 51.9 ijk | |
| AXP13 1.25 | T-1 | 3.7 de | 14.4 de | 29.5 d |
| T-3&T-1 | 2.7 cd | 12.7 d | 28.7 d | |
| T-3 | 6.5 fgh | 20.1 efghi | 36.8 defg | |
| T-7 | 10.9 ijk | 27.4 ijk | 44.1 efijk | |
| AXP13 2.5 | T-1 | 2.7 cd | 8.6 c | 19.7 c |
| T-3 | 5.2 ef | 17.4 defg | 33.0 de | |
| T-7 | 7.5 fghi | 24.4 ghijk | 33.5 de | |
| Cu 1.3 | T-1 | 0.9 ab | 3.4 b | 8.6 b |
| T-3&T-1 | 0.2 a | 1.2 a | 3.2 a | |
| T-3 | 0.9 ab | 2.8 b | 9.6 b | |
| T-7 | 1.5 bc | 4.0 b | 10.7 b | |
| UTC | T-7 | 13.3 jk | 31.2 k | 49.8 ghijk |
| F pr. | <0.001 | <0.001 | <0.001 | |
| Trial Reference | Severity in UTC * | Product | Yield | AUDPC | Dose (gai/ha) | Best Product Efficacy/Severity in UTC |
|---|---|---|---|---|---|---|
| 2020PTMDF | 100% | None | N | N | 0%/60% | |
| 20A6-02FP-1 | 100% | None | N | N | 0%/100% | |
| RFR20-133-518FE | 92% | None | N | N | 0%/92% | |
| FR21F07 | 59% | AXP13 | N | N | 250 | 48%/29% |
| 20-42491-FR01 | 78% | AXP08 | N | Y | 250 | 48%/78% |
| F536421 | 66% | AXP12 | N | N | 250 | 58%/38% |
| 21-48677-FR02 | 86% | AXP12 | N | Y | 250 | 24%/86% |
| 225.F.SAG20 | 24% | AXP08 | N | N | 750 | 63%/24% |
| 21-48677-BE03 | 95% | AXP13 | N | Y | 250 | 52%/79% |
| CT21-4-30DE1 | 61% | AXP15 | Y | N | 250 | 57%/35% |
| 2021PTMDF01 | 100% | AXP15 | Y | Y | 250 | 49%/100% |
| AmoebaLB2020 | 60% | AXP12 | Y | Y | 500 | 56%/40% |
| AmoebaRR 2020 | 28% | AXP08 | Y | Y | 250 | 77%/28% |
| CT20-4-95DE1 | 70% | AXP08 | Y | Y | 500 | 59%/70% |
| 20-42491-BE03 | 84% | AXP08 | N | Y | 250 | 58%/30% |
| Fungicide | Active Ingredient | Dose Rate (L or kg/ha) | Day | Before | P. infestans | Inoculation |
|---|---|---|---|---|---|---|
| T-7 | T-3 | T-1 | T-3&T-1 | |||
| UTC | - | - | A * | - | - | - |
| Cuprozin | Cu 383 g/L | 1.3 | B | I | T | K |
| AXP12 | 200 g/kg | 0.625 | C | L | U | - |
| AXP12 | 200 g/kg | 1.25 | D | M | W | N |
| AXP12 | 200 g/kg | 2.5 | E | O | X | - |
| AXP13 | 217 g/kg | 0.625 | F | P | Y | - |
| AXP13 | 217 g/kg | 1.25 | G | Q | Z | R |
| AXP13 | 217 g/kg | 2.5 | H | S | AA | - |
| Defense Classes and Subclasses | Defense Genes | ||
|---|---|---|---|
| Gene Code | Complete Name | ||
| Chemical and/or physical barriers | PR proteins | PR-1 | Pathogenesis-related protein 1 |
| PR-2 | Pathogenesis-related protein 2 (glucanases) | ||
| PR-4 | Pathogenesis-related protein 4 (hevein-like) | ||
| PR-5 | Pathogenesis-related protein 5 (thaumatin-like, osmotin) | ||
| PR-8 | Pathogenesis-related protein 8 (class III chitinase) | ||
| PR-14 | Pathogenesis-related protein 14 (lipid transfer protein) | ||
| PR-15 | Pathogenesis-related protein 15 (oxalate oxidase) | ||
| Phenylpropanoids | PAL | Phenylalanine ammonia-lyase | |
| CHS | Chalcone synthase | ||
| DFR | Dihydroflavonol reductase | ||
| ANS | Anthocyanidin synthase | ||
| PPO | Polyphenol oxidase | ||
| Isoprenoids | HMGR | Hydroxymethyl glutarate-CoA reductase | |
| FPPS | Farnesyl pyrophosphate synthase | ||
| Far | (E,E)-alpha-farnesene synthase | ||
| Cysteines | CSL | Alliinase | |
| Oxidative stress | APOX | Ascorbate peroxidase | |
| GST | Glutathion S-transférase | ||
| POX | Peroxidase | ||
| Parietal modification | CalS | Callose synthase | |
| Pect | Pectin methyl esterase | ||
| CAD | Cinnamyl alcohol dehydrogenase | ||
| Hormonal signaling | Salicylic acid (SA) | EDS1 | Disease resistance protein EDS 1 |
| WRKY | WRKY transcription factor 30 | ||
| Jasmonic acid (JA) | LOX2 | Lipoxygenase AtLOX2 | |
| JAR | Jasmonate resistant 1 | ||
| Ethylene (ET) | ACCO | 1-aminocyclopropene-1-carboxylate oxidase | |
| EIN3 | EIN3-BINDING F BOX PROTEIN 1 | ||
| 2020PTMDF | Product | Form | CONC. | rate (kg/ha) | AS | Rate (g/ha) |
|---|---|---|---|---|---|---|
| 1 | UTC | |||||
| 2 | DITHANE | WP | 80% | 2.00 | mancozeb | 1600 |
| 3 | AXP08 | WP | 60.5% | 0.42 | amoeba lysate | 250 |
| 4 | AXP08 | WP | 60.5% | 0.84 | amoeba lysate | 500 |
| 5 | AXP08 | WP | 60.5% | 1.25 | amoeba lysate | 750 |
| 6 | AXP12 | SC | 20% | 1.25 | amoeba lysate | 250 |
| 7 | AXP12 | SC | 20% | 2.50 | amoeba lysate | 500 |
| 8 | AXP12 | SC | 20% | 3.75 | amoeba lysate | 750 |
| 2020PTMDA | Product | Form | CONC. | Rate (kg/ha) | AS | Rate (g/ha) |
| 1 | UTC | |||||
| 2 | DITHANE | WP | 80% | 2.00 | mancozeb | 1600 |
| 3 | DITHANE | WP | 80% | 1.00 | amoeba lysate | 800 |
| 4 | AXP08 | WP | 60.5% | 0.42 | amoeba lysate | 250 |
| 5 | AXP08 | WP | 60.5% | 0.84 | amoeba lysate | 500 |
| 6 | AXP08 | WP | 60.5% | 1.25 | amoeba lysate | 750 |
| 7 | AXP08 + DITHANE | WP | 0.42 + 1 | amoeba lysate | 250 + 800 | |
| 8 | AXP08 + DITHANE | WP | 0.84 + 1 | amoeba lysate | 500 + 800 |
| 2021PTMDF1 | Product | Form | CONC. | Rate (kg/ha) | AS | Rate (gai/ha) |
|---|---|---|---|---|---|---|
| 1 | UTC | |||||
| 2 | Standard | SC | 50/25% | 0.40/2.5 | Fluazinam/Copper | 200/500 |
| 3 | AXP12 | SC | 20% | 0.63 | amoeba lysate | 125 |
| 4 | AXP12 | SC | 20% | 1.25 | amoeba lysate | 250 |
| 5 | AXP13 | OD | 21.7% | 0.63 | amoeba lysate | 125 |
| 6 | AXP13 | OD | 21.7% | 1.25 | amoeba lysate | 250 |
| 2021PTMDF2 | Product | Form | CONC. | Rate (kg/ha) | AS | Rate (gai/ha) |
| 1 | UTC | |||||
| 2 | Standard | SC | 25% | 2.5 | Copper | 500 |
| 3 | AXP12 | SC | 20% | 0.63 | amoeba lysate | 125 |
| 4 | AXP12 | SC | 20% | 1.25 | amoeba lysate | 250 |
| 5 | AXP15 | SC | 21.2% | 0.63 | amoeba lysate | 125 |
| 6 | AXP15 | SC | 21.2% | 1.25 | amoeba lysate | 250 |
| 7 | AXP13 | OD | 21.7% | 1.25 | amoeba lysate | 250 |
| 8 | AXP13 | OD | 21.7% | 1.25 | amoeba lysate | 250 |
| Year | Trial Reference | Contractor | Country | Variety | Applications |
|---|---|---|---|---|---|
| 2020 | AmoebaLB2020 | Hubbard | USA | Umatilla | 7T–7D |
| 2020 | AmoebaRR 2020 | Hubbard | USA | Ranger Russet | 7T–7D |
| 2020 | CT20-4-95DE1 | Crop Trials | Germany | Afra | 12T–5/7D |
| 2020 | 2020PTMDF | Hetterich | Germany | Bintje | 6T–7D |
| 2020 | 225.F.SAG20 | Sagea | Italy | Laura | 6T–10/11D |
| 2020 | 20-42491-FR01 | Staphyt | France | Caesar | 12T–7D |
| 2020 | 20A6-02FP-1 | SciencesAgro Atlantique | France | Bintje | 6T–7D |
| 2020 | RFR20-133-518FE | Syntech | France | Bintje | 12T–6/8D |
| 2020 | 20-42491-BE03 | Staphyt | Belgium | Challenger | 8T–7D |
| 2021 | F536421 | CentreExpé | France | El Mundo | 10T–4/12D |
| 2021 | FR21F07 | Ephydia | France | Bintje | 6T–4/8D |
| 2021 | 21-48677-FR02 | Staphyt | France | Désirée | 8T–3/7D |
| 2021 | 21-48677-BE03 | Staphyt | Belgium | Challenger | 10T–4/6D |
| 2021 | CT21-4-30DE1 | Crop Trials | Germany | Eurogrande | 10T–7D |
| 2021 | 2021PTMDF01 | Hetterich | Germany | Euroflora | 10T–6/8D |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Troussieux, S.; Gilgen, A.; Souche, J.-L. A New Biocontrol Tool to Fight Potato Late Blight Based on Willaertia magna C2c Maky Lysate. Plants 2022, 11, 2756. https://doi.org/10.3390/plants11202756
Troussieux S, Gilgen A, Souche J-L. A New Biocontrol Tool to Fight Potato Late Blight Based on Willaertia magna C2c Maky Lysate. Plants. 2022; 11(20):2756. https://doi.org/10.3390/plants11202756
Chicago/Turabian StyleTroussieux, Sandrine, Annabelle Gilgen, and Jean-Luc Souche. 2022. "A New Biocontrol Tool to Fight Potato Late Blight Based on Willaertia magna C2c Maky Lysate" Plants 11, no. 20: 2756. https://doi.org/10.3390/plants11202756
APA StyleTroussieux, S., Gilgen, A., & Souche, J. -L. (2022). A New Biocontrol Tool to Fight Potato Late Blight Based on Willaertia magna C2c Maky Lysate. Plants, 11(20), 2756. https://doi.org/10.3390/plants11202756