
Protein S-Acyl Transferase GhPAT27 Was Associated with Verticillium wilt Resistance in Cotton

Abstract
:1. Introduction
2. Results
2.1. Identification Information for Members of GhPATs
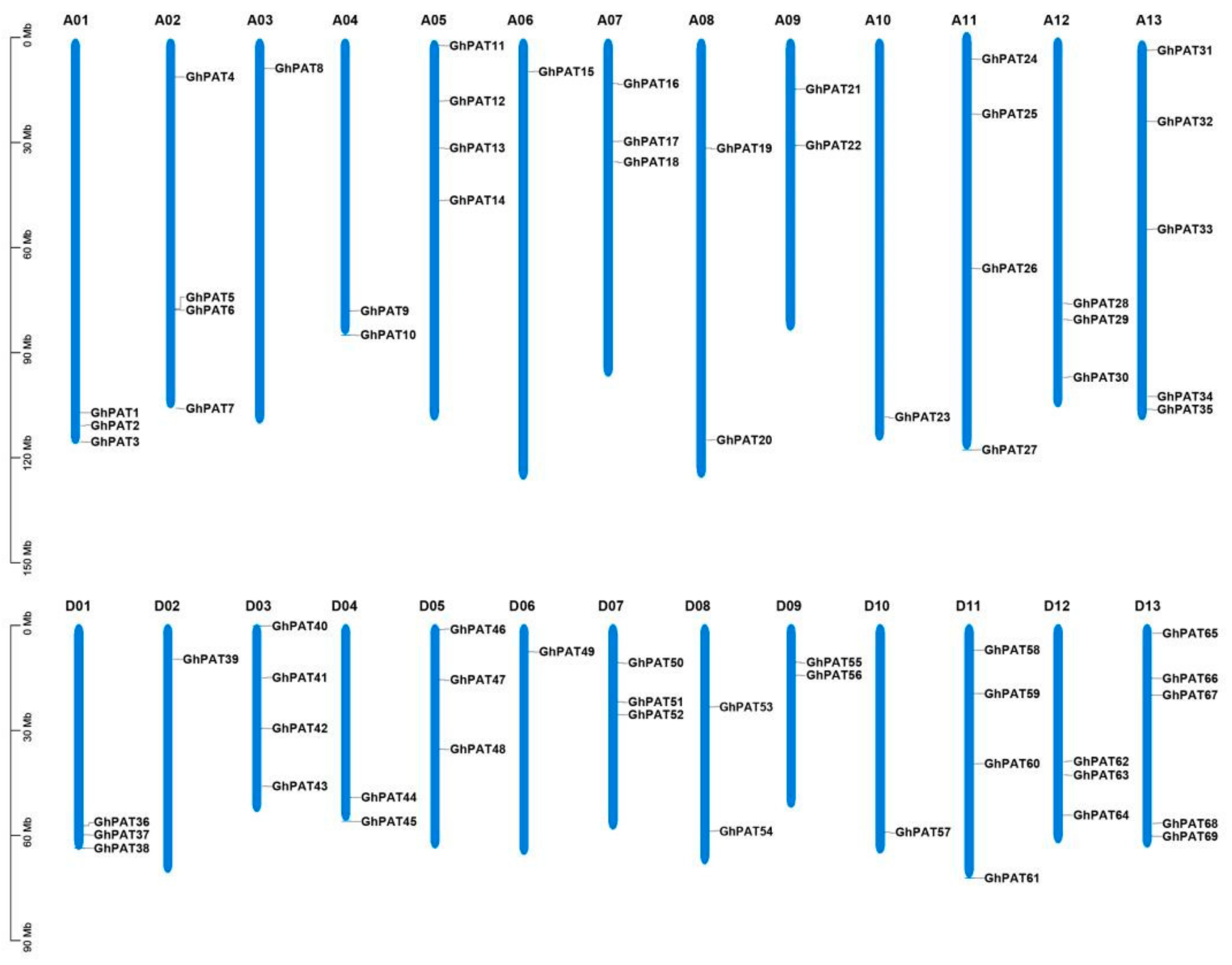
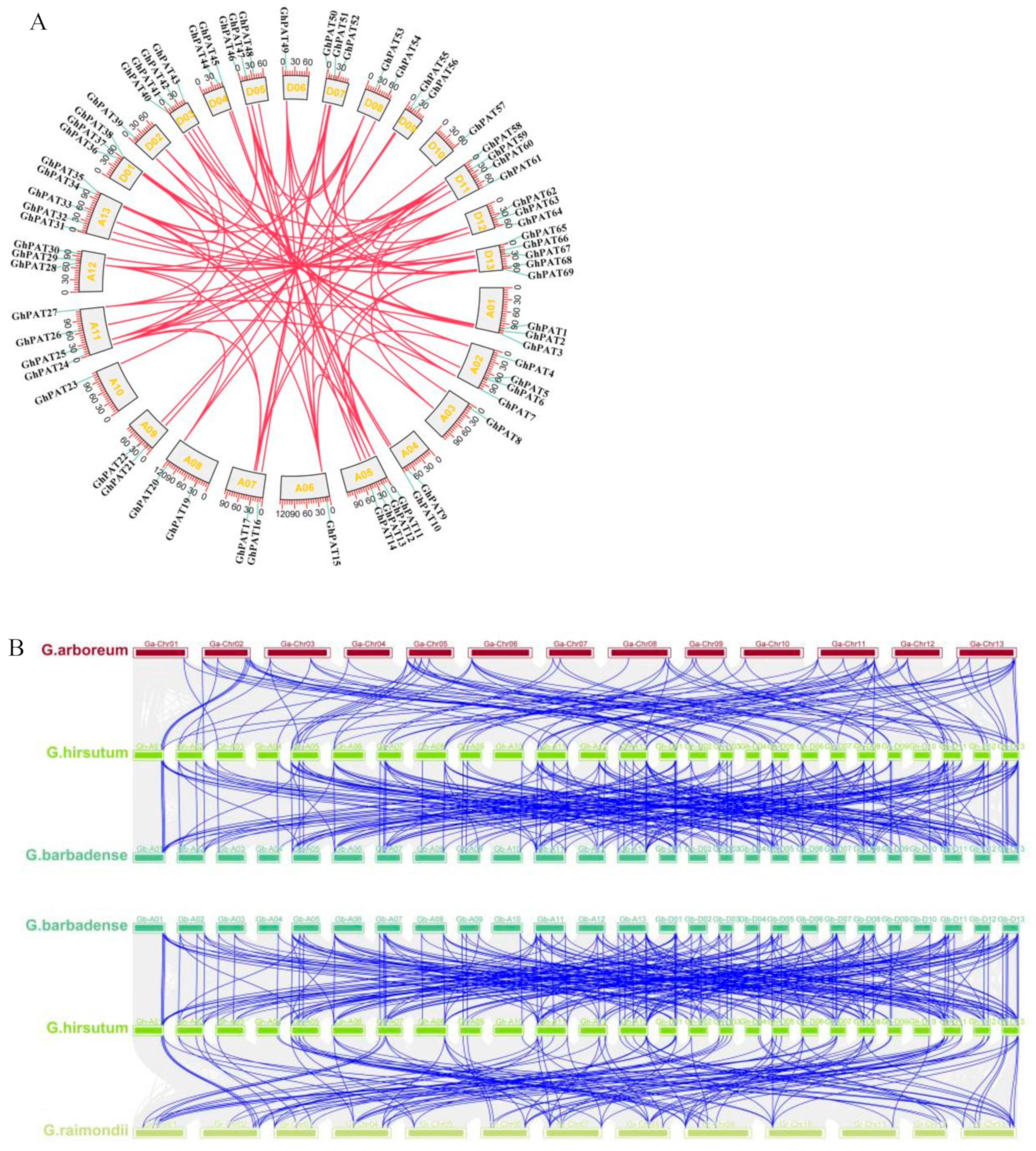
2.2. Chromosomal Distribution of GhPATs
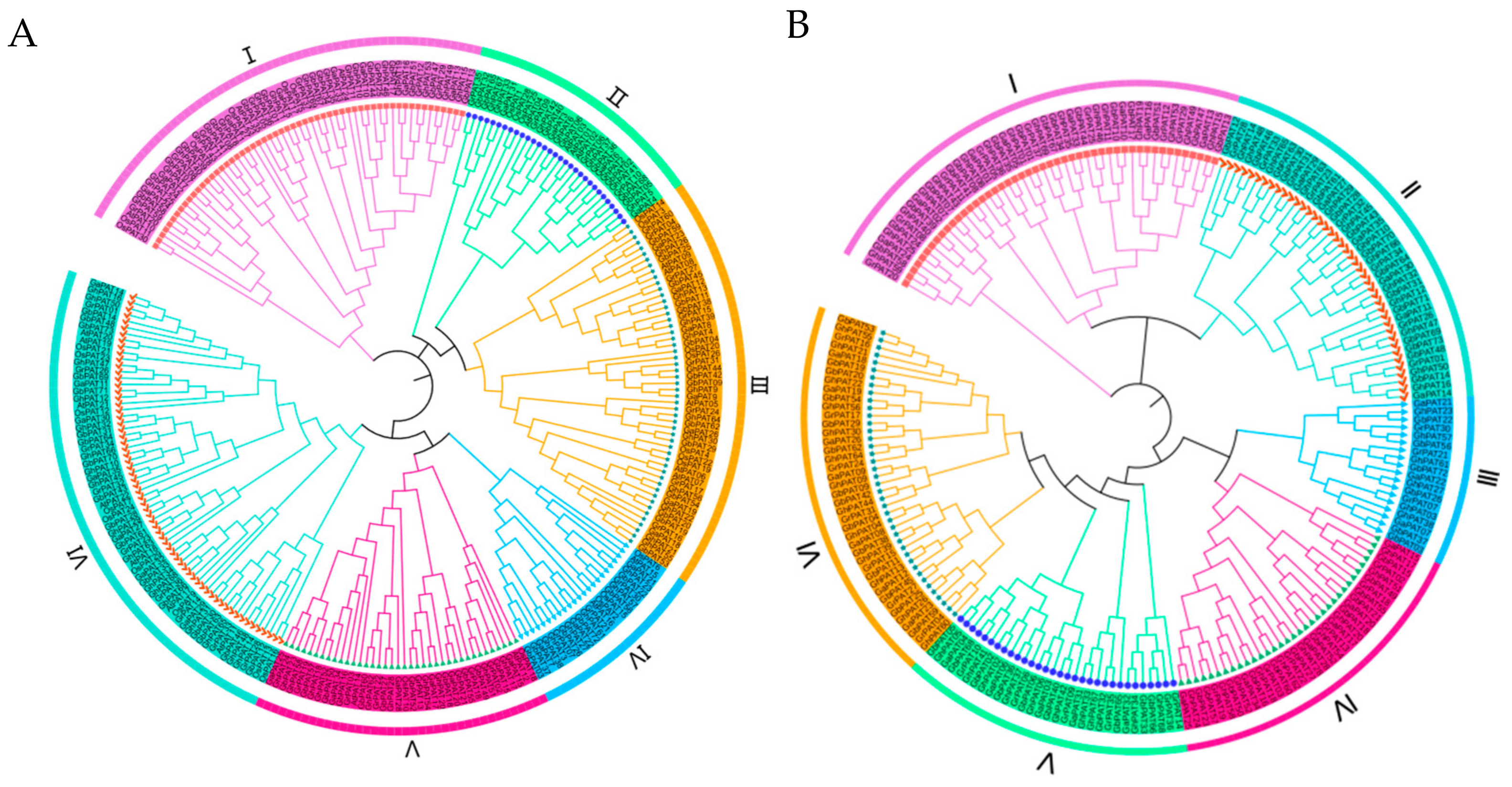
2.3. Phylogenetic Evaluation of GhPATs
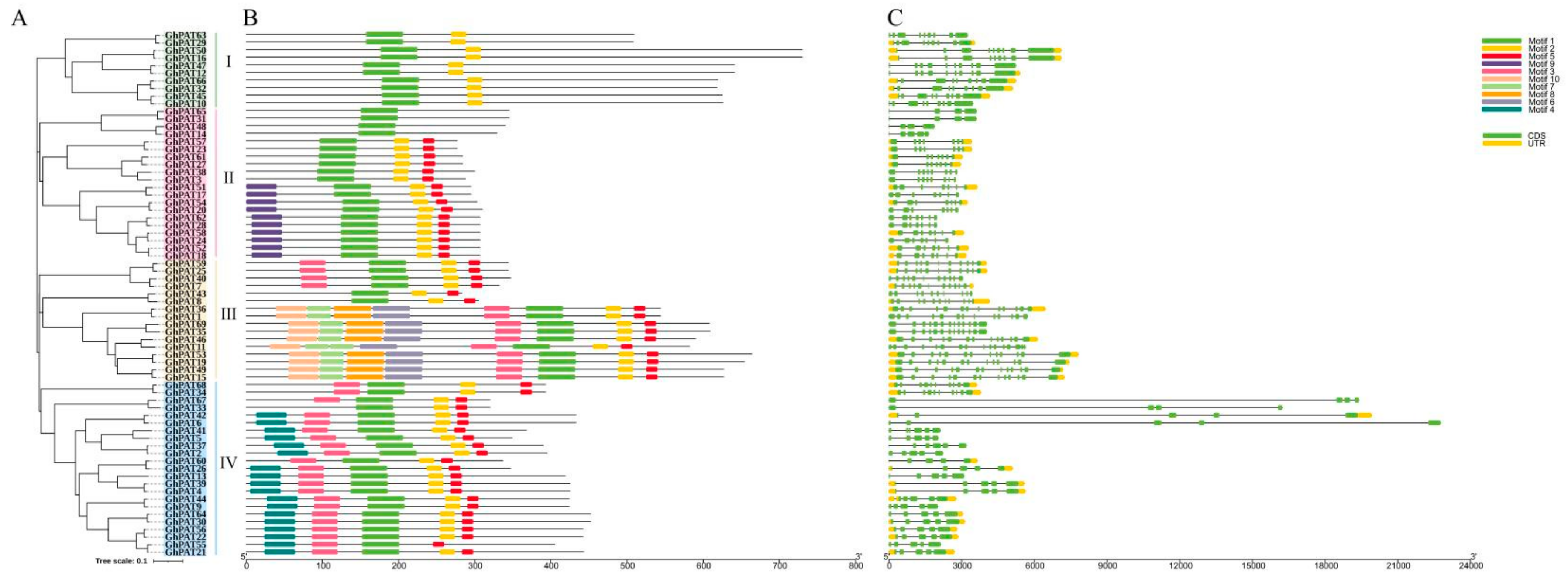
2.4. Gene Structural Analysis and Motif Prediction of GhPAT Proteins
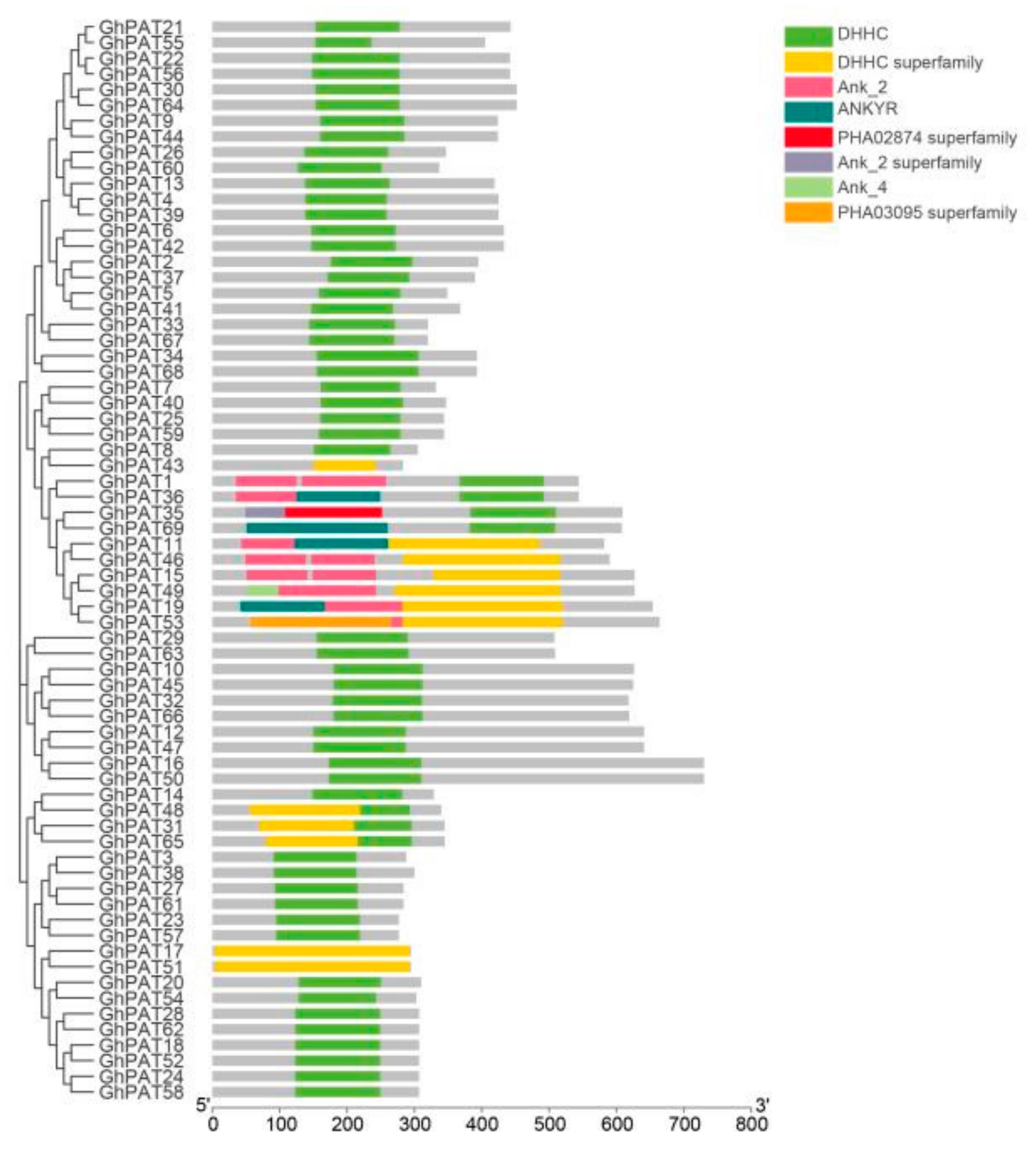
2.5. Structural Domain Analysis and Cis-Acting Element Analysis of GhPATs
2.6. Covariance Analysis of GhPATs
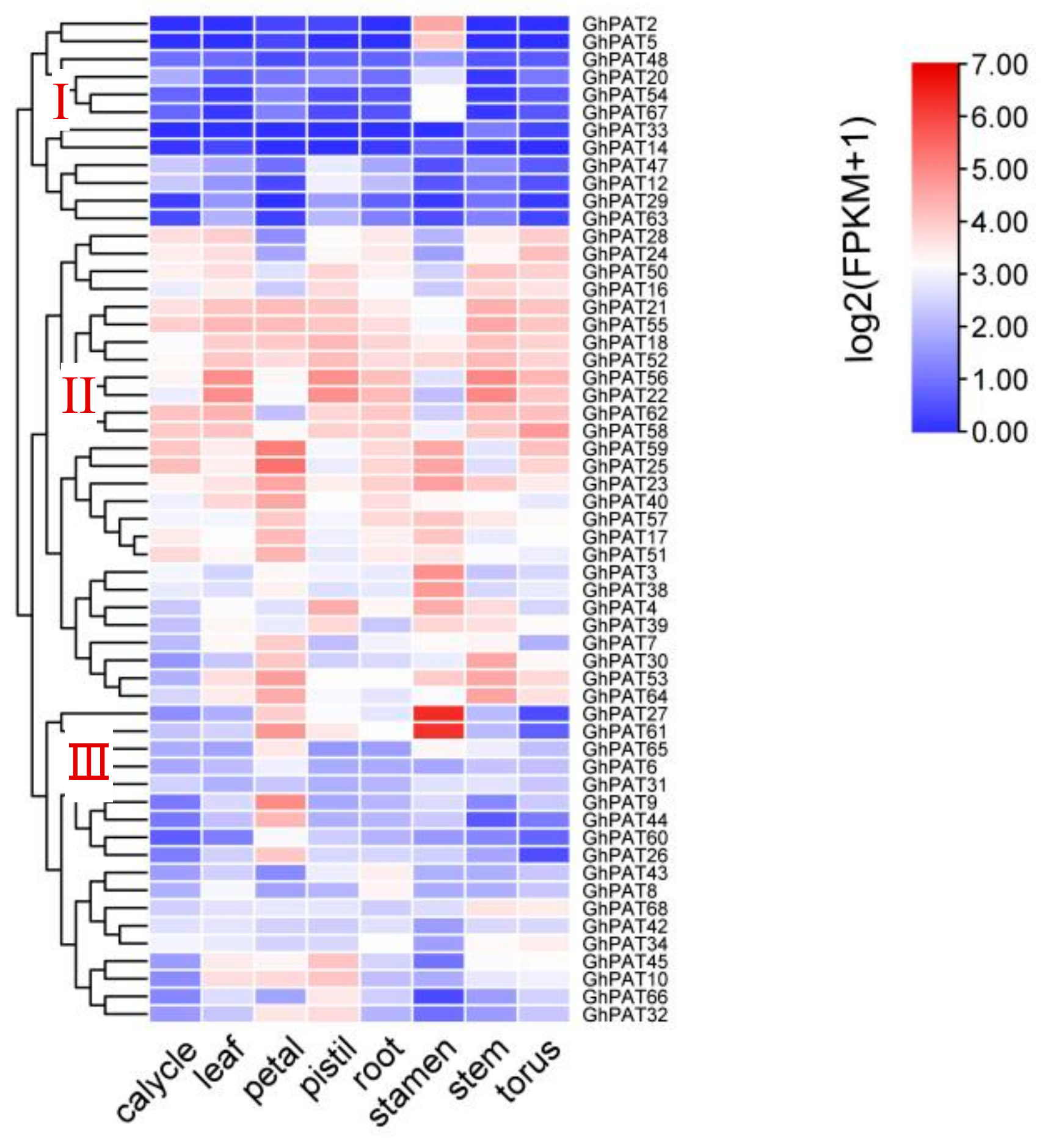
2.7. Expression Analysis of GhPATs
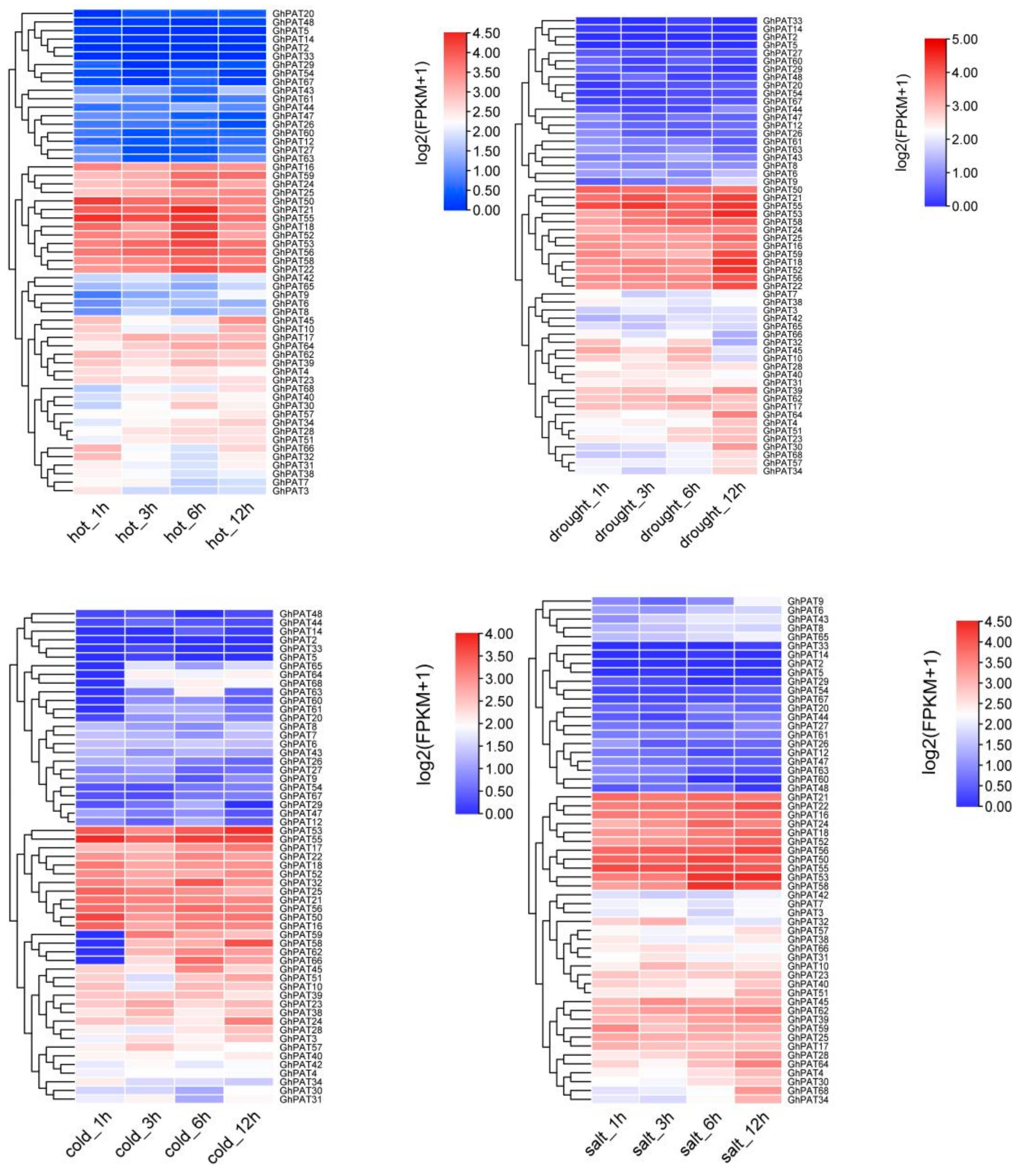
2.8. Expression of the PAT Gene Family under Different Stress Conditions in G. hirsutum
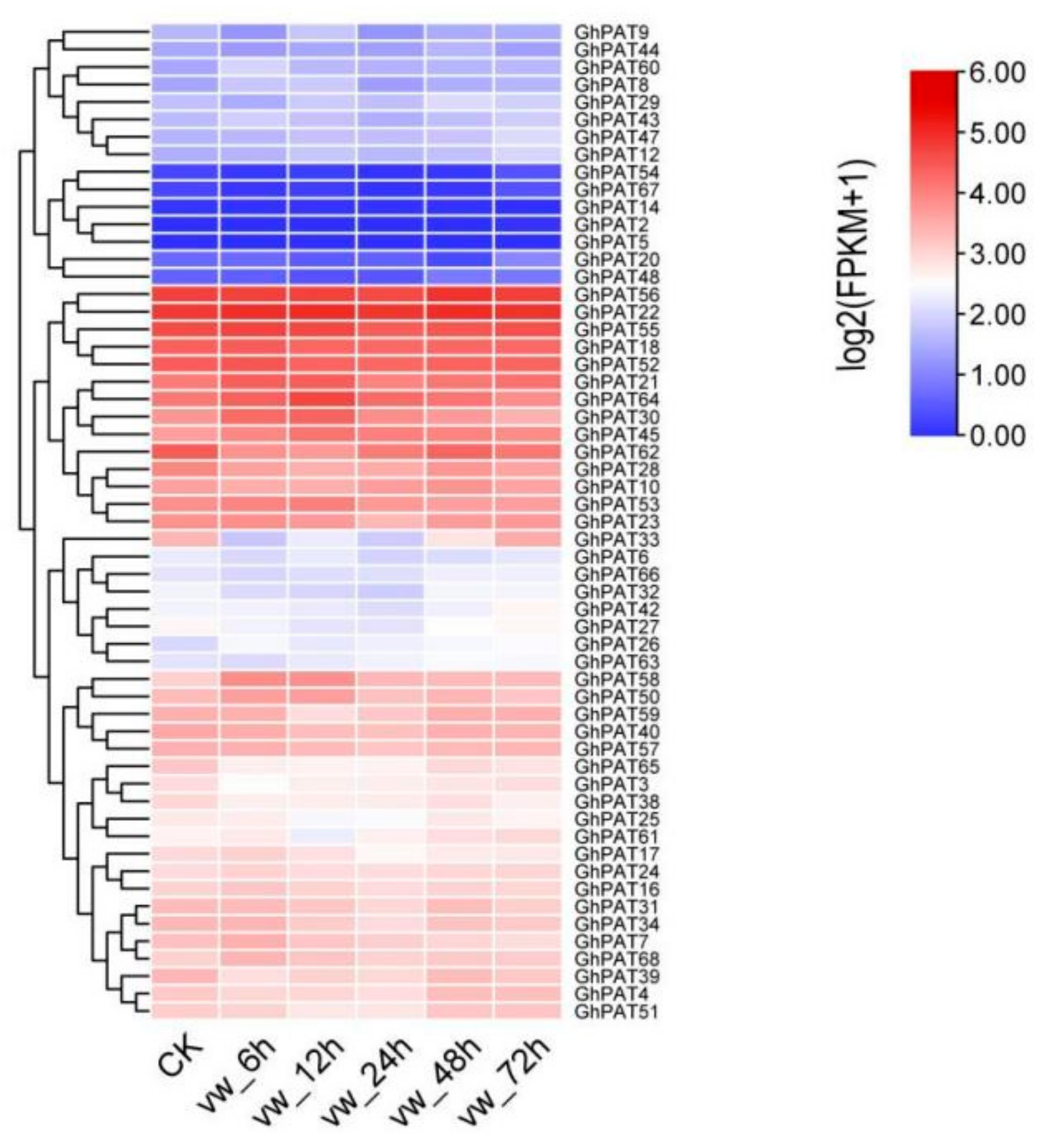
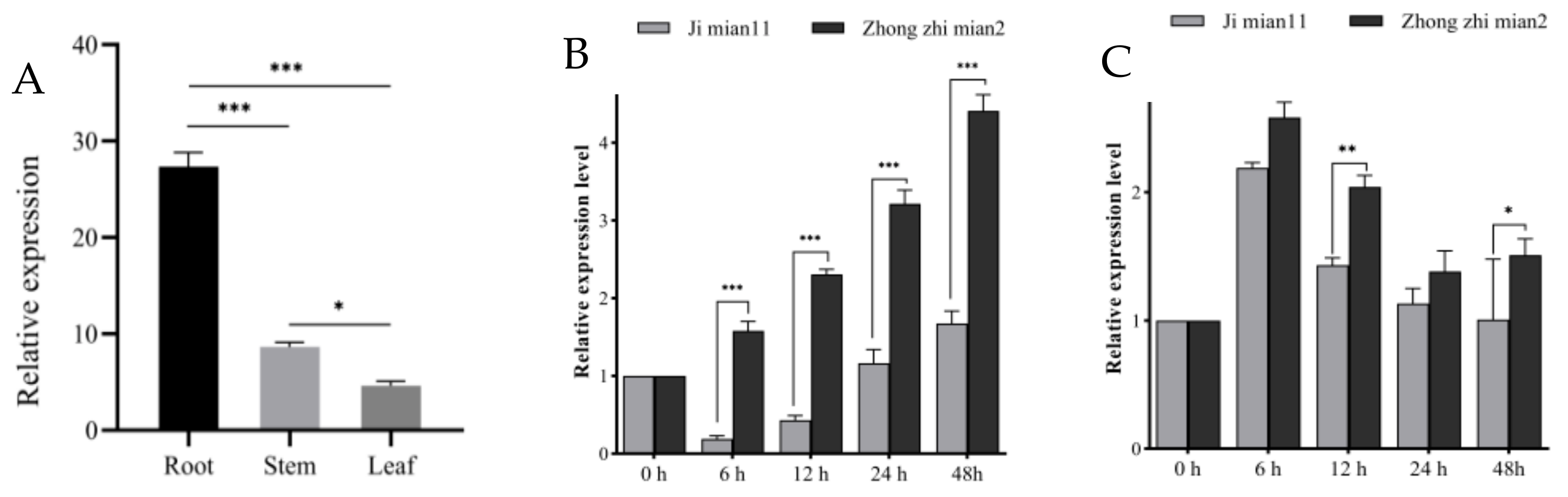
2.9. Expression Analysis of GhPATs in Verticillium Dahliae
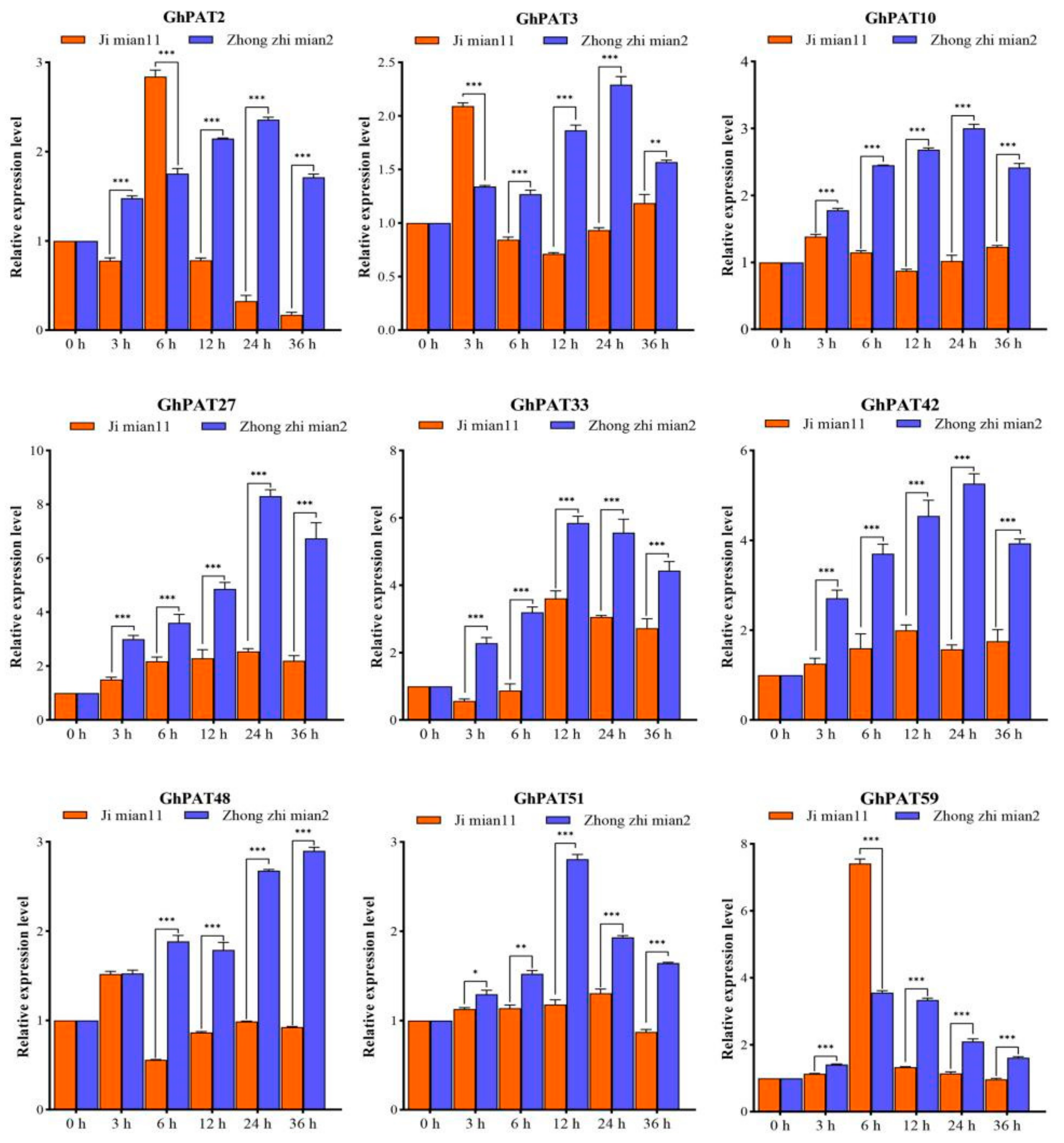
2.10. qRT-PCR to Response of GhPAT Genes to V. dahliae
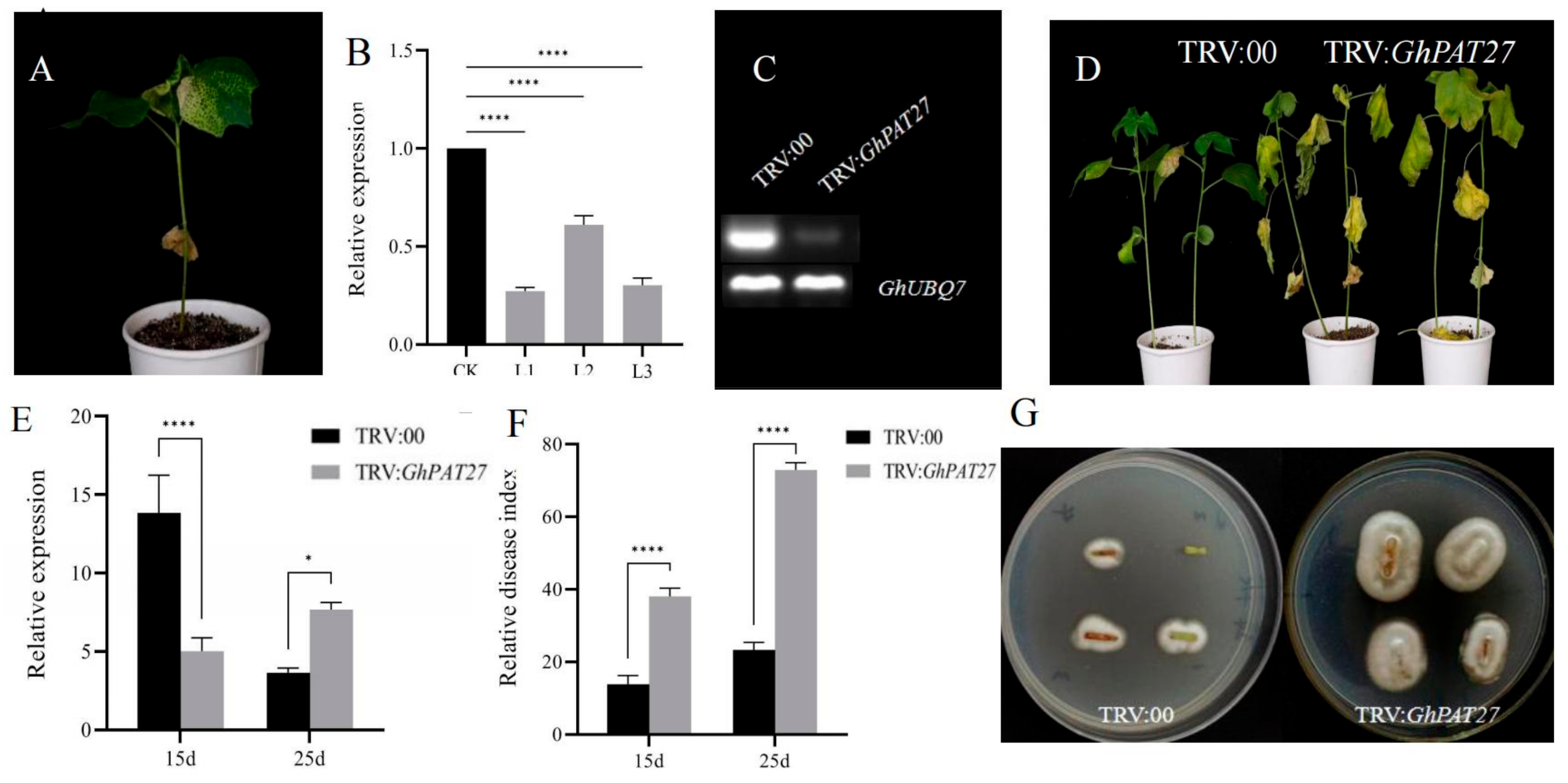
2.11. Silencing GhPAT27
3. Discussion
4. Materials and Methods
4.1. Identification and Protein Features
4.2. Chromosomal Distribution of GhPATs
4.3. Phylogenetic Analysis of GhPATs
4.4. Conserved Motif and Gene Structure Analysis of GhPATs
4.5. Structural Domain and Cis-Acting Element Analysis of GhPATs
4.6. Covariance Analysis of GhPATs
4.7. Expression Analysis of GhPATs in Cotton under Different Tissues and Stresses
4.8. Analysis of the Disease Resistance Expression Pattern of GhPATs
4.9. RNA Extraction and qRT-PCR
4.10. Silencing the Target Gene GhPAT27
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, N.; Sun, Y.; Pei, Y.; Zhang, X.Y.; Wang, P.; Li, X.; Li, F.; Hou, Y. A Pectin methylesterase inhibitor enhances resistance to Verticillium Wilt. Plant Physiol. 2018, 176, 2202–2220. [Google Scholar] [CrossRef] [Green Version]
- Qanmber, G.; Liu, J.; Yu, D.; Liu, Z.; Lu, L.; Mo, H.; Ma, S.; Wang, Z.; Yang, Z. Genome-wide identification and characterization of the PERK gene family in Gossypium hirsutum reveals gene duplication and functional divergence. Int. J. Mol. Sci. 2019, 20, 1750–1769. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Zhang, X.; Deng, F.; Yuan, R.; Shen, F. Genome-wide characterization and expression analyses of superoxide dismutase (SOD) genes in Gossypium hirsutum. BMC Genom. 2017, 18, 376–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, P.; Zhang, Y.; Xiao, G. Origin of a subgenome and genome evolution of allotetraploid cotton species. Mol. Plant 2020, 13, 1238–1240. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Xie, M.; Wang, X.; Wang, G.; Zhang, Y.; Li, Z.; Ma, Z. Identification of cell wall-associated kinases as important regulators involved in Gossypium hirsutum resistance to Verticillium dahliae. BMC Plant Biol. 2021, 21, 220–235. [Google Scholar] [CrossRef]
- Hemsley, P.A. Progress in understanding the mechanisms and functional importance of protein-membrane interactions in plants. New Phytol. 2014, 204, 741–753. [Google Scholar] [CrossRef]
- Das, T.; Yount, J.S.; Hang, H.C. Protein S-palmitoylation in immunity. Open Biol. 2021, 11, 200411. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.J.; Han, Y.Y.; Zhou, J.W. Research progress of palmitoylated proteins and palmitoylation of proteins. Subtrop. Plant Sci. 2018, 47, 395–403. [Google Scholar]
- Thinon, E.; Fernandez, J.P.; Molina, H.; Hang, H.C. Selective enrichment and direct analysis of protein S-palmitoylation sites. J. Proteome Res. 2018, 17, 1907–1922. [Google Scholar] [CrossRef]
- Yu, B.Y. Study on the Function of Arabidopsis Palmitoyl Transferases PAT13 and PAT14 in Regulating Leaf Senescence. Doctoral Dissertation, South China Normal University, Guangzhou, China, 2016. [Google Scholar]
- Li, Y.; Qi, B. Progress toward understanding protein S-acylation: Prospective in plants. Front. Plant Sci. 2017, 8, 346–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaibhav, S.; Weber, J.R.; Erik, M.; Fouke, B.W.; Vincent, B. Proteomic analysis of a poplar cell suspension culture suggests a major role of protein S-acylation in diverse cellular processes. Front. Plant Sci. 2016, 7, 477–489. [Google Scholar]
- Batisti, O. Genomics and localization of the Arabidopsis DHHC-cysteine-rich domain acyltransferase protein family. Plant Physiol. 2012, 160, 1597–1612. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Carr, P.; Turner, S.R. An atlas ofarabidopsisprotein S-acylation reveals its widespread role in plant cell organization and function. Nat. Plants 2022, 8, 670–681. [Google Scholar] [CrossRef] [PubMed]
- Hemsley, P.A.; Weimar, T.; Lilley, K.S.; Dupree, P.; Grierson, C.S. A proteomic approach identifies many novel palmitoylated proteins in Arabidopsis. New Phytol. 2013, 197, 805–814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Zhang, C.; Tang, R.J.; Xu, H.X.; Lan, W.Z.; Zhao, F.; Luan, S. Calcineurin B-Like proteins CBL4 and CBL10 mediate two independent salt tolerance pathways in Arabidopsis. Int. J. Mol. Sci. 2019, 20, 2421–2435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linder, M.E.; Deschenes, R.J.; Mitchell, D.A.; Vasudevan, A. Protein palmitoylation by a family of DHHC protein S-acyltransferases. J. Lipid Res. 2013, 47, 1118–1127. [Google Scholar]
- Rana, M.S.; Lee, C.J.; Banerjee, A. The molecular mechanism of DHHC protein acyltransferases. Biochem. Soc. Trans. 2019, 47, 157–167. [Google Scholar] [CrossRef]
- Li, Y.; Xue, J.; Wang, F.Z.; Huang, X.; Gong, B.Q.; Tao, Y.; Shen, W.; Tao, K.; Yao, N.; Xiao, S. Plasma membrane-nucleo-cytoplasmic coordination of a receptor-like cytoplasmic kinase promotes EDS1-dependent plant immunity. Nat. Plants. 2022, 8, 802–816. [Google Scholar] [CrossRef]
- Fang, L.; Gong, H.; Hu, Y.; Liu, C.; Zhou, B.; Huang, T.; Wang, Y.; Chen, S.; Fang, D.D.; Du, X. Genomic insights into divergence and dual domestication of cultivated allotetraploid cottons. Genome Biol. 2017, 18, 33–56. [Google Scholar] [CrossRef] [Green Version]
- Pang, H.G. Identification of Palmitoyltransferase Gene Family and Functional Study of PbPAT14 in Pear. Doctoral Dissertation, Hebei Agricultural University, Baoding, China, 2020. [Google Scholar]
- Qian, W.; Zhang, J. Genomic evidence for adaptation by gene duplication. Genome Res. 2014, 24, 1356–1362. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.J.; Wu, A.M.; Wei, H.L.; Hao, P.B.; Zhang, Q.; Tian, M.M.; Yang, X.; Cheng, S.S.; Fu, X.K.; Ma, L.; et al. Genome-wide identification and expression patterns analysis of the RPD3/HDA1 gene family in cotton. BMC Genom. 2020, 21, 643. [Google Scholar]
- Lai, J.; Yu, B.; Cao, Z.; Chen, Y.; Wu, Q.; Huang, J.; Yang, C. Two homologous protein S-acyltransferases, PAT13 and PAT14, cooperatively regulate leaf senescence in Arabidopsis. J. Exp. Bot. 2015, 66, 6345–6353. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Liu, P.; Liu, Q.; Wang, T.; Dong, J. Dynamic protein S-acylation in plants. Int. J. Mol. Sci. 2019, 20, 560–579. [Google Scholar] [CrossRef] [Green Version]
- Lan, T.; Delalande, C.; Dickinson, B.C. Inhibitors of DHHC family proteins. Curr. Opin. Chem. Biol. 2021, 65, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Wightman, R.; Atanassov, I.; Gupta, A.; Hurst, C.H.; Hemsley, P.A.; Turner, S. S-acylation of the cellulose synthase complex is essential for its plasma membrane localization. Science 2016, 353, 166–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greaves, J.; Carmichael, J.A.; Chamberlain, L.H. The palmitoyl transferase DHHC2 targets a dynamic membrane cycling pathway: Regulation by a C-terminal domain. Mol. Biol. Cell. 2011, 22, 1887–1895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, Y.; Zeng, H.; Wu, J.; Huang, J.; Gao, Q.; Tang, D.; Cai, L.; Liao, Z.; Wang, Y.; Liu, X.; et al. Screening dhhcs of S-acylated proteins using an OsDHHC cDNA library and bimolecular fluorescence complementation in rice. Plant J. 2022, 110, 1763–1780. [Google Scholar] [CrossRef]
- De, I.; Sadhukhan, S. Emerging roles of DHHC-mediated protein S-palmitoylation in physiological and pathophysiological context. Eur. J. Cell Biol. 2018, 97, 319–338. [Google Scholar] [CrossRef]
- Wan, Z.Y.; Chai, S.; Ge, F.R.; Feng, Q.N.; Zhang, Y.; Li, S. Arabidopsis protein S-acylation transferase4 mediates root hair growth. Plant J. 2017, 90, 249–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imran, M.; Shafiq, S.; Naeem, M.K.; Widemann, E.; Munir, M.Z.; Jensen, K.B.; Wang, R.R.-C. Histone deacetylase (HDAC) gene family in allotetraploid cotton and its diploid progenitors: In silico identification, molecular characterization, and gene expression analysis under multiple abiotic stresses, DNA damage and phytohormone treatments. Int. J. Mol. Sci. 2020, 21, 321. [Google Scholar] [CrossRef] [Green Version]
- Qin, A.; Lin, X.; Immunology, D.; Zun, Y. Advances in study of protein palmitoylation. Chem. Life 2012, 35, 36–49. [Google Scholar]
- Inoue, M.; Horimoto, K. Relationship between regulatory pattern of gene expression level and gene function. PLoS ONE 2017, 12, e0177430. [Google Scholar] [CrossRef] [Green Version]
- Lescot, M. Plantcare, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Zhou, J.M.; Zhang, Y. Plant immunity: Danger perception and signaling. Cell 2020, 181, 978–989. [Google Scholar] [CrossRef]
- Chen, X.; Ding, Y.; Yang, Y.; Song, C.; Wang, B.; Yang, S.; Guo, Y.; Gong, Z. Protein kinases in plant responses to drought, salt, and cold stress. J. Integr. Plant Biol. 2021, 63, 53–78. [Google Scholar] [CrossRef]
- Gao, J.; Huang, G.; Chen, X.; Zhu, Y. Protein S-acyl transferase 13/16 modulate disease resistance by S-acylation of the nucleotide binding, leucine-rich repeat protein R5L1 in Arabidopsis. J. Integr. Plant Biol. 2022, 64, 1789–1802. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, A. Genome-wide expression analysis reveals involvement of asparagine synthetase family in cotton development and nitrogen metabolism. BMC Plant Biol. 2022, 22, 122–135. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.Z.; Hu, Y.; Jiang, W.K.; Fang, L.; Guan, X.Y.; Li, R.Q.; Chen, Z.J. Sequencing of allotetraploid cotton (Gossypium hirsutum L. acc. TM-1) provides a resource for fiber improvement. Nat. Biotechnol. 2015, 33, 531–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duvaud, S.; Gabella, C.; Lisacek, F. Expasy, the Swiss Bioinformatics Resource Portal, as designed by its users. Nucleic Acids Res. 2021, 49, 216–227. [Google Scholar] [CrossRef]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.P.; Zhang, J.L.; Sun, Z.R.; Liu, X.Y.; Shu, L.Z.; Wu, H.; Song, Y.; He, D.H. Genome-wide identification and characterization of terpene synthase genes in Gossypium hirsutum. Gene 2022, 828, 146462. [Google Scholar] [CrossRef]
- Nystrom, S.L.; Mckay, D.J. Memes: A motif analysis environment in R using tools from the MEME suite. PLoS Comput. Biol. 2021, 6, 326–349. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative Toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Jung, S.; Cheng, C.H.; Ficklin, S.P.; Lee, T.; Zheng, P.; Jones, D.; Percy, R.G.; Main, D. CottonGen: A genomics, genetics and breeding database for cotton research. Nucleic Acids Res. 2013, 42, 1219–1236. [Google Scholar] [CrossRef] [Green Version]
- Zhu, T.; Liang, C.; Meng, Z. CottonFGD: An integrated functional genomics database for cotton. BMC Plant Biol. 2017, 17, 101. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.B.; Wang, X.P.; Wang, Y.C.; Chen, Y.H.; Luo, J.W.; Li, D.D.; Li, X.B. Genome-wide identification and functional characterization of cotton (Gossypium hirsutum) MAPKKK gene family in response to drought stress. BMC Plant Biol. 2020, 20, 217. [Google Scholar] [CrossRef]
- Mustafa, R.; Shafiq, M.; Mansoor, S.; Briddon, R.W.; Scheffler, B.E.; Scheffler, J.; Amin, I. Virus-induced gene silencing in cultivated cotton (Gossypium spp.) using Tobacco rattle virus. Mol. Biotechnol. 2016, 58, 65–72. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Name | Genomic Location | Protein Length (bp) | Molecular Weight (Da) | Theoretical Isoelectric Point (pI) |
|---|---|---|---|---|---|
| Gh_A01G201000 | GhPAT1 | A01: 106,644,168–106,649,891 | 544 | 60,329.10 | 7.67 |
| Gh_A01G217400 | GhPAT2 | A01: 110,355,592–110,357,821 | 395 | 44,677.10 | 8.13 |
| Gh_A01G248800 | GhPAT3 | A01: 114,993,971–114,996,727 | 288 | 32,621.46 | 8.74 |
| Gh_A02G074800 | GhPAT4 | A02: 10,783,870–10,789,492 | 425 | 48,254.43 | 7.95 |
| Gh_A02G139500 | GhPAT5 | A02: 77,013,588–77,015,623 | 349 | 39,891.37 | 9.09 |
| Gh_A02G139600 | GhPAT6 | A02: 77,326,344–77,349,111 | 433 | 49,247.55 | 6.5 |
| Gh_A02G207100 | GhPAT7 | A02: 105,408,865–105,412,345 | 332 | 38,266.80 | 8.56 |
| Gh_A03G054400 | GhPAT8 | A03: 8,316,187–8,320,341 | 305 | 34,420.65 | 8.48 |
| Gh_A04G122000 | GhPAT9 | A04: 77,576,070–77,578,084 | 424 | 48,357.21 | 8.69 |
| Gh_A04G170800 | GhPAT10 | A04: 84,471,911–84,475,376 | 626 | 68,684.92 | 8.59 |
| Gh_A05G010000 | GhPAT11 | A05: 1,406,399–1,412,030 | 582 | 64,090.24 | 7.09 |
| Gh_A05G162000 | GhPAT12 | A05: 17,248,575–17,253,983 | 641 | 70,277.50 | 8.38 |
| Gh_A05G258700 | GhPAT13 | A05: 30,687,878–30,690,980 | 419 | 47,780.74 | 7.48 |
| Gh_A05G301400 | GhPAT14 | A05: 45,644,709–45,646,354 | 329 | 36,863.26 | 8.93 |
| Gh_A06G057800 | GhPAT15 | A06: 9,253,637–9,260,874 | 627 | 69,025.32 | 6.71 |
| Gh_A07G090000 | GhPAT16 | A07: 12,680,862–12,687,974 | 730 | 79,039.88 | 8.56 |
| Gh_A07G144800 | GhPAT17 | A07: 29,190,890–29,193,768 | 295 | 33,122.02 | 8.21 |
| Gh_A07G158400 | GhPAT18 | A07: 35,012,548–35,015,765 | 307 | 34,645.40 | 7.69 |
| Gh_A08G096600 | GhPAT19 | A08: 31,121,482–31,128,915 | 654 | 71,782.47 | 7.21 |
| Gh_A08G198400 | GhPAT20 | A08: 114,519,379–114,522,238 | 310 | 34,997.23 | 8.61 |
| Gh_A09G042400 | GhPAT21 | A09: 14,285,815–14,288,513 | 443 | 49,732.03 | 8.41 |
| Gh_A09G053600 | GhPAT22 | A09: 30,324,111–30,326,963 | 442 | 49,694.26 | 8.53 |
| Gh_A10G209400 | GhPAT23 | A10: 107,898,560–107,901,981 | 277 | 31,495.96 | 8.75 |
| Gh_A11G086800 | GhPAT24 | A11: 7,557,651–7,560,097 | 307 | 34,754.33 | 7.42 |
| Gh_A11G181600 | GhPAT25 | A11: 23,238,492–23,242,550 | 344 | 39,451.13 | 8.17 |
| Gh_A11G240600 | GhPAT26 | A11: 67,374,084–67,379,186 | 347 | 39,639.16 | 8.57 |
| Gh_A11G375800 | GhPAT27 | A11: 119,164,180–119,167,139 | 284 | 31,907.41 | 8.53 |
| Gh_A12G117500 | GhPAT28 | A12: 75,796,826–75,798,806 | 307 | 34,960.57 | 6.59 |
| Gh_A12G137100 | GhPAT29 | A12: 80,379,332–80,382,877 | 508 | 58,141.21 | 10.27 |
| Gh_A12G216400 | GhPAT30 | A12: 96,932,476–96,935,605 | 452 | 51,452.17 | 8.41 |
| Gh_A13G025000 | GhPAT31 | A13: 2,645,301–2,648,903 | 345 | 38,583.93 | 8.94 |
| Gh_A13G084800 | GhPAT32 | A13: 22,978,902–22,984,011 | 618 | 68,051.95 | 8.51 |
| Gh_A13G107100 | GhPAT33 | A13: 53,794,071–53,810,304 | 320 | 36,354.90 | 8.54 |
| Gh_A13G196000 | GhPAT34 | A13: 101,505,059–101,508,851 | 393 | 44,823.33 | 9.13 |
| Gh_A13G228400 | GhPAT35 | A13: 105,162,566–105,166,602 | 609 | 66,921.37 | 7.43 |
| Gh_D01G198500 | GhPAT36 | D01: 57,318,599–57,325,051 | 544 | 60,259.05 | 7.67 |
| Gh_D01G213700 | GhPAT37 | D01: 59,934,237–59,937,432 | 390 | 44,173.59 | 8.33 |
| Gh_D01G243100 | GhPAT38 | D01: 63,762,802–63,765,621 | 300 | 34,025.02 | 9.28 |
| Gh_D02G077600 | GhPAT39 | D02: 9,776,929–9,782,516 | 425 | 48,308.45 | 7.99 |
| Gh_D03G003900 | GhPAT40 | D03: 257,592–260,644 | 347 | 39,868.76 | 8.2 |
| Gh_D03G067200 | GhPAT41 | D03: 15,062,867–15,064,986 | 368 | 42,022.34 | 8.93 |
| Gh_D03G086100 | GhPAT42 | D03: 29,483,457–29,503,379 | 433 | 49,139.33 | 6.19 |
| Gh_D03G142600 | GhPAT43 | D03: 46,045,864–46,049,299 | 283 | 32,108.88 | 7.85 |
| Gh_D04G161600 | GhPAT44 | D04: 49,275,947–49,278,715 | 424 | 48,386.27 | 8.9 |
| Gh_D04G217000 | GhPAT45 | D04: 56,134,857–56,139,025 | 625 | 68,509.73 | 8.59 |
| Gh_D05G014000 | GhPAT46 | D05: 1,289,795–1,295,927 | 590 | 65,581.87 | 7.02 |
| Gh_D05G179000 | GhPAT47 | D05: 15,635,522–15,640,760 | 641 | 70,308.72 | 8.26 |
| Gh_D05G310400 | GhPAT48 | D05: 35,421,295–35,423,177 | 340 | 37,957.65 | 8.65 |
| Gh_D06G056100 | GhPAT49 | D06: 7,676,920–7,684,097 | 627 | 69,085.33 | 6.74 |
| Gh_D07G088700 | GhPAT50 | D07: 10,795,175–10,802,285 | 730 | 78,955.76 | 8.39 |
| Gh_D07G143900 | GhPAT51 | D07: 21,950,634–21,954,273 | 295 | 32,994.85 | 8.3 |
| Gh_D07G157400 | GhPAT52 | D07: 25,658,584–25,661,866 | 307 | 34,660.37 | 7.69 |
| Gh_D08G098900 | GhPAT53 | D08: 23,438,004–23,445,817 | 664 | 72,891.51 | 7.19 |
| Gh_D08G194900 | GhPAT54 | D08: 58,917,795–58,921,035 | 303 | 34,398.53 | 8.47 |
| Gh_D09G037600 | GhPAT55 | D09: 10,675,800–10,677,929 | 405 | 45,321.87 | 8.42 |
| Gh_D09G045200 | GhPAT56 | D09: 10,675,800–10,677,929 | 442 | 49,607.18 | 8.47 |
| Gh_D10G232300 | GhPAT57 | D10: 59,176,702–59,180,114 | 277 | 31,577.09 | 8.75 |
| Gh_D11G087400 | GhPAT58 | D11: 7,193,621–7,196,712 | 307 | 34,806.38 | 7.42 |
| Gh_D11G183900 | GhPAT59 | D11: 19,600,308–19,604,330 | 344 | 39,533.21 | 8.09 |
| Gh_D11G241300 | GhPAT60 | D11: 39,646,391–39,650,033 | 337 | 38,582.85 | 8.37 |
| Gh_D11G383600 | GhPAT61 | D11: 72,301,279–72,304,312 | 284 | 31,960.38 | 8.53 |
| Gh_D12G117600 | GhPAT62 | D12: 39,003,340–39,005,322 | 307 | 35,016.57 | 6.41 |
| Gh_D12G137300 | GhPAT63 | D12: 42,848,700–42,851,950 | 509 | 58,398.57 | 10.3 |
| Gh_D12G209200 | GhPAT64 | D12: 54,340,099–54,343,145 | 452 | 51,129.85 | 8.64 |
| Gh_D13G026800 | GhPAT65 | D13: 2,399,443–2,403,053 | 345 | 38,584.87 | 8.69 |
| Gh_D13G087200 | GhPAT66 | D13: 15,254,636–15,259,872 | 619 | 68,219.12 | 8.47 |
| Gh_D13G098700 | GhPAT67 | D13: 20,048,482–20,067,879 | 320 | 36,331.89 | 8.47 |
| Gh_D13G198500 | GhPAT68 | D13: 56,689,778–56,693,401 | 393 | 44,679.16 | 8.97 |
| Gh_D13G232500 | GhPAT69 | D13: 60,344,782–60,348,822 | 608 | 67,203.66 | 7.43 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, Y.; Chen, Q.; Qu, Y. Protein S-Acyl Transferase GhPAT27 Was Associated with Verticillium wilt Resistance in Cotton. Plants 2022, 11, 2758. https://doi.org/10.3390/plants11202758
Deng Y, Chen Q, Qu Y. Protein S-Acyl Transferase GhPAT27 Was Associated with Verticillium wilt Resistance in Cotton. Plants. 2022; 11(20):2758. https://doi.org/10.3390/plants11202758
Chicago/Turabian StyleDeng, Yahui, Quanjia Chen, and Yanying Qu. 2022. "Protein S-Acyl Transferase GhPAT27 Was Associated with Verticillium wilt Resistance in Cotton" Plants 11, no. 20: 2758. https://doi.org/10.3390/plants11202758
APA StyleDeng, Y., Chen, Q., & Qu, Y. (2022). Protein S-Acyl Transferase GhPAT27 Was Associated with Verticillium wilt Resistance in Cotton. Plants, 11(20), 2758. https://doi.org/10.3390/plants11202758