
Phenylalanine Ammonia Lyase GmPAL1.1 Promotes Seed Vigor under High-Temperature and -Humidity Stress and Enhances Seed Germination under Salt and Drought Stress in Transgenic Arabidopsis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Cloning and Subcellular Localization of GmPAL1.1
2.2. Expression Patterns of GmPAL1.1 in Various Tissues and under HTH Stress
2.3. The Overexpression of GmPAL1.1 in Arabidopsis Reduced ROS Accumulation in Plants at Physiological Maturity Stage under HTH Stress
2.4. The Overexpression of GmPAL1.1 in Arabidopsis Enhanced Tolerance of Developing Seeds to HTH Stress and Produced High-Vigor Seeds
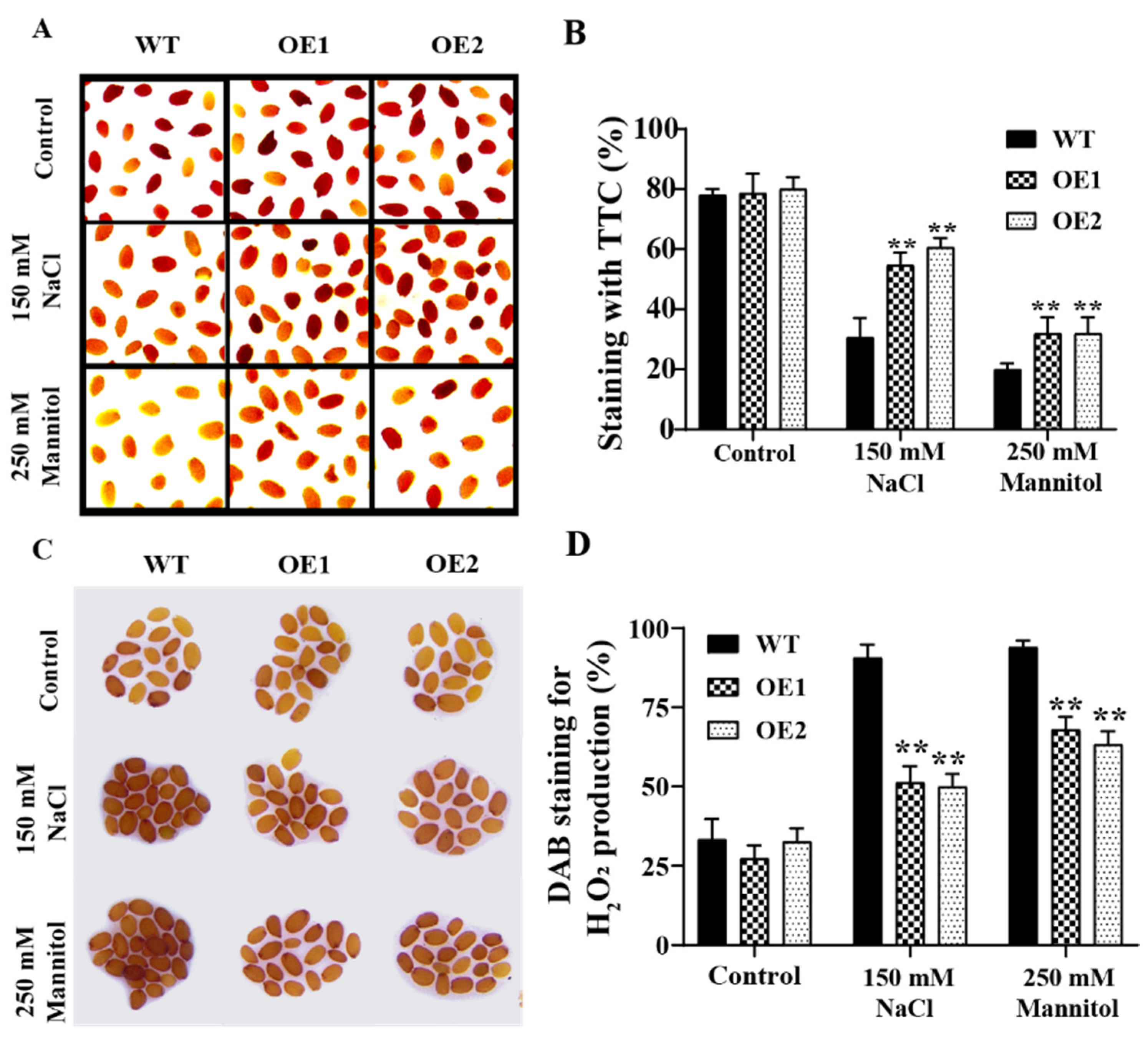
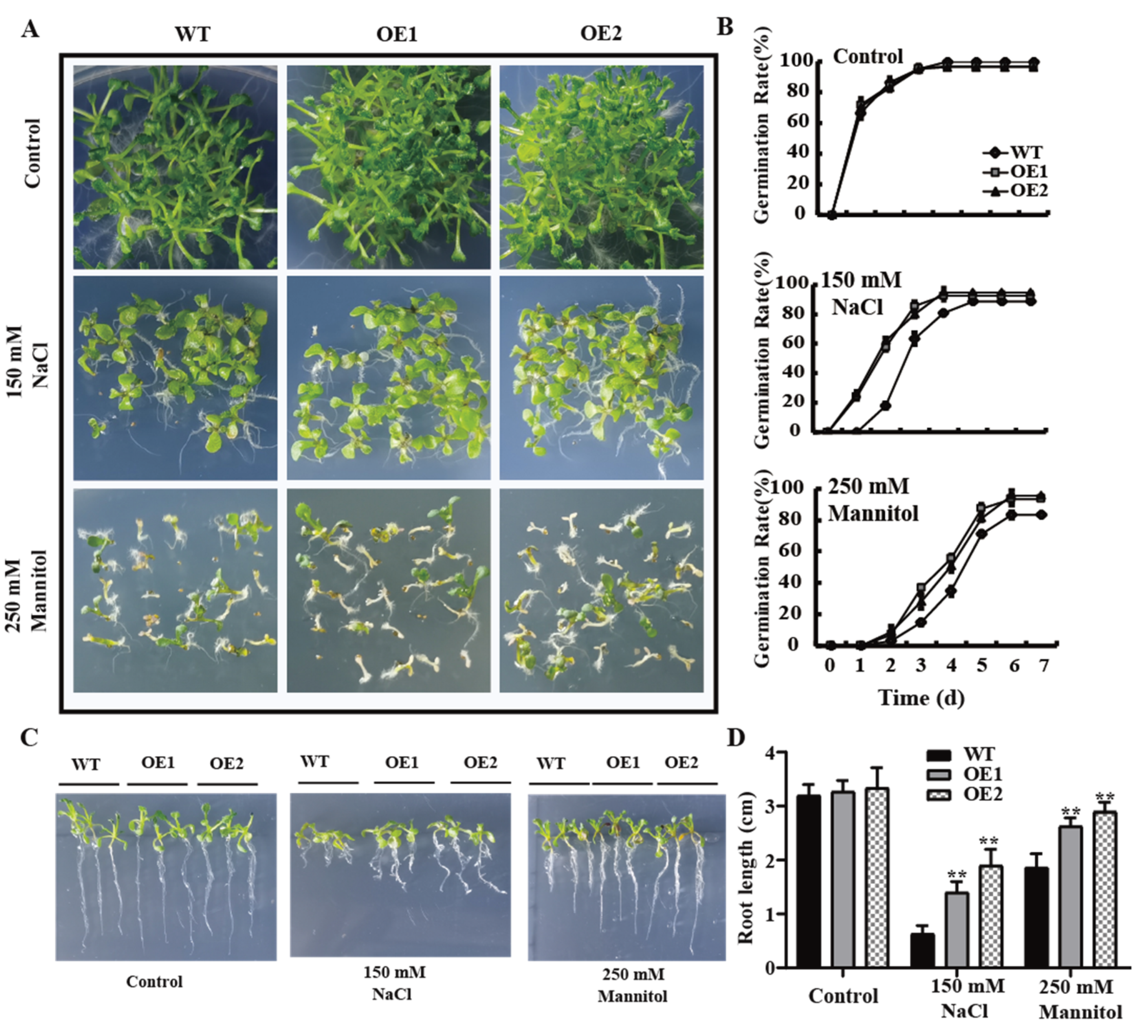
2.5. The Overexpression of GmPAL1.1 in Arabidopsis Improved Seed Vigor and Enhanced Tolerance to Salt and Drought Stresses during Germination
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Treatments
4.2. Cloning and Bioinformatics Analysis of GmPAL1.1
4.3. Subcellular Localization
4.4. Expression Analysis
4.5. Genetic Transformation and Screening of Transgenic Lines in Arabidopsis
4.6. TTC-Staining, DAB-Staining, and ROS Content Assays
4.7. Measurements of Flavonoids and Anthocyanins Contents
4.8. Measurements of SOD, POD, CAT, and MDA Contents
4.9. Statistics Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| ABA | Abscisic acid |
| BF | Bright field |
| CAT | Catalase |
| CDS | Coding sequence |
| DAB | 3,3′-diaminobenzidine |
| GFP | Green fluorescent protein |
| H2DCFDA | 2′,7′-Dichlorodihydrofluorescein diacetate |
| HTH | High temperature and humidity |
| MDA | Malondialdehyde |
| MS | Murashige-Skoog medium |
| NBT | Tetranitroblue tetrazolium chloride |
| OE | Over expression |
| ORF | Open Reading Frame |
| PCR | Polymerase Chain Reaction |
| PAL | Phenylalanine ammonia lyase |
| POD | Peroxidase |
| qRT-PCR | Quantitative real-time PCR |
| RH | Relative humidity |
| SOD | Superoxide dismutase |
| TTC | 2,3,5-triphenyl tetrazolium chloride |
| WT | Wild type |
| R5 | Beginning seed development |
| R7 | Physiological maturity |
References
- Olsen, K.M.; Lea, U.S.; Slimestad, R.; Verheul, M.; Lillo, C. Differential expression of four Arabidopsis PAL genes; PAL1 and PAL2 have functional specialization in abiotic environmental-triggered flavonoid synthesis. J. Plant Physiol. 2008, 165, 1491–1499. [Google Scholar] [CrossRef] [PubMed]
- Rohde, A.; Morreel, K.; Ralph, J.; Goeminne, G.; Hostyn, V.; De Rycke, R.; Kushnir, S.; Van Doorsselaere, J.; Joseleau, J.P.; Vuylsteke, M.; et al. Molecular phenotyping of the pal1 and pal2 mutants of Arabidopsis thaliana reveals far-reaching consequences on phenylpropanoid, amino acid, and carbohydrate metabolism. Plant Cell 2004, 16, 2749–2771. [Google Scholar] [CrossRef] [Green Version]
- Elkind, Y.; Edwards, R.; Mavandad, M.; Hedrick, S.A.; Ribak, O.; Dixon, R.A.; Lamb, C.J. Abnormal plant development and down-regulation of phenylpropanoid biosynthesis in transgenic tobacco containing a heterologous phenylalanine ammonia-lyase gene. Proc. Natl. Acad. Sci. USA 1990, 87, 9057–9061. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Gu, M.; Lai, Z.; Fan, B.; Shi, K.; Zhou, Y.; Yu, J.; Chen, Z. Functional analysis of the Arabidopsis PAL gene family in plant growth, development, and response to environmental stress. Plant Physiol. 2010, 153, 1526–1538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, V.; Wang, Z.; Wei, C.; Amo, A.; Ahmed, B.; Yang, X.; Zhang, X. Phenylpropanoid Pathway Engineering: An Emerging Approach towards Plant Defense. Pathogens 2020, 9, 312. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Liu, Y.; Yuan, D.; Duan, M.; Liu, Y.; Shen, Z.; Yang, C.; Qiu, Z.; Liu, D.; Wen, P.; et al. An R2R3 MYB transcription factor confers brown planthopper resistance by regulating the phenylalanine ammonia-lyase pathway in rice. Proc. Natl. Acad. Sci. USA 2020, 117, 271–277. [Google Scholar] [CrossRef]
- Van Eck, L.; Schultz, T.; Leach, J.E.; Scofield, S.R.; Peairs, F.B.; Botha, A.; Lapitan, N.L.V. Virus-induced gene silencing of WRKY53 and an inducible phenylalanine ammonia-lyase in wheat reduces aphid resistance. Plant Biotechnol. J. 2010, 8, 1023–1032. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, X.; Zhang, F.; Dong, L.; Wu, J.; Cheng, Q.; Qi, D.; Yan, X.; Jiang, L.; Fan, S.; et al. Phenylalanine ammonia-lyase2.1 contributes to the soybean response towards Phytophthora sojae infection. Sci. Rep. 2017, 7, 7242. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhang, X.; Liu, G.; Tang, Y.; Zhou, C.; Zhang, L.; Lv, J. The spike plays important roles in the drought tolerance as compared to the flag leaf through the phenylpropanoid pathway in wheat. Plant Physiol. Bioch. 2020, 152, 100–111. [Google Scholar] [CrossRef] [PubMed]
- Jiao, C.; Yang, R.; Zhou, Y.; Gu, Z. Nitric oxide mediates isoflavone accumulation and the antioxidant system enhancement in soybean sprouts. Food Chem. 2016, 204, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Reed, R.C.; Bradford, K.J.; Khanday, I. Seed germination and vigor: Ensuring crop sustainability in a changing climate. Heredity 2022, 128, 450–459. [Google Scholar] [CrossRef]
- Petla, B.P.; Kamble, N.U.; Kumar, M.; Verma, P.; Ghosh, S.; Singh, A.; Rao, V.; Salvi, P.; Kaur, H.; Saxena, S.C.; et al. Rice PROTEIN l-ISOASPARTYL METHYLTRANSFERASE isoforms differentially accumulate during seed maturation to restrict deleterious isoAsp and reactive oxygen species accumulation and are implicated in seed vigor and longevity. New Phytol. 2016, 211, 627–645. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, C.D.; Chen, J.; Clark, D.; Perez, H.; Huo, H.A. Effects of Maternal Environment on Seed Germination and Seedling Vigor of Petunia x hybrida under Different Abiotic Stresses. Plants 2021, 10, 581. [Google Scholar] [CrossRef]
- Wei, J.; Liu, X.; Li, L.; Zhao, H.; Liu, S.; Yu, X.; Shen, Y.; Zhou, Y.; Zhu, Y.; Shu, Y.; et al. Quantitative proteomic, physiological and biochemical analysis of cotyledon, embryo, leaf and pod reveals the effects of high temperature and humidity stress on seed vigor formation in soybean. BMC Plant Biol. 2020, 20, 127. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Ma, H.; Song, L.; Shu, Y.; Gu, W. Comparative proteomics analysis reveals the mechanism of pre-harvest seed deterioration of soybean under high temperature and humidity stress. J. Proteom. 2012, 75, 2109–2127. [Google Scholar] [CrossRef]
- Ling, A.; Li, X.; Hu, X.; Ma, Z.; Wu, K.; Zhang, H.; Hao, M.; Wei, S. Dynamic changes in polyphenol compounds, antioxidant activity, and PAL gene expression in different tissues of buckwheat during germination. J. Sci. Food Agr. 2018, 98, 5723–5730. [Google Scholar] [CrossRef]
- Li, X.; Dong, S.; Bai, W.; Jia, J.; Gu, R.; Zhao, C.; Liu, X.; Wang, Y. Metabolic and transcriptional regulation of phenolic conversion and tocopherol biosynthesis during germination of sesame (Sesamum indicum L.) seeds. Food Funct. 2020, 11, 9848–9857. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Ding, Y.; Wang, Q.; Wang, P.; Han, Y.; Gu, Z.; Yang, R. NaCl treatment on physio-biochemical metabolism and phenolics accumulation in barley seedlings. Food Chem. 2020, 331, 127282. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, J.L.; Austin, M.B.; Stewart, C.J.; Noel, J.P. Structure and function of enzymes involved in the biosynthesis of phenylpropanoids. Plant Physiol. Bioch. 2008, 46, 356–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Liu, C.J. Multifaceted regulations of gateway enzyme phenylalanine ammonia-lyase in the biosynthesis of phenylpropanoids. Mol. Plant 2015, 8, 17–27. [Google Scholar] [CrossRef]
- Okada, T.; Mikage, M.; Sekita, S. Molecular characterization of the phenylalanine ammonia-lyase from Ephedra sinica. Biol. Pharm. Bull. 2008, 31, 2194–2199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, H.; Wada, M.; Muraya, K.; Malik, K.; Shiraishi, T.; Ichinose, Y.; Yamada, T. Characterization of nuclear factors for elicitor-mediated activation of the promoter of the pea phenylalanine ammonia-lyase gene 1. Plant Physiol. 1995, 108, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.H.; Namasivayam, P.; Abdullah, M.P. The PAL2 promoter activities in relation to structural development and adaptation in Arabidopsis thaliana. Planta 2012, 235, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, R.J.; Grotewold, E.; Stam, M. Cis-regulatory sequences in plants: Their importance, discovery, and future challenges. Plant Cell 2022, 34, 718–741. [Google Scholar] [CrossRef]
- Foyer, C.H.; Shigeoka, S. Understanding oxidative stress and antioxidant functions to enhance photosynthesis. Plant Physiol. 2011, 155, 93–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.H.; Chu, P.; Zhou, Y.L.; Ding, Y.; Li, Y.; Liu, J.; Jiang, L.W.; Huang, S.Z. Ectopic expression of NnPER1, a Nelumbo nucifera 1-cysteine peroxiredoxin antioxidant, enhances seed longevity and stress tolerance in Arabidopsis. Plant J. 2016, 88, 608–619. [Google Scholar] [CrossRef]
- Dias, M.C.; Pinto, D.; Silva, A. Plant Flavonoids: Chemical Characteristics and Biological Activity. Molecules 2021, 26, 5377. [Google Scholar] [CrossRef]
- Zhang, F.; Wang, Y.; Liu, C.; Chen, F.; Ge, H.; Tian, F.; Yang, T.; Ma, K.; Zhang, Y. Trichoderma harzianum mitigates salt stress in cucumber via multiple responses. Ecotoxicol. Environ. Saf. 2019, 170, 436–445. [Google Scholar] [CrossRef]
- Guo, H.; Wang, L.; Yang, C.; Zhang, Y.; Zhang, C.; Wang, C. Identification of novel cis-elements bound by BplMYB46 involved in abiotic stress responses and secondary wall deposition. J. Integr. Plant Biol. 2018, 60, 1000–1014. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, E.A. Seed priming to alleviate salinity stress in germinating seeds. J. Plant Physiol. 2016, 192, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Ali, L.G.; Nulit, R.; Ibrahim, M.H.; Yien, C. Efficacy of KNO3, SiO2 and SA priming for improving emergence, seedling growth and antioxidant enzymes of rice (Oryza sativa), under drought. Sci. Rep. 2021, 11, 3864. [Google Scholar] [CrossRef]
- Fan, S.; Liu, H.; Liu, J.; Hua, W.; Li, J. BnGF14-2c Positively Regulates Flowering via the Vernalization Pathway in Semi-Winter Rapeseed. Plants 2022, 11, 2312. [Google Scholar] [CrossRef]
- Liu, S.; Liu, Y.; Liu, C.; Li, Y.; Zhang, F.; Ma, H. Isolation and Characterization of the GmMT-II Gene and Its Role in Response to High Temperature and Humidity Stress in Glycine max. Plants 2022, 11, 1503. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Dai, Y.; Zheng, C.; Yang, Y.; Chen, W.; Wang, Q.; Chandrasekaran, U.; Du, J.; Liu, W.; Shu, K. The ABI4-RbohD/VTC2 regulatory module promotes reactive oxygen species (ROS) accumulation to decrease seed germination under salinity stress. New Phytol. 2021, 229, 950–962. [Google Scholar] [CrossRef]
- Kaur, H.; Petla, B.P.; Kamble, N.U.; Singh, A.; Rao, V.; Salvi, P.; Ghosh, S.; Majee, M. Differentially expressed seed aging responsive heat shock protein OsHSP18.2 implicates in seed vigor, longevity and improves germination and seedling establishment under abiotic stress. Front. Plant Sci. 2015, 6, 713. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Wei, J.; Wang, S.; Zhang, X.; Mu, K.; Liu, S.; Ma, H. The Copper Chaperone Protein Gene GmATX1 Promotes Seed Vigor and Seedling Tolerance under Heavy Metal and High Temperature and Humidity Stresses in Transgenic Arabidopsis. Plants 2022, 11, 1325. [Google Scholar] [CrossRef] [PubMed]
- Burbulis, I.E.; Iacobucci, M.; Shirley, B.W. A null mutation in the first enzyme of flavonoid biosynthesis does not affect male fertility in Arabidopsis. Plant Cell 1996, 8, 1013–1025. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Li, Y.J.; Zhang, F.J.; Zhang, G.Z.; Jiang, X.Y.; Yu, H.M.; Hou, B.K. The Arabidopsis UDP-glycosyltransferases UGT79B2 and UGT79B3, contribute to cold, salt and drought stress tolerance via modulating anthocyanin accumulation. Plant J. 2017, 89, 85–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talaat, N.B.; Shawky, B.T. Synergistic Effects of Salicylic Acid and Melatonin on Modulating Ion Homeostasis in Salt-Stressed Wheat (Triticum aestivum L.) Plants by Enhancing Root H(+)-Pump Activity. Plants 2022, 11, 416. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Shen, Y.; Mu, K.; Cai, W.; Zhao, Y.; Shen, H.; Wang, X.; Ma, H. Phenylalanine Ammonia Lyase GmPAL1.1 Promotes Seed Vigor under High-Temperature and -Humidity Stress and Enhances Seed Germination under Salt and Drought Stress in Transgenic Arabidopsis. Plants 2022, 11, 3239. https://doi.org/10.3390/plants11233239
Zhang X, Shen Y, Mu K, Cai W, Zhao Y, Shen H, Wang X, Ma H. Phenylalanine Ammonia Lyase GmPAL1.1 Promotes Seed Vigor under High-Temperature and -Humidity Stress and Enhances Seed Germination under Salt and Drought Stress in Transgenic Arabidopsis. Plants. 2022; 11(23):3239. https://doi.org/10.3390/plants11233239
Chicago/Turabian StyleZhang, Xi, Yingzi Shen, Kebing Mu, Wanhan Cai, Yangyang Zhao, Hang Shen, Xinhui Wang, and Hao Ma. 2022. "Phenylalanine Ammonia Lyase GmPAL1.1 Promotes Seed Vigor under High-Temperature and -Humidity Stress and Enhances Seed Germination under Salt and Drought Stress in Transgenic Arabidopsis" Plants 11, no. 23: 3239. https://doi.org/10.3390/plants11233239
APA StyleZhang, X., Shen, Y., Mu, K., Cai, W., Zhao, Y., Shen, H., Wang, X., & Ma, H. (2022). Phenylalanine Ammonia Lyase GmPAL1.1 Promotes Seed Vigor under High-Temperature and -Humidity Stress and Enhances Seed Germination under Salt and Drought Stress in Transgenic Arabidopsis. Plants, 11(23), 3239. https://doi.org/10.3390/plants11233239