

New Proposal of Epiphytic Bromeliaceae Functional Groups to Include Nebulophytes and Shallow Tanks

 , , , ,
, , , ,  , ,
, ,  , , ,
, , ,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
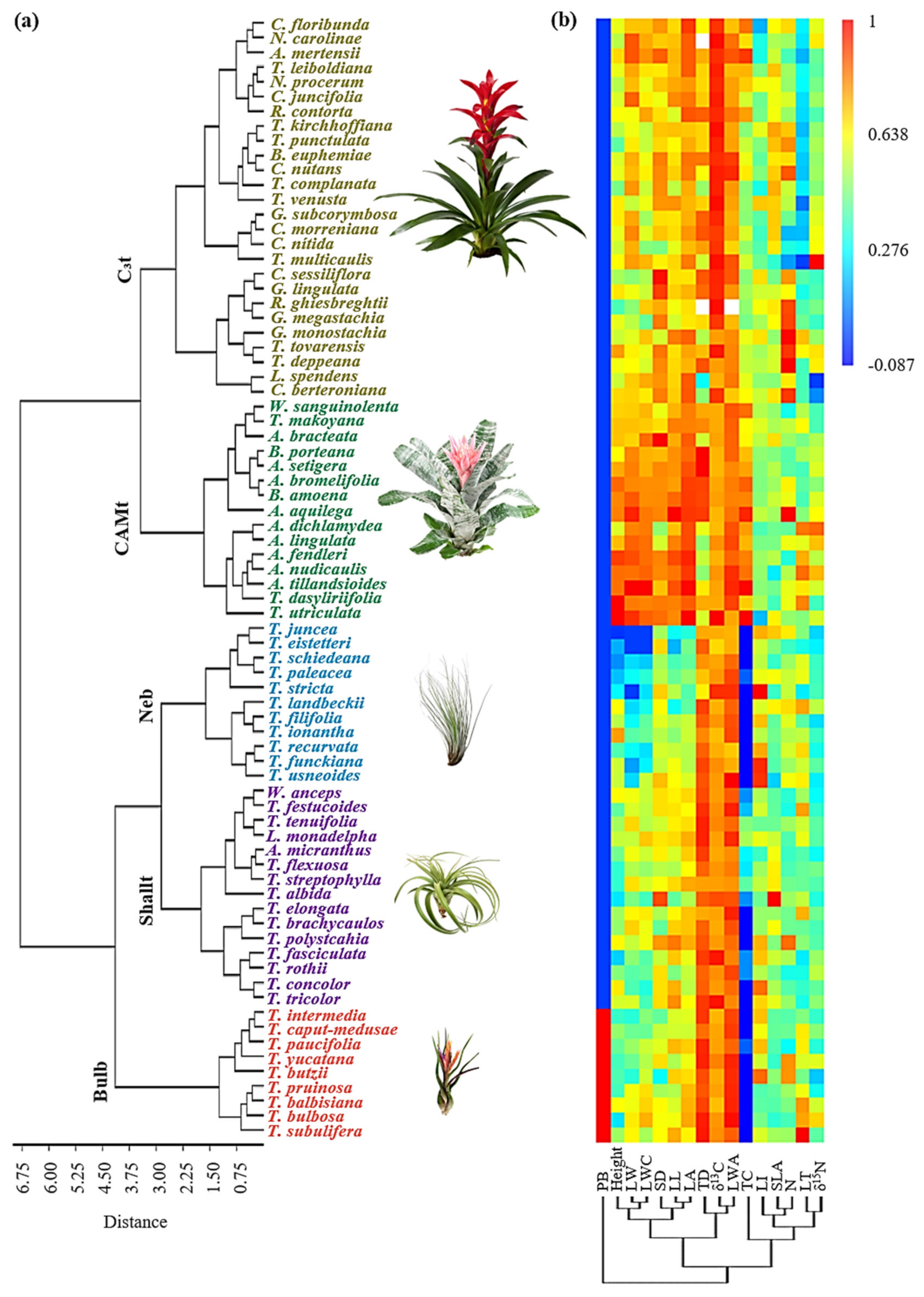
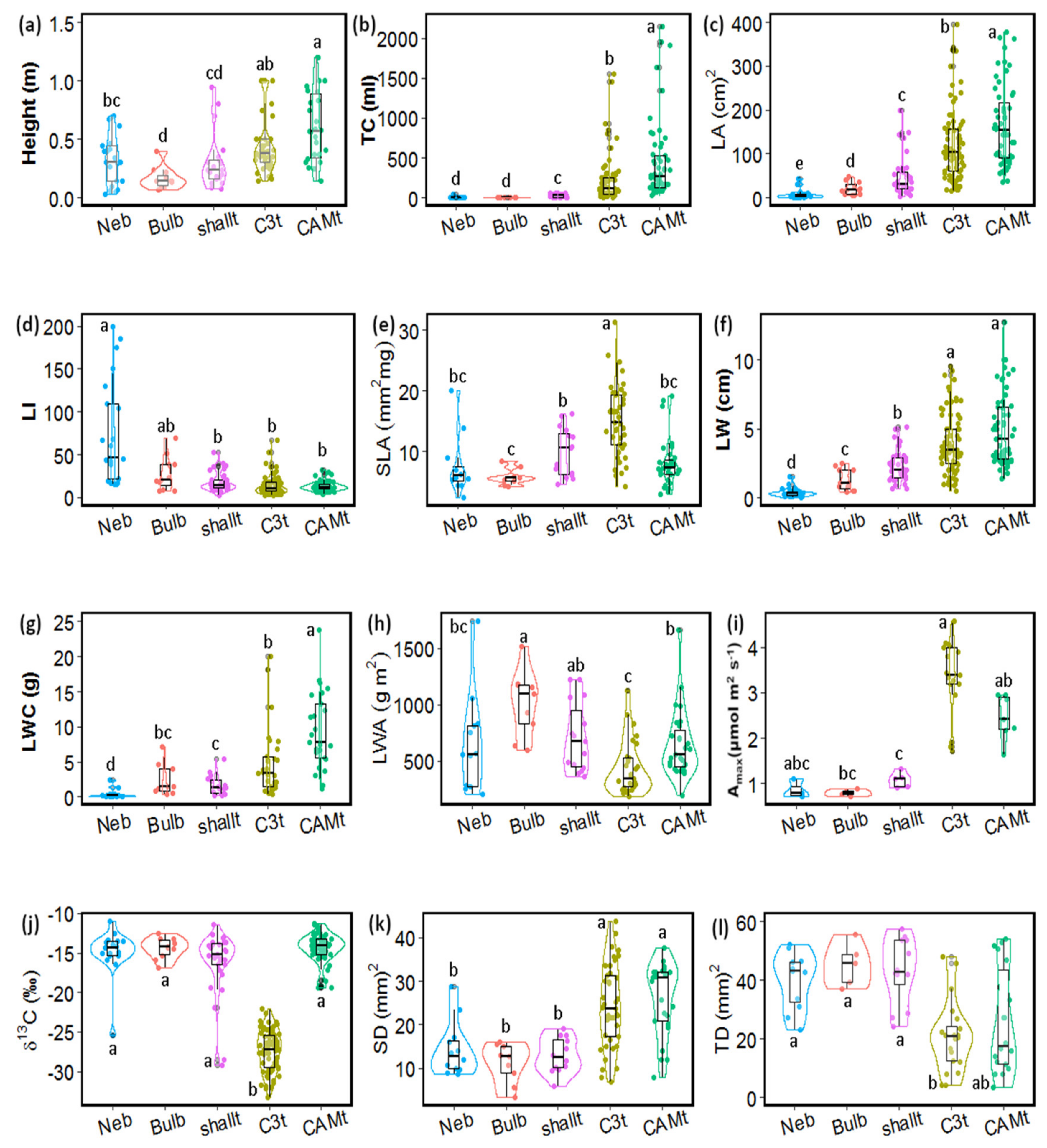
2.1. Functional Groups and Associated Functional Traits
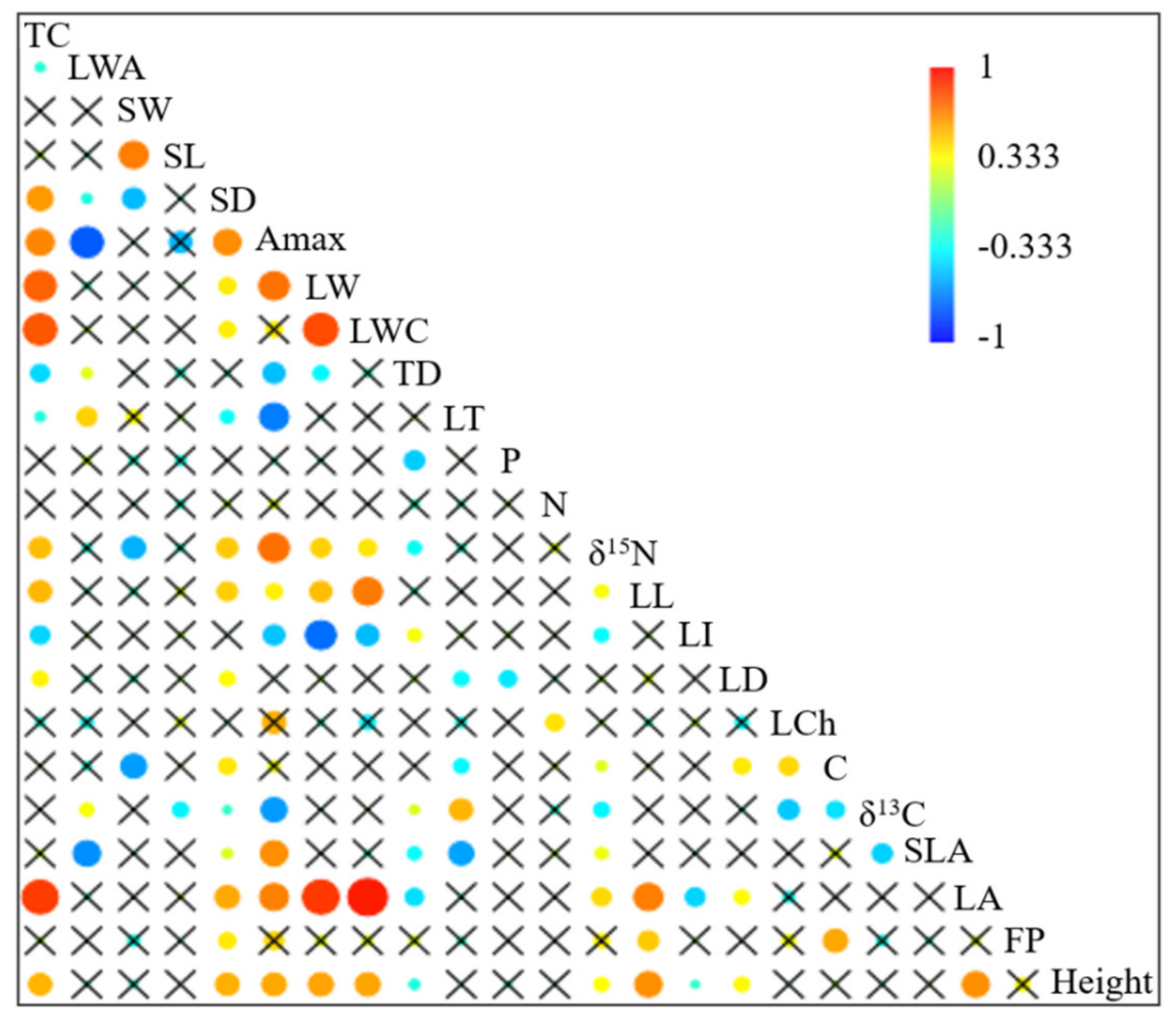
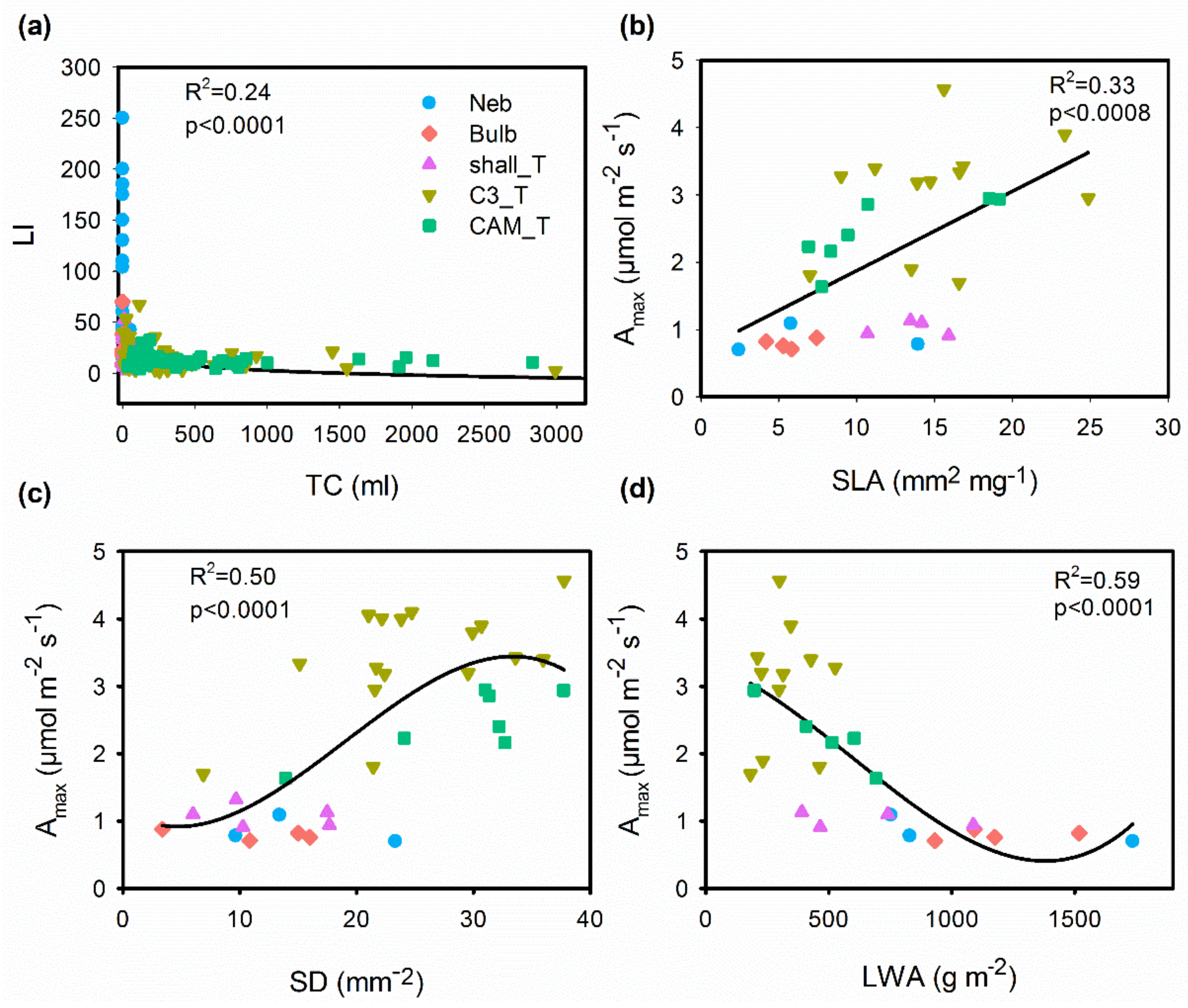
2.2. Correlations between Functional Traits
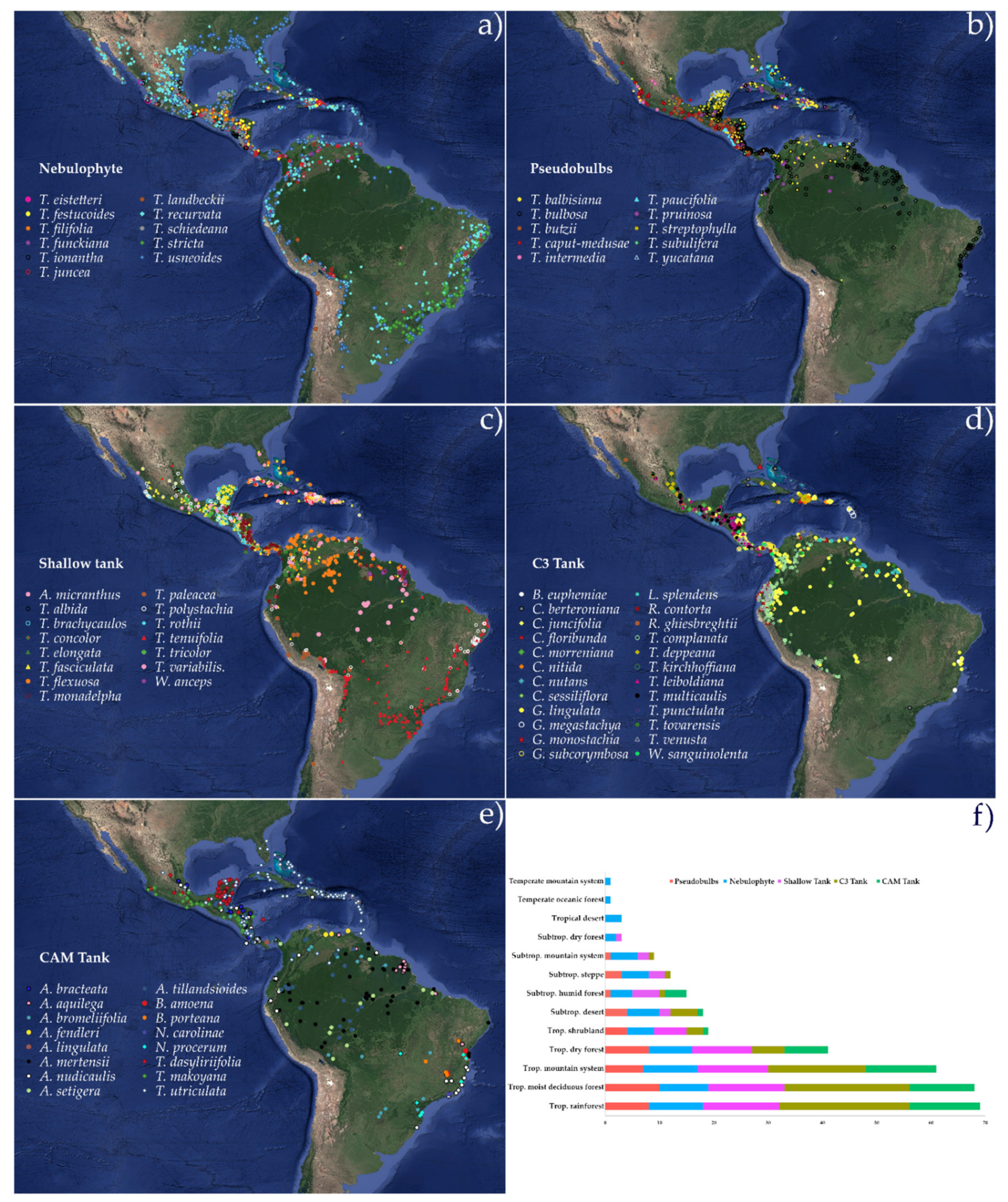
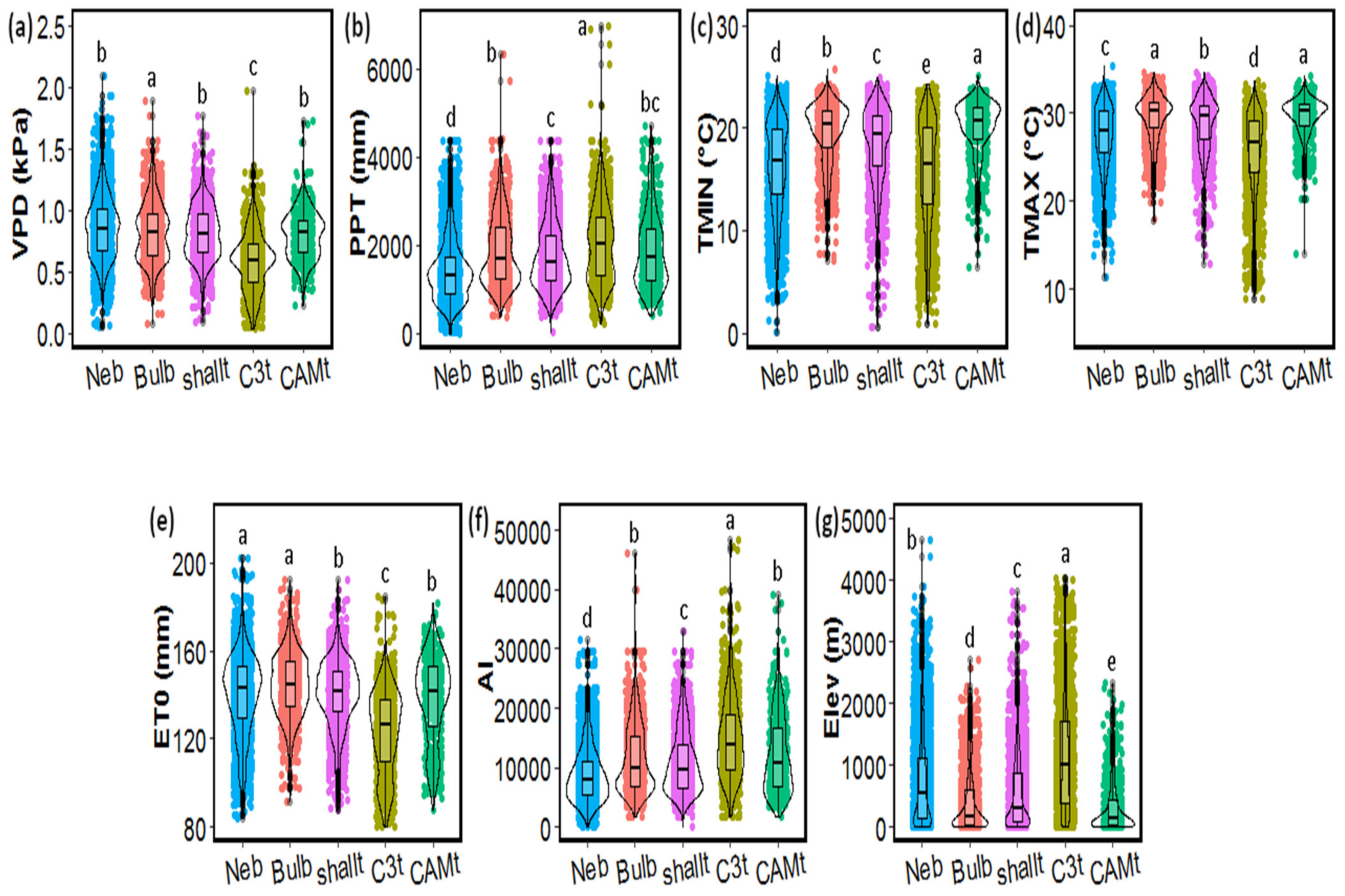
2.3. Functional Groups’ Habitat and Distribution
3. Discussion
4. Materials and Methods
4.1. Trait Data
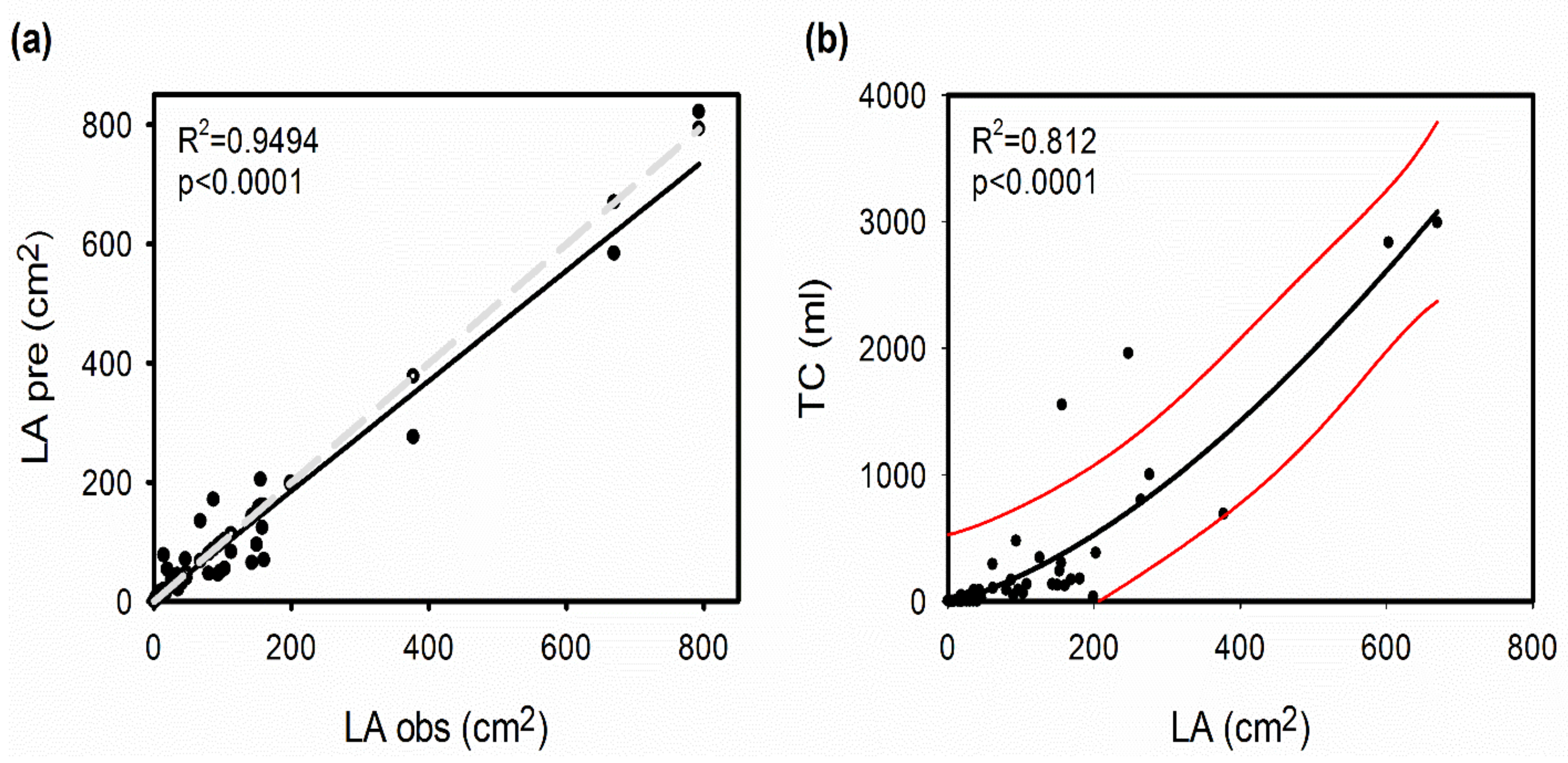
4.2. Gapfilling of Trait Data
4.3. Environmental Data
4.4. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grime, J.P. Plant Strategies and Vegetation Processes; John Wiley: Chichester, UK, 1979. [Google Scholar]
- Körner, C. Scaling from species to vegetation: The usefulness of functional groups. In Biodiversity and Ecosystem Function; Springer: Berlin/Heidelberg, Germany, 1994; pp. 117–140. [Google Scholar]
- Spake, R.; Ezard, T.H.; Martin, P.A.; Newton, A.C.; Doncaster, C.P. A meta-analysis of functional group responses to forest recovery outside of the tropics. Conser. Biol. 2015, 29, 1695–1703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westoby, M. A leaf-height-seed (LHS) plant ecology strategy scheme. Plant Soil 1998, 199, 213–227. [Google Scholar] [CrossRef]
- Donovan, L.A.; Maherali, H.; Caruso, C.M.; Huber, H.; de Kroon, H. The evolution of the worldwide leaf economics spectrum. Trends Ecol. Evol. 2011, 26, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Hietz, P.; Wagner, K.; Nunes Ramos, F.; Sarmento Cabral, J.; Agudelo, C.; Benavides, A.M.; Cach-Pérez, M.J.; Cardelús, C.L.; Chilpa-Galván, N.; Nascimento da Costa, L.E.; et al. Putting vascular epiphytes on the traits map. J. Ecol. 2022, 110, 340–358. [Google Scholar] [CrossRef]
- Lüttge, U. Setting the Scene. In Vascular Plants as Epiphytes: Evolution and Ecophysiology; Lüttge, U., Ed.; Ecological Studies; Springer: Berlin/Heidelberg, Germany, 1989; Volume 76. [Google Scholar]
- Zotz, G. Plants on Plants-The Biology of Vascular Epiphytes; Springer International Publishing: Cham, Switzerland, 2016; p. 282. [Google Scholar]
- Males, J. Think tank: Water relations of Bromeliaceae in their evolutionary context. Bot. J. Linn. Soc. 2016, 181, 415–440. [Google Scholar] [CrossRef] [Green Version]
- Hietz, P. Population dynamics of epiphytes in a Mexican humid montane forest. J. Ecol. 1997, 85, 767–775. [Google Scholar] [CrossRef]
- Sarmento Cabral, J.; Petter, G.; Mendieta-Leiva, G.; Wagner, K.; Zotz, G.; Kreft, H. Branchfall as a demographic filter for epiphyte communities: Lessons from forest floor-based sampling. PLoS ONE 2015, 10, e0128019. [Google Scholar] [CrossRef] [PubMed]
- Cortes-Anzures, B.O.; Corona-López, A.M.; Toledo-Hernández, V.H.; Valencia-Díaz, S.; Flores-Palacios, A. Branch mortality influences phorophyte quality for vascular epiphytes. Botany 2017, 95, 709–716. [Google Scholar] [CrossRef]
- Taylor, A.; Zotz, G.; Weigelt, P.; Cai, L.; Karger, D.N.; König, C.; Kreft, H. Vascular epiphytes contribute disproportionately to global centers of plant diversity. Glob. Ecol. Biogeogr. 2021, 31, 62–74. [Google Scholar] [CrossRef]
- Zotz, G.; Weigelt, P.; Kessler, M.; Kreft, H.; Taylor, A. EpiList 1.0—A global checklist of vascular epiphytes. Ecology 2021, 102, e03326. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.A.C. Epiphytic bromeliads. In Vascular Plants as Epiphytes; Lüttge, U., Ed.; Springer: Berlin/Heidelberg, Germany, 1989; pp. 109–138. [Google Scholar]
- Zizka, A.; Azevedo, J.; Leme, E.; Neves, B.; da Costa, A.F.; Caceres, D.; Zizka, G. Biogeography and conservation status of the pineapple family (Bromeliaceae). Divers. Distrib. 2020, 26, 183–195. [Google Scholar] [CrossRef]
- Pittendrigh, C.S. The bromeliad-Anopheles-malaria complex in Trinidad. I- The bromeliad flora. Evolution 1948, 2, 58–89. [Google Scholar] [PubMed] [Green Version]
- Schimper, A.F.W. Die Epiphytische Vegetation Amerikas; Fischer, J.G., Ed.; Kessinger Publishing: Berlin, Germany, 1888. [Google Scholar]
- Mez, C. Physiologische Bromeliaceen-atudien I. Die wasser-ökonomie der extrem atmosphärischen Tillandsien. Jahrbücher Für Wiss. Bot. 1904, 40, 157–229. [Google Scholar]
- Tietze, E. Physiologische Bromeliaceen-Studien II. Die entwicklung der wasseraufnehmenden Bromeliaceen-Trichome. Z. Für Nat. 1906, 78, 1–50. [Google Scholar]
- Benzing, D.H. Bromeliaceae: Profile of an Adaptive Radiation; Cambridge University Press: Cambridge, UK, 2000; p. 710. [Google Scholar]
- Benzing, D.H.; Renfrow, A. The significance of photosynthetic efficiency to habitat preference and phylogeny among tillandsioid bromeliads. Bot. Gaz. 1971, 132, 19–30. [Google Scholar] [CrossRef]
- Adams, W.W.; Martin, C.E. Physiological consequences of changes in life form of the Mexican epiphyte Tillandsia deppeana (Bromeliaceae). Oecologia 1986, 70, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Reyes-García, C.; Griffiths, H.; Rincón, E.; Huante, P. Niche differentiation in tank and atmospheric epiphytic bromeliads of a seasonally dry forest. Biotropica 2008, 40, 168–175. [Google Scholar] [CrossRef]
- Reyes-García, C.; Griffiths, H.; Mejia-Chang, M. High but not dry: Diverse epiphytic bromeliad adaptations to exposure within a seasonally dry tropical forest community. New Phytol. 2012, 193, 745–754. [Google Scholar] [CrossRef]
- Cach-Pérez, M.J.; Andrade, J.L.; Reyes-García, C. Morphophysiological plasticity in epiphytic bromeliads across a precipitation gradient in the Yucatan Peninsula, Mexico. Trop. Conserv. Sci. 2018, 11, 1940082918781926. [Google Scholar] [CrossRef]
- Chávez-Sahagún, E.; Andrade, J.L.; Zotz, G.; Reyes-García, C. Dew can prolong photosynthesis and water status during drought in some epiphytic bromeliads from a seasonally dry tropical forest. Trop. Conserv. Sci. 2019, 12, 1–11. [Google Scholar] [CrossRef]
- Griffiths, H.; Smith, J.A.C. Photosynthetic pathways in the Bromeliaceae of Trinidad: Relations between life-forms, habitat preference and the occurrence of CAM. Oecologia 1983, 60, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Mejia-Chang, M.; Reyes-Garcia, C.; Seibt, U.; Royles, J.; Meyer, M.T.; Jones, G.D.; Winter, K.; Arnedo, M.; Griffiths, H. Leaf water δ18O reflects water vapour exchange and uptake by C3 and CAM epiphytic bromeliads in Panama. Funct. Plant Biol. 2021, 48, 732–742. [Google Scholar] [CrossRef]
- Barradas, V.L.; Glez-Medellin, M.G. Dew and its effect on two heliophile understorey species of a tropical dry deciduous forest in Mexico. Int. J. Biometeorol. 1999, 43, 1–7. [Google Scholar] [CrossRef]
- Andrade, J.L. Dew deposition on epiphytic bromeliad leaves: An important event in a Mexican tropical dry deciduous forest. J. Trop. Ecol. 2003, 19, 479–488. [Google Scholar] [CrossRef]
- Nobel, P.S. Physicochemical and Environmental Plant Physiology, 4th ed.; Academic Press: San Diego, CA, USA, 2009; p. 572. [Google Scholar]
- Martorell, C.; Ezcurra, E. Rosette scrub occurrence and fog availability in arid mountains of Mexico. J. Veg. Sci. 2002, 13, 651–662. [Google Scholar] [CrossRef]
- Martorell, C.; Ezcurra, E. The narrow-leaf syndrome: A functional and evolutionary approach to the form of fog-harvesting rosette plants. Oecologia 2007, 151, 561–573. [Google Scholar] [CrossRef] [PubMed]
- Martorell, C. Morfología funcional de la rosetofilia: ¿Plantas que cosechan la neblina? Ph.D. Thesis, Universidad Nacional Autónoma de México, México DF, México, 2002. [Google Scholar]
- Rundel, P.W.; Palma, B.; Dillon, M.O.; Sharifi, M.R.; Nilsen, E.T.; Boonpragob, K. Tillandsia landbeckii in the coastal Atacama Desert of northern Chile. Rev. Chil. Hist. Nat. 1997, 70, 341–349. [Google Scholar]
- González, A.L.; Fariña, J.M.; Pinto, R.; Pérez, C.; Weathers, K.C.; Armesto, J.J.; Marquet, P.A. Bromeliad growth and stoichiometry: Responses to atmospheric nutrient supply in fog-dependent ecosystems of the hyper-arid Atacama Desert, Chile. Oecologia 2011, 167, 835–845. [Google Scholar] [CrossRef]
- Guevara-Escobar, A.; Cervantes-Jiménez, M.; Suzán-Azpiri, H.; González-Sosa, E.; Hernández-Sandoval, L.; Malda-Barrera, G.; Martínez-Díaz, M. Fog interception by Ball moss (Tillandsia recurvata). HESS 2011, 15, 2509–2518. [Google Scholar]
- Benzing, D.H. An investigation of two bromeliad myrmecophytes: Tillandsia butzii Mez, Tillandsia caput-medusae E. Morren, and their ants. Bull. Torrey Bot. Club 1970, 97, 109–115. [Google Scholar] [CrossRef]
- Chew, T.; De Luna, E.; González, D. Phylogenetic relationships of the pseudobulbous Tillandsia species (Bromeliaceae) inferred from cladistic analyses of ITS 2, 5.8 S ribosomal RNA gene, and ETS sequences. Syst. Bot. 2010, 35, 86–95. [Google Scholar] [CrossRef]
- Stefano, M.; Papini, A.; Brighigna, L. A new quantitative classification of ecological types in the bromeliad genus Tillandsia (Bromeliaceae) based on trichomes. Rev. Biol. Trop. 2008, 56, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Benzing, D.H.; Burt, K.M. Foliar permeability among twenty species of the Bromeliaceae. Bull. Torrey Bot. Club. 1970, 97, 269–279. [Google Scholar] [CrossRef]
- Pierce, S. The jeweled armor of Tillandsia—Multifaceted or elongated trichomes provide photoprotection. Aliso A J. Syst. Florist. Bot. 2007, 23, 44–52. [Google Scholar] [CrossRef]
- Agudelo, C.M.; Benavides, A.M.; Taylor, T.; Feeley, K.J.; Duque, A. Functional composition of epiphyte communities in the Colombian Andes. Ecology 2019, 100, e02858. [Google Scholar] [CrossRef]
- De Paula Oliveira, R.; Zotz, G.; Wanek, W.; Franco, A.C. Leaf trait co-variation and trade-offs in gallery forest C3 and CAM epiphytes. Biotropica 2021, 53, 520–535. [Google Scholar] [CrossRef]
- Guzmán-Jacob, V.; Guerrero-Ramírez, N.R.; Craven, D.; Brant Paterno, G.; Taylor, A.; Krömer, T.; Wanek, W.; Zotz, G.; Kreft, H. Broad-and small-scale environmental gradients drive variation in chemical, but not morphological, leaf traits of vascular epiphytes. Funct. Ecol. 2022, 36, 1858–1872. [Google Scholar] [CrossRef]
- Richards, J.H.; Damschen, E.I. Leaf economics in a three-dimensional environment: Testing leaf trait responses in vascular epiphytes to land use, climate and tree zone. Funct. Ecol. 2022, 36, 727–738. [Google Scholar] [CrossRef]
- Wagner, K.; Wanek, W.; Zotz, G. Functional traits of a rainforest vascular epiphyte community: Trait covariation and indications for host specificity. Diversity 2021, 13, 97. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.; Diemer, M. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations (FAO). Global Ecological Zones for Fao Forest Reporting: 2010 Update; Forest Resources Assessment Working Paper 179; FAO: Rome, Italy, 2012. [Google Scholar]
- Rodríguez-Robles, U.; Arredondo, T. The role of the geologic substrate on Tillandsia recurvata infestation and the development of forest decaying on a semiarid oak forest. Catena 2022, 208, 105724. [Google Scholar] [CrossRef]
- Westbeld, A.; Klemm, O.; Grießbaum, F.; Sträter, E.; Larrain, H.; Osses, P.; Cereceda, P. Fog deposition to a Tillandsia carpet in the Atacama Desert. Ann. Geophys. 2009, 27, 3571–3576. [Google Scholar] [CrossRef] [Green Version]
- Barfuss, M.H.; Till, W.; LEME, E.M.; Pinzón, J.P.; Manzanares, J.M.; Halbritter, H.; Brown, G.K. Taxonomic revision of Bromeliaceae subfam. Tillandsioideae based on a multi-locus DNA sequence phylogeny and morphology. Phytotaxa 2016, 279, 1–97. [Google Scholar] [CrossRef]
- Donadío, S.; Till, W.; Pozner, R.; Barfuss, M.H.; Giussani, L.M.; Starr, J.R. Phylogenetic relationships and evolutionary trends in Tillandsia subgenus Diaphoranthema and xerophytic species of subgenus Phytarrhiza (Bromeliaceae: Tillandsioideae). Bot. J. Linn. Soc. 2022, 200, 1–22. [Google Scholar] [CrossRef]
- Terreros-Olivares, L. Revisión Sistemática del Complejo de Especies de Tillandsia juncea (Ruiz & Pav.) Poir. Bromeliaceae. Master’s Thesis, Universidad Autónoma Metropolitana, México DF, México, 2012. [Google Scholar]
- Borthagaray, A.I.; Fuentes, M.A.; Marquet, P.A. Vegetation pattern formation in a fog-dependent ecosystem. J. Theor. Biol. 2010, 265, 18–26. [Google Scholar] [CrossRef]
- Benzing, D.H. Vascular Epiphytes; Cambridge University Press: Cambridge, UK, 1990. [Google Scholar]
- Zotz, G. Vascular epiphytes in the temperate zones–A review. Plant Ecol. 2005, 176, 173–183. [Google Scholar] [CrossRef] [Green Version]
- Reyes-García, C.; Mejia-Chang, M.; Jones, G.D.; Griffiths, H. Water vapour isotopic exchange by epiphytic bromeliads in tropical dry forests reflects niche differentiation and climatic signals. Plant Cell Environ. 2008, 31, 828–841. [Google Scholar] [CrossRef]
- Dejean, A.; Olmsted, I.; Snelling, R.R. Tree-epiphyte-ant relationships in the low inundated forest of Sian Ka’an Biosphere Reserve, Quintana Roo, Mexico. Biotropica 1995, 27, 57–70. [Google Scholar] [CrossRef]
- Crayn, D.M.; Winter, K.; Schulte, K.; Smith, J.A.C. Photosynthetic pathways in Bromeliaceae: Phylogenetic and ecological significance of CAM and C3 based on carbon isotope ratios for 1893 species. Bot. J. Linn. Soc. 2015, 178, 169–221. [Google Scholar] [CrossRef] [Green Version]
- Gentry, A.H.; Dodson, C. Contribution of nontrees to species richness of a tropical rain forest. Biotropica 1987, 19, 149–156. [Google Scholar] [CrossRef]
- Nieder, J.; Prosperí, J.; Michaloud, G. Epiphytes and their contribution to canopy diversity. In Tropical Forest Canopies: Ecology and Management; Springer: Dordrecht, The Netherlands, 2001; pp. 51–63. [Google Scholar]
- Steege, H.T.; Cornelissen, J.H.C. Distribution and ecology of vascular epiphytes in lowland rain forest of Guyana. Biotropica 1989, 21, 331–339. [Google Scholar] [CrossRef]
- Krömer, T.; Kessler, M.; Gradstein, S.R. Vertical stratification of vascular epiphytes in submontane and montane forest of the Bolivian Andes: The importance of the understory. Plant Ecol. 2007, 189, 261–278. [Google Scholar] [CrossRef]
- Chilpa-Galván, N.; Tamayo-Chim, M.; Andrade, J.L.; Reyes-García, C. Water table depth may influence the asymmetric arrangement of epiphytic bromeliads in a tropical dry forest. Plant Ecol. 2013, 214, 1037–1048. [Google Scholar] [CrossRef]
- Zotz, G.; Leja, M.; Aguilar-Cruz, Y.; Einzmann, H.J. How much water is in the tank? An allometric analysis with 205 bromeliad species. Flora 2020, 264, 151557. [Google Scholar] [CrossRef]
- Belluau, M.; Shipley, B. Linking hard and soft traits: Physiology, morphology and anatomy interact to determine habitat affinities to soil water availability in herbaceous dicots. PLoS ONE 2018, 13, e0193130. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Harguindeguy, N.; Díaz, S.; Garnier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; Bret-Harte, M.S.; Cornwell, W.K.; Craine, J.M.; Gurvich, D.E.; et al. New handbook for standardised measurement of plant functional traits worldwide. Aust. J. Bot. 2013, 61, 167–234. [Google Scholar] [CrossRef]
- Xiong, D.; Chen, J.; Yu, T.; Gao, W.; Ling, X.; Li, Y.; Peng, S.; Huang, J. SPAD-based leaf nitrogen estimation is impacted by environmental factors and crop leaf characteristics. Sci. Rep. 2015, 5, 1–12. [Google Scholar] [CrossRef] [Green Version]
- WFO. World Flora Online. Available online: http://www.worldfloraonline.org/ (accessed on 21 July 2022).
- Bromeliad.org.au. Tillandsia. Available online: http://www.bromeliad.org.au/pictures/Tillandsia.htm (accessed on 24 July 2022).
- Chileflora.com. Tillandsia landbeckii Phil. Available online: http://www.chileflora.com/Florachilena/FloraSpanish/LowResPages/SH1153.htm (accessed on 26 July 2022).
- Abramoff, M.D.; Magalhaes, P.J.; Ram, S.J. Imagen processing with Image. J. Biophotonics Int. 2004, 11, 36–42. [Google Scholar]
- Tropicos.org. Missouri Botanical Garden. Available online: https://tropicos.org (accessed on 25 July 2022).
- POWO. Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. Available online: http://plantsoftheworldonline.org (accessed on 21 July 2022).
- Biokic. Biodiversity Knowledge Integration Center from the University of Arizona. Available online: https://biokic.asu.edu/ (accessed on 26 July 2022).
- Red de Herbarios del Noroeste de México. Bromeliaceae. Available online: https://herbanwmex.net/portal/taxa/index.php?tid=5442&clid=0&pid=0&taxauthid=1 (accessed on 26 July 2022).
- DGRU. Portal de Datos Abiertos UNAM, Colecciones Universitarias. Available online: https://datosabiertos.unam.mx/ (accessed on 26 July 2022).
- Matos, J.A.; Rudolph, D.C. Aspects of the Life History of Tillandsia Deppeana. In World Bromeliad Conference; Gardner, C.S., Ed.; Mission Press: Corpus Christi, TX, USA, 1984; pp. 71–75. [Google Scholar]
- Cogliatti-Carvalho, L.; Rocha-Pessôa, T.C.; Nunes-Freitas, A.F.; Duarte Rocha, C.F. Water volume stored in bromeliad tanks in Brazilian restinga hábitats. Acta Bot. Bras. 2010, 24, 84–95. [Google Scholar] [CrossRef]
- Dézerald, O.; Talaga, S.; Leroy, C.; Carrias, J.F.; Corbara, B.; Dejean, A.; Céréghino, R. Environmental determinants of macroinvertebrate diversity in small water bodies: Insights from tank-bromeliads. Hydrobiologia 2014, 723, 77–86. [Google Scholar] [CrossRef] [Green Version]
- Nuzhyna, N.V.; Kolomiyets, T.V. Differences in anatomical and morphological structure of plants from the genus Neoregelia LB Sm. (Bromeliaceae) as a result of adaptation to arid environmental conditions. Mod. Phytomorphol. 2015, 8, 137–144. [Google Scholar]
- Zotz, G.; Andrade, J.L. Water relations of two co-occurring epiphytic bromeliads. J. Plant Physiol. 1998, 152, 545–554. [Google Scholar] [CrossRef]
- Johnson, T.F.; Isaac, N.J.; Paviolo, A.; González-Suárez, M. Handling missing values in trait data. Glob. Ecol. Biogeogr. 2021, 30, 51–62. [Google Scholar] [CrossRef]
- Jin, Y.; Qian, H.V. PhyloMaker: An R package that can generate very large phylogenies for vascular plants. Ecography 2019, 42, 1353–1359. [Google Scholar] [CrossRef] [Green Version]
- GBIF. Database Global Biodiversity Information Facility. Available online: https://www.gbif.org/ (accessed on 19 July 2022).
- Abatzoglou, J.T.; Dobrowski, S.Z.; Parks, S.A.; Hegewisch, K.C. TerraClimate, a high-resolution global dataset of monthly climate and climatic water balance from 1958–2015. Sci. Data 2018, 5, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Zomer, R.J.; Xu, J.; Trabucco, A. Version 3 of the global aridity index and potential evapotranspiration database. Sci. Data 2022, 9, 1–15. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- QGIS.org, QGIS Geographic Information System. QGIS Association. 2018. Available online: http://www.qgis.org (accessed on 3 August 2022).
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Bolar, K. STAT: Interactive Document for Working with Basic Statistical Analysis; R package Version 0.1.0; Publisher: Manipal, India, 2019. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Aoyama, E.M.; Sajo, M. Estructura foliar de Aechmea Ruiz & Pav. subgênero Lamprococcus (Beer) Baker e espécies relacionadas (Bromeliaceae). Braz. J. Bot. 2003, 26, 461–473. [Google Scholar]
- Belmonte, E.; Arriaza, B.; Arismendi, M.; Sepúlveda, G. Foliar Anatomy of Three Native Species of Tillandsia L. from the Atacama Desert, Chile. Plants 2022, 11, 870. [Google Scholar] [CrossRef]
- Benzing, D.H.; Renfrow, A. The mineral nutrition of Bromeliaceae. Bot. Gaz. 1974, 135, 281–288. [Google Scholar] [CrossRef]
- Benzing, D.H.; Friedman, W.E. Patterns of foliar pigmentation in Bromeliaceae and their adaptive significance. Selbyana 1981, 5, 224–240. [Google Scholar]
- Benzing, D.H.; Davidson, E.A. Oligotrophic Tillandsia circinnata Schlecht (Bromeliaceae): An assessment of its patterns of mineral allocation and reproduction. Am. J. Bot. 1979, 66, 386–397. [Google Scholar] [CrossRef]
- Braga, M.M.N. Anatomia foliar de Bromeliaceae da Campina. Acta Amazon. 1977, 7, 5–74. [Google Scholar]
- Brighigna, L.; Cecchi Fiordi, A.; Palandri, M.R. Structural characteristics of the mesophyll in some Tillandsia species. Phytomorphology 1984, 34, 191–200. [Google Scholar]
- Caballero-Rueda, L.M.; Rodríguez, N.; Martin, C. Dinámica de elementos en epifitos de un bosque altoandino de la Cordillera Oriental de Colombia. Caldasia 1997, 19, 311–322. [Google Scholar]
- Cach-Pérez, M.J.; Andrade, J.L.; Cetzal-Ix, W.; Reyes-García, C. Environmental influence on the inter-and intraspecific variation in the density and morphology of stomata and trichomes of epiphytic bromeliads of the Yucatan Peninsula. Bot. J. Linn. Soc. 2016, 181, 441–458. [Google Scholar] [CrossRef] [Green Version]
- Cardelús, C.L.; Mack, M. The nutrient status of epiphytes and their host trees along an elevational gradient in Costa Rica. Plant Ecol. 2010, 207, 25–37. [Google Scholar] [CrossRef]
- Casañas, O.L.; Jáuregui, D. Morfoanatomía foliar de epífitas presentes en un bosque nublado, Altos de Pipe, Estado Miranda, Venezuela. Acta Bot. Venez. 2011, 34, 153–176. [Google Scholar]
- Leroy, C.; Gril, E.; Si Ouali, L.; Coste, S.; Gérard, B.; Maillard, P.; Mercier, H.; Stahl, C. Water and nutrient uptake capacity of leaf-absorbing trichomes vs. roots in epiphytic tank bromeliads. Environ. Exp. Bot. 2019, 163, 112–123. [Google Scholar] [CrossRef]
- Crayn, D.M.; Winter, K.; Smith, J.A.C. Multiple origins of crassulacean acid metabolism and the epiphytic habit in the Neotropical family Bromeliaceae. Proc. Natl. Acad. Sci. USA 2004, 101, 3703–3708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Oliveira, M.L.; de Melo, E.J.; Miguens, F.C. Tillandsia stricta Sol (Bromeliaceae) leaves as monitors of airborne particulate matter—A comparative SEM methods evaluation: Unveiling an accurate and odd HP-SEM method. Microsc. Res. Tech. 2016, 79, 869–879. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, R.S.; de Oliveira Souza, S.; Aona, L.Y.S.; Souza, F.V.D.; Rossi, M.L.; de Souza, E.H. Leaf structure of Tillandsia species (Tillandsioideae: Bromeliaceae) by light microscopy and scanning electron microscopy. Microsc. Res. Tech. 2022, 85, 253–269. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Álvarez, E.A.; de la Barrera, E. Drying protocol does not alter plant δ13C and δ15N: A baseline survey for ecological studies. Isotopes Environ. Health Stud. 2019, 55, 526–531. [Google Scholar] [CrossRef] [PubMed]
- Einzmann, H.J.R.; Beyschlag, J.; Hofhansl, F.; Wanek, W.; Zotz, G. Host tree phenology affects vascular epiphytes at the physiological, demographic and community level. AoB Plants 2015, 7, plu073. [Google Scholar] [CrossRef] [Green Version]
- Felix, J.D.; Avery, G.B.; Mead, R.N.; Kieber, R.J.; Willey, J.D. Nitrogen content and isotopic composition of Spanish Moss (Tillandsia usneoides L.): Reactive nitrogen variations and source implications across an urban coastal air shed. Environ. Process. 2016, 3, 711–722. [Google Scholar] [CrossRef]
- Gómez, M.A.; Winkler, S. Bromelias en manglares del Pacífico de Guatemala. Rev. Biol. Trop. 1991, 39, 207–214. [Google Scholar]
- Griffiths, H. Carbon balance during CAM: An assessment of respiratory CO22 recycling in the epiphytic bromeliads Aechmea nudicaulis and Aechmea fendleri. Plant Cell Environ. 1988, 11, 603–611. [Google Scholar] [CrossRef]
- Griffiths, H.; Lüttge, U.; Stimmel, K.H.; Crook, C.E.; Griffiths, N.M.; Smith, J.A.C. Comparative ecophysiology of CAM and C3 bromeliads. III. Environmental influences on CO2 assimilation and transpiration. Plant Cell Environ. 1986, 9, 385–393. [Google Scholar] [CrossRef]
- Griffiths, H.; Smith, J.A.C.; Lüttge, U.; Popp, M.; Cram, W.J.; Diaz, M.A.; Lee, H.S.L.; Medina, E.; Schäfer, C.; Stimmel, K.H. Ecophysiology of xerophytic and halophytic vegetation of a coastal alluvial plain in northern Venezuela. IV. Tillandsia flexu-osa Sw. and Schomburgkia humboldtiana Reichb., epiphytic CAM plants. New Phytol. 1989, 111, 273–282. [Google Scholar] [CrossRef]
- Hietz, P.; Wanek, W.; Popp, M. Stable isotopic composition of carbon and nitrogen and nitrogen content in vascular epiphytes along an altitudinal transect. Plant Cell Environ. 1999, 22, 1435–1443. [Google Scholar] [CrossRef]
- Hietz, P.; Wanek, W. Size-dependent variation of carbon and nitrogen isotope abundances in epiphytic bromeliads. Plant Biol. 2003, 5, 137–142. [Google Scholar] [CrossRef]
- Husk, G.J.; Weishampel, J.E.; Schlesinger, W.H. Mineral dynamics in spanish moss, Tillandsia usneoides L. (Bromeliaceae), from Central Florida, USA. Sci. Total Environ. 2004, 321, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Adams, W.W.; Martin, C.E. Morphological changes accompanying the transition from juvenile (atmospheric) to adult (tank) forms in the Mexican epiphyte Tillandsia deppeana (Bromeliaceae). Am. J. Bot. 1986, 73, 1207–1214. [Google Scholar]
- Isley, P.T. Tillandsia: The World’s Most Unusual Air Plants, 1st ed.; Botanical Press: San Diego, CA, USA, 1987; p. 256. [Google Scholar]
- Latorre, C.; González, A.L.; Quade, J.; Fariña, J.M.; Pinto, R.; Marquet, P.A. Establishment and formation of fog-dependent Tillandsia landbeckii dunes in the Atacama Desert: Evidence from radiocarbon and stable isotopes. J. Geophys. Res. Biogeosci. 2011, 116, 1–12. [Google Scholar] [CrossRef]
- Leroy, C.; Carrias, J.F.; Céréghino, R.; Corbara, B. The contribution of microorganisms and metazoans to mineral nutrition in bromeliads. J. Plant Ecol. 2016, 9, 241–255. [Google Scholar] [CrossRef] [Green Version]
- Leroy, C.; Carrias, J.F.; Corbara, B.; Pélozuelo, L.; Dézerald, O.; Brouard, O.; Céréghino, R. Mutualistic ants contribute to tank-bromeliad nutrition. Ann. Bot. 2013, 112, 919–926. [Google Scholar] [CrossRef] [Green Version]
- Leroy, C.; Corbara, B.; Dejean, A.; Céréghino, R. Ants mediate foliar structure and nitrogen acquisition in a tank-bromeliad. New Phytol. 2009, 183, 1124–1133. [Google Scholar] [CrossRef]
- López-Ferrari, A.R.; Espejo-Serna, A.; Blanco, P. Circunscripción de Tillandsia chaetophylla Mez y descripción de Tillandsia sessemocinoi (Bromeliaceae: Tillandsioideae). Acta Bot. Mex. 2006, 76, 77–88. [Google Scholar] [CrossRef] [Green Version]
- Males, J.; Griffiths, H. Economic and hydraulic divergences underpin ecological differentiation in the Bromeliaceae. Plant Cell Environ. 2018, 41, 64–78. [Google Scholar] [CrossRef] [Green Version]
- Males, J. Structure-Function Relationships in the Water-Use Strategies and Ecological Diversity of the Bromeliaceae. Ph.D. Thesis, University of Cambridge, Cambridge, UK, 2017. [Google Scholar]
- Marino, N.A.; Guariento, R.D.; Dib, V.; Azevedo, F.D.; Farjalla, V.F. Habitat size determine algae biomass in tank-bromeliads. Hydrobiologia 2011, 678, 191–199. [Google Scholar] [CrossRef]
- Martin, C.E. Physiological Ecology of the Bromeliaceae. Bot. Rev. 1994, 60, 1–82. [Google Scholar] [CrossRef]
- Martins, J.P.R.; Martins, A.D.; Pires, M.F.; Braga Junior, R.A.; Reis, R.O.; Dias, G.D.M.G.; Pasqual, M. Anatomical and physiological responses of Billbergia zebrina (Bromeliaceae) to copper excess in a controlled microenvironment. Plant Cell Tissue Organ Cult. 2016, 126, 43–57. [Google Scholar] [CrossRef]
- Maxwell, K.; Griffiths, H.; Borland, A.M.; Young, A.J.; Broadmeadow, M.S.J.; Fordham, M.C. Short-term photosynthetic responses of the C3-CAM epiphyte Guzmania monostachia var. monostachia to tropical seasonal transitions under field conditions. Aust. J. Plant Physiol. 1995, 22, 771–781. [Google Scholar] [CrossRef]
- Meisner, K. Functional Relevance of Heteroblasty in Bromeliaceae. Ph.D. Thesis, Carl von Ossietzky Universität, Oldenburg, Germany, 2012. [Google Scholar]
- Palací, C.A.; Brown, G.K.; Tuthill, D.E. Vegetative Morphology and Leaf Anatomy of Catopsis (Tillandsioideae: Bromeliaceae). Selbyana 2004, 25, 138–150. [Google Scholar]
- Pereira-Zaldívar, N.A.; Patiño-López, L.D.; Rodríguez-García, R.; Andrade, J.L.; Cach-Pérez, M.J.; Espadas-Manrique, C.; Barredo-Pool, F.; Reyes-García, C. Life form and anatomical traits related to trichome mediated external water transport in epiphytic Bromeliaceae. Plant Cell Environ. 2022. Submitted. [Google Scholar]
- Pereira, T.A.R.; de Oliveira, T.S.; da Silva, L.C.; Azevedo, A.A. Comparative leaf anatomy of four species of Bromelioideae (Bromeliaceae) occurring in the Atlantic Forest, Brazil. Botany 2011, 89, 243–253. [Google Scholar] [CrossRef]
- Petter, G.; Wagner, K.; Wanek, W.; Sánchez Delgado, E.J.; Zotz, G.; Cabral, J.S.; Kreft, H. Functional leaf traits of vascular epiphytes: Vertical trends within the forest, intra- and interspecific trait variability, and taxonomic signals. Func. Ecol. 2016, 30, 188–198. [Google Scholar] [CrossRef] [Green Version]
- Pierce, S.; Winter, K.; Griffiths, H. Carbon isotope ratio and the extent of daily CAM use by Bromeliaceae. New Phytol. 2002, 156, 75–83. [Google Scholar] [CrossRef]
- Proença, S.L.; Sajo, M.D.G. Anatomia foliar de bromélias ocorrentes em áreas de cerrado do Estado de São Paulo, Brasil. Acta Bot. Brasil. 2007, 21, 657–673. [Google Scholar] [CrossRef]
- Puccio, P. Tillandsia tenuifolia L. Monaco nature Encyclopedia. Available online: https://www.monaconatureencyclopedia.com/tillandsia-tenuifolia/?lang=es (accessed on 19 July 2022).
- Rammler, H. Ökophysiologie von Epiphyten und Bäumen in einem Bergregenwald in Mexiko und einem Tieflandregenwald in Costa Rica. Master Thesis, Universität für Bodenkultur, Wien, Germany, 2004. [Google Scholar]
- Reyes-García, C. Ecofisiología de epifitas de selva baja caducifolia del género Tillandsia (Bromeliaceae): Estacionalidad y fotosíntesis. Bachelor’s Thesis, Universidad nacional Autónoma de México, Mexico City, Mexico, 2001. [Google Scholar]
- Ribeiro, F.; Aoyama, E. Anatomia foliar de Bromeliaceae da trilha do rio Timbuí na Estação Biológica de Santa Lúcia, Santa Teresa–es. Enciclopédia Biosfera 2019, 16, 185–197. [Google Scholar] [CrossRef]
- Rosado-Calderón, A.T.; Tamayo-Chim, M.; de la Barrera, E.; Ramírez-Morillo, I.M.; Andrade, J.L.; Briones, O.; Reyes-García, C. High resilience to extreme climatic changes in the CAM epiphyte Tillandsia utriculata L. (Bromeliaceae). Physiol. Plant. 2020, 168, 547–562. [Google Scholar] [CrossRef]
- Schmidt, G.; Stuntz, S.; Zotz, G. Plant size—An ignored parameter in epiphyte ecophysiology. Plant. Ecol. 2001, 153, 65–72. [Google Scholar] [CrossRef]
- Silva, E.C. Aechmea bromeliifolia (Rudge) Baker (Bromeliaceae) Cultivada In Vitro e Ex Vitro: Morfologia, Anatomia e Ultraestrutura. Master Thesis, Universidade Federal de Goiás, Goiânia, Brazil, 2016. [Google Scholar]
- Smith, J.A.C.; Griffiths, H.; Lüttge, U.; Crook, C.E.; Griffiths, N.M.; Stimmel, K.H. Comparative ecophysiology of CAM and C3 bromeliads. IV. Plant water relations. Plant Cell Environ. 1986, 9, 395–410. [Google Scholar] [CrossRef]
- Souza, G.M.D.; Estelita, M.E.M.; Wanderley, M.D.G.L. Anatomia foliar de espécies brasileiras de Aechmea subg. Chevaliera (Gaudich. ex Beer) Baker, Bromelioideae-Bromeliaceae. Braz. J. Bot. 2005, 28, 603–613. [Google Scholar] [CrossRef] [Green Version]
- Stuntz, S.; Zotz, G. Photosynthesis in vascular epiphytes—A survey of 27 species of diverse taxonomic origin. Flora 2001, 196, 132–141. [Google Scholar] [CrossRef]
- Susan-Tepetlan, T.M.; Velázquez-Rosas, N.; Krömer, T. Cambios en las características funcionales de epífitas vasculares de bosque mesófilo de montaña y vegetación secundaria en la región central de Veracruz, México. Bot. Sci. 2015, 93, 1–11. [Google Scholar] [CrossRef]
- Versieux, L.M.; Medeiros, A.S.M. Leaf anatomical characterization of Guzmania Ruiz & Pav. and Mezobromelia LB Sm. (Tillandsioideae, Bromeliaceae). J. Bromeliad Soc. 2017, 67, 8–26. [Google Scholar]
- Vite-Posadas, J.A.; Brechú-Franco, A.E.; Laguna-Hernández, G.; Rojas-Bribiesca, M.G.; Osuna-Fernández, H.R. Morphoanatomical characterization and antimicrobial activity of Tillandsia imperialis (Bromeliaceae). Polibotánica 2011, 31, 20–29. [Google Scholar]
- Wanek, W.; Zotz, G. Are vascular epiphytes nitrogen or phosphorus limited? A study of plant 15N fractionation and foliar N: P stoichiometry with the tank bromeliad Vriesea sanguinolenta. New Phytol. 2011, 192, 462–470. [Google Scholar] [CrossRef]
- Wania, R.; Hietz, P.; Wanek, W. Natural 15N abundance of epiphytes depends on the position within the forest canopy: Source signals and isotope fractionation. Plant Cell Environ. 2002, 25, 581–589. [Google Scholar] [CrossRef]
- Wester, S.; Mendieta-Leiva, G.; Nauheimer, L.; Wanek, W.; Kreft, H.; Zotz, G. Physiological diversity and biogeography of vascular epiphytes at Río Changuinola. Panama Flora 2011, 206, 66–79. [Google Scholar] [CrossRef]
- Zambrano, A.R.C.; Linis, V.C.; Nepacina, M.R.J.; Silvestre, M.L.T.; Foronda, J.R.F.; Janairo, J.I.B. Wetting properties and foliar water uptake of Tillandsia L. Biotribology 2019, 19, 100103. [Google Scholar] [CrossRef]
- Zorger, B.B.; Arrivabene, H.P.; Milanez, C.R.D. Adaptive morphoanatomy and ecophysiology of Billbergia euphemiae, a hemiepiphyte Bromeliaceae. Rodriguésia 2019, 70, 1–10. [Google Scholar] [CrossRef]
- Zotz, G. How prevalent is crassulacean acid metabolism among vascular epiphytes? Oecologia 2004, 138, 184–192. [Google Scholar] [CrossRef] [Green Version]
- Zotz, G.; Asshoff, R. Growth in epiphytic bromeliads: Response to the relative supply of phosphorus and nitrogen. Plant Biol. 2010, 12, 108–113. [Google Scholar] [CrossRef]
- Zotz, G.; Ziegler, H. The occurrence of crassulacean acid metabolism among vascular epiphytes from Central Panama. New Phytol. 1997, 137, 223–229. [Google Scholar] [CrossRef]
- Zotz, G.; Enslin, A.; Hartung, W.; Ziegler, H. Physiological and anatomical changes during the early ontogeny of the heteroblastic bromeliad, Vriesea sanguinolenta, do not concur with the morphological change from atmospheric to tank form. Plant Cell Environ. 2004, 27, 1341–1350. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Unit | Abbreviation | Median | Range | Records | Species | Genera |
|---|---|---|---|---|---|---|---|
| Adult plant height * | m | Height | 0.4 | (0.03–2.7) | 507 | 126 | 16 |
| Force to punch | N mm−1 | FP | 1.4 | (0.17–6.2) | 706 | 36 | 7 |
| Leaf area * | cm2 | LA | 8678 | (29–1033) | 1702 | 201 | 23 |
| Leaf carbon content per dry mass | % | C | 45.2 | (27.9–69.3) | 502 | 59 | 11 |
| Leaf carbon isotope signature * | ‰ | δ13C | −15.9 | (−35.08–11) | 903 | 197 | 23 |
| Leaf chlorophyll content per dry mass | µg g−1 | LCh | 2800 | (200–12200) | 43 | 35 | 6 |
| Leaf dry matter content | mg g | LD | 161.2 | (37.3–520) | 1440 | 49 | 8 |
| Leaf index * | LI | 13.4 | (2.2–250) | 212 | 202 | 22 | |
| Leaf length * | cm | LL | 38.7 | (1.8–161) | 539 | 204 | 23 |
| Leaf nitrogen isotope signature * | ‰ | δ15N | −4.7 | (−15.2–7.5) | 643 | 69 | 12 |
| Leaf nitrogen content per dry mass * | mg g | N | 7.4 | (1–25.5) | 554 | 71 | 12 |
| Leaf phosphorus content per dry mass | mg g | P | 0.6 | (0.02–5) | 185 | 61 | 11 |
| Leaf thickness * | mm | LT | 0.6 | (0.07–4.5) | 1751 | 97 | 17 |
| Leaf trichome density * | mm−2 | TD | 35.2 | (2.8–120.9) | 1104 | 74 | 11 |
| Leaf water content on area basis * | g m2 | LWA | 540.8 | (91.7–6017) | 936 | 92 | 16 |
| Leaf width * | cm | LW | 3.0 | (0.05–17.5) | 449 | 202 | 22 |
| Light saturated photosynthetic rate per leaf area | µmol m2 s | Amax | 2.3 | (0.7–4.7) | 42 | 36 | 9 |
| Specific leaf area * | mm2 mg−1 | SLA | 8.9 | (0.01–68.5) | 1709 | 109 | 16 |
| Stomatal density * | mm−2 | SD | 21.0 | (2.8–88.8) | 975 | 102 | 14 |
| Stomatal length | µm | SL | 39.8 | (14.4–284.3) | 257 | 45 | 8 |
| Stomatal width | µm | SW | 35.0 | (9.7–338.5) | 56 | 32 | 7 |
| Tank capacity * | ml | TC | 85.7 | (0–4924) | 213 | 190 | 23 |
| Total leaf water content * | g | LWC | 3.1 | (0.001–147) | 738 | 92 | 16 |
| Pseudobulb presence * | PB | 204 | 204 | 23 | |||
| Photosynthetic pathway | 204 | 204 | 23 | ||||
| TOTAL | 16,574 | 204 | 23 |
| Genera | Records | Species | Traits |
|---|---|---|---|
| Aechmea Mez | 1549 | 36 | 25 |
| Araeococcus Brongn. | 43 | 2 | 18 |
| Billbergia Thunb. | 280 | 8 | 22 |
| Canistropsis (Mez) Leme | 52 | 1 | 10 |
| Canistrum E.Morren | 18 | 2 | 8 |
| Catopsis Griseb. | 1138 | 7 | 25 |
| Fascicularia Mez | 78 | 1 | 11 |
| Goudaea W.Till & Barfuss | 15 | 1 | 10 |
| Guzmania Ruiz & Pav. | 2069 | 31 | 25 |
| Josemania W.Till & Barfuss | 13 | 1 | 12 |
| Lemeltonia Barfuss & W.Till | 206 | 1 | 21 |
| Lutheria Barfuss & W.Till | 21 | 1 | 18 |
| Lymania Read | 10 | 1 | 9 |
| Mezobromelia L. B. Sm. | 9 | 1 | 9 |
| Neoregelia L. B. Sm. | 63 | 5 | 14 |
| Nidularium Lem. | 92 | 8 | 19 |
| Quesnelia Gaudich. | 21 | 2 | 11 |
| Racinaea M.A.Spencer & L.B.Sm. | 878 | 5 | 22 |
| Ronnbergia E.Morren & Andre | 18 | 2 | 8 |
| Tillandsia L. | 9522 | 81 | 25 |
| Vriesea Lindl. | 93 | 5 | 14 |
| Wallisia E.Morr. | 411 | 1 | 24 |
| Werauhia J.R.Grant. | 75 | 6 | 19 |
| Functional Group | Description | Main Water Source | C3 or CAM | Genera | Previous Classification |
|---|---|---|---|---|---|
| Nebulophytes | Acicular leaves, usually with high leaf index, no tank capacity | Fog | Mostly CAM | Tillandsia, Araeococcus | (P) Type IV Atmosphere-Absorbing trichome (B) Type V |
| Pseudobulbs | Neotenic, forming pseudobulbs, highly succulent, thick leaves, no tank capacity | Rain/ internal reserves | CAM | Tillandsia | (P) Type IV Atmosphere-Absorbing trichome (B) Type V |
| Shallow tanks | 2–60 mL tank capacity, small sized, thin leaves | Dew/rain | Mostly CAM | Tillandsia, Araeococcus, Aechmea, Wallisia, Lemeltonia, Canistrum, Neoregelia, Nidularium, Billbergia, Quesnelia, Ronnbergia | (P) Type IV Atmosphere-Absorbing trichome (B) Type V |
| CAM tank | >61 mL tank capacity, large size | Rain | CAM | Aechmea, Tillandsia, Billbergia, Neoregelia, Nidularium, Canistropsis, Canistrum, Lymania, Quesnelia | (P) Type III Tank-Absorbing trichome (B)Type III |
| C3 tank | >5 mL tank capacity, low specific leaf area, medium to large size | Rain | C3 | Tillandsia, Catopsis, Guzmania, Billbergia, Lutheria, Werauhia, Racinaeae, Fascicularia, Goudaea, Josemania, Mesobromelia, Vriesea | (P) Type III Tank-Absorbing trichome (B)Type IV |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reyes-García, C.; Pereira-Zaldívar, N.A.; Espadas-Manrique, C.; Tamayo-Chim, M.; Chilpa-Galván, N.; Cach-Pérez, M.J.; Ramírez-Medina, M.; Benavides, A.M.; Hietz, P.; Zotz, G.; et al. New Proposal of Epiphytic Bromeliaceae Functional Groups to Include Nebulophytes and Shallow Tanks. Plants 2022, 11, 3151. https://doi.org/10.3390/plants11223151
Reyes-García C, Pereira-Zaldívar NA, Espadas-Manrique C, Tamayo-Chim M, Chilpa-Galván N, Cach-Pérez MJ, Ramírez-Medina M, Benavides AM, Hietz P, Zotz G, et al. New Proposal of Epiphytic Bromeliaceae Functional Groups to Include Nebulophytes and Shallow Tanks. Plants. 2022; 11(22):3151. https://doi.org/10.3390/plants11223151
Chicago/Turabian StyleReyes-García, Casandra, Narcy Anai Pereira-Zaldívar, Celene Espadas-Manrique, Manuela Tamayo-Chim, Nahlleli Chilpa-Galván, Manuel Jesús Cach-Pérez, Marypaz Ramírez-Medina, Ana Maria Benavides, Peter Hietz, Gerhard Zotz, and et al. 2022. "New Proposal of Epiphytic Bromeliaceae Functional Groups to Include Nebulophytes and Shallow Tanks" Plants 11, no. 22: 3151. https://doi.org/10.3390/plants11223151
APA StyleReyes-García, C., Pereira-Zaldívar, N. A., Espadas-Manrique, C., Tamayo-Chim, M., Chilpa-Galván, N., Cach-Pérez, M. J., Ramírez-Medina, M., Benavides, A. M., Hietz, P., Zotz, G., Andrade, J. L., Cardelús, C., de Paula Oliveira, R., Einzmann, H. J. R., Guzmán Jacob, V., Krömer, T., Pinzón, J. P., Sarmento Cabral, J., Wanek, W., & Woods, C. (2022). New Proposal of Epiphytic Bromeliaceae Functional Groups to Include Nebulophytes and Shallow Tanks. Plants, 11(22), 3151. https://doi.org/10.3390/plants11223151