


Tendril Anatomy: A Tool for Correct Identification among Cucurbitaceous Taxa


 ,
, 
 ,
,  ,
,  and
and
Abstract
:1. Introduction
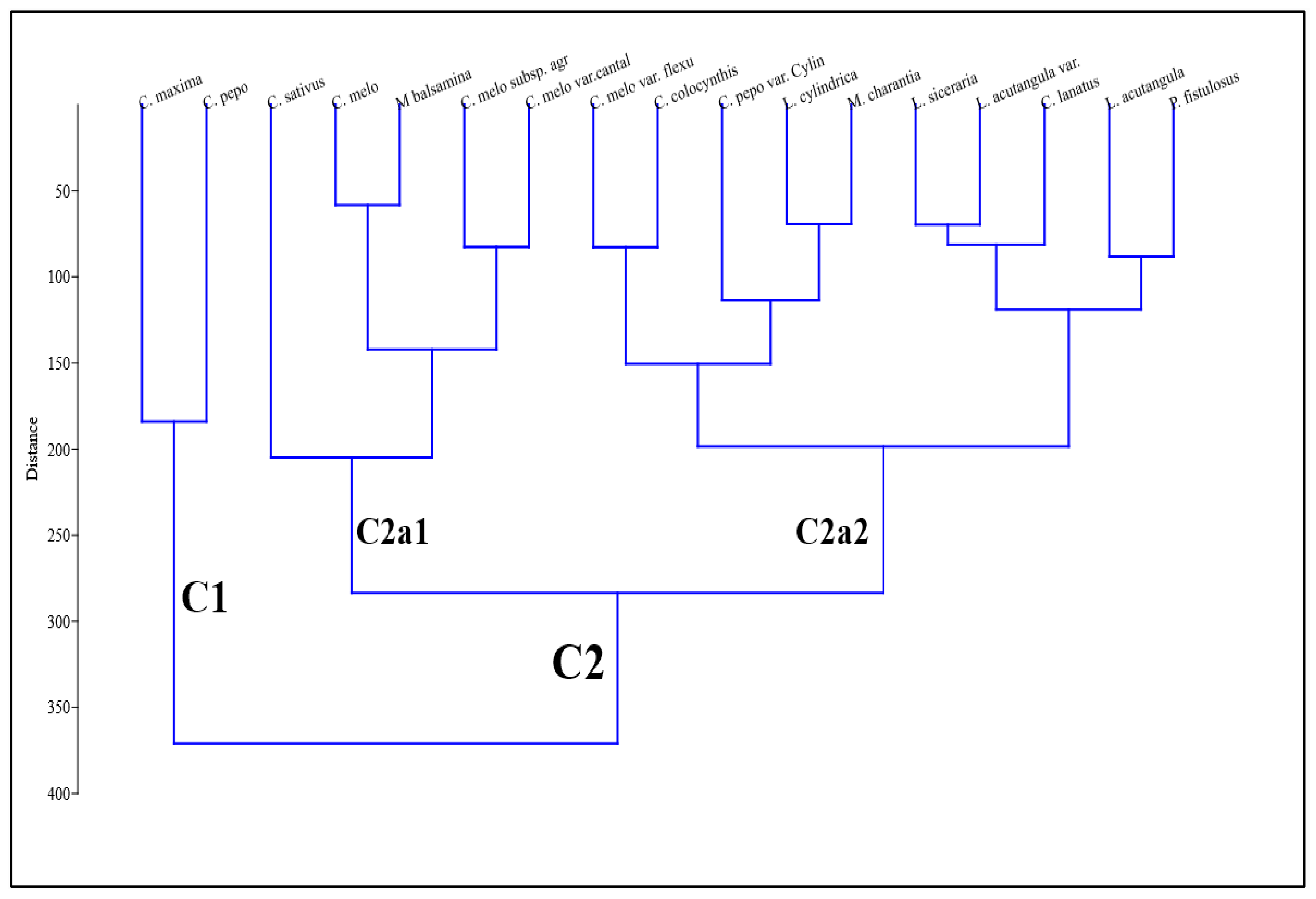
2. Results

Identification Keys Based on Cucurbitaceous Tendril Features
| 1 + Lamellar Collenchyma …………………………………………………...........................2 |
| − Angular Collenchyma…….……………………………………………………….............5 |
| 2 + Vascular bundle with round shape, irregular tendril outline…...…….............L. siceraria |
| - Lamellar and angular collenchyma………………….…………………………..…...……3 |
| 3 + Irregular vascular bundle, 6-angled tendril outline………………………………..C. pepo |
| - Oval and irregular vascular bundle………………..………………………………..……..4 |
| 4 + C shaped tendril shape…………………………………..……….C. pepo var. cylindrica |
| - Angular collenchyma cell layers……..…………………………………...……………….5 |
| 5 + Tendril outline oval, vascular bundle slightly oval……..……………..……C. colocynthis |
| - Subsidiary type vascular bundles………………………………………………………….6 |
| 6 + V shaped tendril, elliptical shape vascular bundle…………….………………...C. lanatus |
| - Irregular tendril outline…………...……………………………………………………….7 |
| 7 + Subsidiary type vascular bundle, rectangular epidermal cells……………………...C. melo |
| - Polygonal sclerenchyma cells……………………………….……………………………..8 |
| 8 + 4-angled tendril outline, irregular vascular bundle………………..C. melo subsp. agrestis |
| - Irregular tendril shape…………..………………………………………………................9 |
| 9 + Tetragonal sclerenchyma cells…………………….………………C. melo var. flexuosus - |
| Triangular to polygonal sclerenchyma cells………..………………………………………10 |
| 10 + Central vascular bundle type..………......................................C. melo var. cantalupensis |
| - Elliptical and irregular vascular bundle shape…………………………………………...11 |
| 11 + Star shape tendril outline……………………………………………………….C. sativus |
| - Rectangular epidermal cells, irregular tendril…………………….……………………12 |
| 12 + Dumbbell type vascular bundle………………………………………………..C. maxima |
| - Rounded vascular bundle shape………………………..………………………………13 |
| 13 + Irregular with hollow pith tendril sape……………………..…………..…..L. acutangula |
| - Rounded to oval vascular bundle shape………….……………………………………14 |
| 14 + Polygonal tendril outline…………………………………...… L. acutangula var.amara |
| - Hexagonal tendril outline.…………………………………………15 |
| 15 + Hexagonal to polygonal chlorenchyma cells………………………….……L. cylindrica |
| - 4-angled tendril, rectangular to polygonal chlorenchyma cells………………………16 |
| 16 + Central type vascular bundle, tetragonal sclerenchyma cells………………M. balsamina |
| - Pentagonal chlorenchyma cells…………………………………………………………17 |
| 17 + Rounded and elliptical vascular bundle type……………………………M. charantia |
| - Rectangular epidermal cells, subsidiary vascular bundle type…………………………18 |
| 18 + Polygonal slightly U shaped tendril outline………………………………….P. fistulosus |









3. Discussion
4. Materials and Methods
4.1. Study Site and Selected Species of Family Cucurbitaceae
4.2. Tendril Fixation
4.3. Histological Sectioning
4.4. Tendril Micromorphology
4.5. Light Microscopy
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schaefer, H.; Renner, S.S. Phylogenetic relationships in the order Cucurbitales and a new classification of the gourd family (Cucurbitaceae). Taxon 2011, 60, 122–138. [Google Scholar] [CrossRef]
- Jeffrey, C. A new system of Cucurbitaceae. Бoтанический Журнал 2005, 90, 332–335. [Google Scholar]
- Hutchinson, J.; Dalziel, J. Flora of West Tropical Africa; Crown Agents for Oversea Governments and Administration: London, UK, 1954; Volume 1, Part 1. [Google Scholar]
- Okoli, B. Fluted Pumpkin, Telfairia-the Under-Exploited Golden Treasure; University of Port Harcourt Press: Port Harcourt, Nigeria, 2013. [Google Scholar]
- Jeffrey, C. Key to the Cucurbitaceae of West Tropical Africa, with a Guide to Localities of Rare and Little-known Species. J. Wes Afr. Sci. Assoc. 1964, 9, 79–97. [Google Scholar]
- Ghebretinsae, A.G.; Thulin, M.; Barber, J.C. Relationships of cucumbers and melons unraveled: Molecular phylogenetics of Cucumis and related genera (Benincaseae, Cucurbitaceae). Am. J. Bot. 2007, 94, 1256–1266. [Google Scholar] [CrossRef] [PubMed]
- Omokhua-Uyi, A.G.; Abdalla, M.A.; Leonard, C.M.; Aro, A.; Uyi, O.O.; Van Staden, J.; McGaw, L.J. Flavonoids isolated from the South African weed Chromolaena odorata (Asteraceae) have pharmacological activity against uropathogens. BMC Complement. Med. Ther. 2020, 20, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Loy, B. Tendril Morphology in Bush and Vine Genotypes of Squash and Pumpkin. Cucurbit Genet. Coop. Rep. 2013, 35–36, 27–30. [Google Scholar]
- Hernandes-Lopes, J.; Sousa-Baena, M.S.; Lemos, R.C.; Corrêa, T.C.; Van Sluys, M.A.; Melo-de-Pinna, G.F.d.A. Toward understanding inflorescence development and architecture in Passiflora: Insights from comparative anatomy and expression of APETALA1. Am. J. Bot. 2019, 106, 1173–1189. [Google Scholar] [CrossRef] [PubMed]
- Sperotto, P.; Acevedo-Rodríguez, P.; Vasconcelos, T.N.; Roque, N. Towards a standardization of terminology of the climbing habit in plants. Bot. Rev. 2020, 86, 180–210. [Google Scholar] [CrossRef]
- Stace, C. Plant Taxonomy and Biosystematics; Edward Arnold Limited: London, UK, 1980. [Google Scholar]
- Josephine, A.; Chimezie, E.; Bosa, E.O. Morpho-anatomical characters of Zehneria capillacea (Schumach) C. Jeffrey and Zehneria scabra (LF) Sond Cucurbitaceae. Afr. J. Plant Sci. 2015, 9, 457–465. [Google Scholar] [CrossRef] [Green Version]
- Agbagwa, I.O.; Ndukwu, B.C. The value of morpho-anatomical features in the systematics of Cucurbita L. (Cucurbitaceae) species in Nigeria. Afr. J. Biotechnol. 2004, 3, 541–546. [Google Scholar]
- Ajuru, M.G.; Okoli, B.E. Comparative vegetative anatomy of some species of the family Cucurbitaceae Juss in Nigeria. Res. J. Bot. 2013, 8, 15. [Google Scholar] [CrossRef]
- Amonwu, I.; Okoli, B. Observations on tendrils: Contributions from Cucurbitaceae. Int. J. Life Sci. 2013, 2, 176–180. [Google Scholar] [CrossRef]
- Luchian, V.; Teodosiu, G. Research results regarding the anatomy of some medicinal plants of Cucurbitaceae. Sci. Pap. Ser. B Hortic. 2019, 63, 635–641. [Google Scholar]
- Ielciu, I.I.; Păltinean, R.; Vlase, L.; Ştefănescu, C.; Crişan, G.C. Morphological and histo-anatomical study of Bryonia alba L. (Cucurbitaceae). Not. Bot. Horti Agrobot. Cluj-Napoca 2015, 43, 47–52. [Google Scholar]
- Săvulescu, E.; Hoza, G. Anatomy study of Physalis peruviana L. species (Solanaceae). Lucr. Științ.-Univ. Științe Agron. Med. Vet. Bucur. Ser. B Hortic. 2011, 55, 643–646. [Google Scholar]
- Mohammed, I.A.; Guma, A.G.N. Anatomical diversity among certain genera of family Cucurbitaceae. Int. J. Res. Stud. Biosci. 2015, 3, 85–91. [Google Scholar]
- Ekeke, C.; Agbagwa, I.O.; Ogazie, A.C. Comparative anatomy of stem, petiole and flower stalks and its significance in the taxonomy of some members of Cucurbits. Jordan J. Biol. Sci. 2017, 10, 185–191. [Google Scholar]
- Abdulrahaman, A.; Oyedotun, R.; Oladele, F. Diagnostic significance of leaf epidermal features in the family Cucurbitaceae. Insight Bot. 2011, 1, 22–27. [Google Scholar]
- Ekeke, C.; Agogbua, J.; Okoli, B.E. Comparative anatomy of tendril and fruit stalk in Curcubitaceae Juss. from Nigeria. Int. J. Biol. Chem. Sci. 2015, 9, 1875–1887. [Google Scholar] [CrossRef] [Green Version]
- Hasson, S.M.; AL-Qaraawi, H.A.H.; Abu-Serag, N.A. Study of some taxonomic aspects for some species of cucurbitaceae in Iraq. Plant Arch. 2019, 19, 1966–1973. [Google Scholar]
- Metcalfe, C.R.; Chalk, L. Anatomy of the Dicotyledons: Leaves, stem, and wood, in relation to taxonomy, with notes on economic uses. In Anatomy of the Dicotyledons: Leaves, Stem, and Wood, in Relation to Taxonomy, with Notes on Economic Uses; Claredon Press: Oxford, UK, 1950. [Google Scholar]
- Ani, E.; Adekunle, A.A.; Kadiri, A.B.; Njoku, K.L. Rhizoremediation of hydrocarbon contaminated soil using Luffa aegyptiaca (Mill) and associated fungi. Int. J. Phytoremediation 2021, 23, 1444–1456. [Google Scholar] [CrossRef] [PubMed]
- Kadiri, A.B.; Olowokudejo, J.D. Systematic significance of foliar epidermis and tendril morphology in three West African genera of Cucurbitaceae: Momordica L., Luffa Mill. and Trichosanthes L. Webbia 2016, 71, 91–105. [Google Scholar] [CrossRef]
- Shivhare, Y.; Jain, A.P. Potential emphasis of formulated herbal cream on wound healing. Asian J. Pharm. Res. Dev. 2020, 8, 73–77. [Google Scholar] [CrossRef]
- Ishnava, K.B.; Patel, K.S. In vitro study of Praecitrulus fistulosus (Stocks) Pangalo (Cucurbitaceae) fruit–A potential candidate of Anthelmintic activity. Bull. Natl. Res. Cent. 2020, 44, 1–10. [Google Scholar] [CrossRef]
- Madhu, C.; Balaji, K.; Shankar, J.; Gowda, S.S.; Sharada, A. Biofabrication of silver nanoparticles using Praecitrullus fistulosus fruit extract exhibits in vitro antibacterial and anticancer activity. J. Drug Deliv. Sci. Technol. 2022, 72, 103329. [Google Scholar] [CrossRef]
- Okwuchukwu, A.B.; Uwabukeonye, O.C. Taxonomic significance of stem and petiole anatomy of three white varieties of Vigna unguiculata (L.) Walp. Arch. Agric. Environ. Sci. 2017, 2, 109–112. [Google Scholar]
- Akhtar, A.; Ahmad, M.; Mahmood, T.; Khan, A.M.; Arfan, M.; Abbas, Q.; Zafar, M.; Sultana, S.; Batool, R.; Fatima, A. Microscopic characterization of petiole anatomy of Asteraceous taxa of Western Himalaya-Pakistan. Microsc. Res. Tech. 2022, 85, 591–606. [Google Scholar] [CrossRef]
- Boris, P.; Andrey, K. Using petiole anatomy to identify hybrids between and species of Populus sections Aigeiros and Tacamahaca. Turczaninowia 2019, 22, 80–90. [Google Scholar]
- Jehanzeb, S.; Zafar, M.; Ahmad, M.; Sultana, S.; Zaman, W.; Ullah, F. Comparative petioler anatomy of tribe Mentheae subfamily Nepetoideae, Lamiaceae from Pakistan. Feddes Repert. 2020, 131, 163–174. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sr No. | Cucurbitaceous Taxa | Epidermal Cell Shape | Collenchyma Cell Type | Chlorenchyma Cell Shape | Sclerenchyma Cell Shape | Parenchyma Cell Shape | Vascular Bundle Shape | Vascular Bundles | Tendril Outline in Transverse View |
|---|---|---|---|---|---|---|---|---|---|
| 1. | Citrullus colocynthis (L) Schrad. | Rectangular and isodiametric | Angular | Rectangular to irregular | Tetragonal to polygonal | Polygonal | Oval | Subsidiary | Slightly oval |
| 2. | Citrullus lanatus (Thunb.) Matsum.& Nakai | Rectangular | Angular | Irregular | Polygonal to irregular | Irregular | Elliptical | Subsidiary | Irregular, slightly v shaped |
| 3. | Cucumis melo L. | Rectangular | Angular | Irregular | Irregular | Irregular | Irregular | Subsidiary | Irregular shaped |
| 4. | Cucumis melo subsp. agrestis (Naudin) Pangalo | Oval to irregular | Angular | Irregular | Polygonal | Polygonal | Irregular | Subsidiary | 4 angled |
| 5. | Cucumis melo var. flexuosus (L.) Naudin | Rectangular, square, and isodiametric | Angular | Irregular | Tetragonal to polygonal | Polygonal | Irregular | Subsidiary | Irregular shaped |
| 6. | Cucumis melo var.cantalupensis Naudin | Rectangular to square | Angular | Polygonal | Triangular to polygonal | Irregular | Irregular | Central | Irregular shaped |
| 7. | Cucumis sativus L. | Rectangular | Angular | Rectangular | Tetragonal to polygonal | Irregular | Elliptical and irregular | Subsidiary | Star-shaped |
| 8. | Cucurbita maxima Duchesne | Rectangular | Angular | Rectangular to polygonal | Irregular | Irregular | Dumbbell and irregular | Subsidiary | Irregular shaped |
| 9. | Cucurbita pepo L. | Rectangular to square | Lamellar and angular | Polygonal | Irregular | Irregular | Irregular | Subsidiary | Six angled |
| 10. | Cucurbita pepo var. cylindrica | Rectangular, square to polygonal | Lamellar and angular | Polygonal | Irregular | Irregular | Oval and irregular | Subsidiary | Slightly c shaped |
| 11. | Lagenaria siceraria (Molina) Standl. | Rectangular | Lamellar | Rectangular to irregular | Irregular | Irregular | Rounded | Subsidiary | Irregular with hollow pith |
| 12. | Luffa acutangula (L.) Roxb. | Irregular | Angular | Irregular | Irregular | Irregular | Rounded | Subsidiary | Irregular with hollow pith |
| 13. | Luffa acutangula var. amara C.B.Clarke | Rectangular, square to polygonal | Angular | Polygonal | Polygonal and irregular | Irregular | Round and oval | Subsidiary | Irregular polygonal |
| 14. | Luffa cylindrica (L) M.Roem | Rectangular to square | Angular | Hexagonal to polygonal | Tetragonal to polygonal | Irregular | Round and elliptical | Subsidiary | Six angled |
| 15. | Momordica charantia L. | Rectangular to irregular | Angular | Pentagonal to polygonal | Irregular | Irregular | Rounded and elliptical | Subsidiary | 4 angled |
| 16 | Momordica balsamina L. | Pentagonal to hexagonal | Angular | Rectangular to polygonal | Tetragonal to hexagonal | Pentagonal to irregular | Oval and irregular | Central | 4 angled |
| 17 | Praecitrullus fistulosus (Stocks) Pangalo | Rectangular | Angular | Pentagonal to polygonal | Irregular | Irregular | Irregular | Subsidiary | Polygonal, slightly U shaped |
| Sr No. | Cucurbitaceous Taxa | No. Of Vascular Bundles | Epidermal Cell Layer | Collenchyma Cell Layer | Chlorenchyma Cell Layer | Sclerenchyma Cell Layer | Parenchyma Cell Layer | Vessel Elements |
|---|---|---|---|---|---|---|---|---|
| 1. | Citrullus colocynthis (L) Schrad. | 4 | 1 | 3–4 | 1–2 | 3–5 | 4 | 6 |
| 2. | Citrullus lanatus (Thunb.) Matsum.& Nakai | 6 | 1 | 2–3 | 1 | 3–4 | 4 | 5 |
| 3. | Cucumis melo L. | 5 | 1 | 3 | 1 | 3 | 4 | 8 |
| 4. | Cucumis melo subsp. agrestis (Naudin) Pangalo | 5 | 1 | 3 | 1–2 | 2–3 | 4 | 8 |
| 5. | Cucumis melo var. flexuosus (L.) Naudin | 7 | 1 | 3 | 2 | 2–3 | 3 | 5 |
| 6. | Cucumis melo var.cantalupensis Naudin | 6 | 1 | 3 | 2 | 2–3 | 4 | 4 |
| 7. | Cucumis sativus L. | 5 | 1 | 4 | 1 | 2–4 | 6 | 15 |
| 8. | Cucurbita maxima Duchesne | 7 | 1 | 3 | 2 | 5 | 4 | 9 |
| 9. | Cucurbita pepo L. | 6 | 1 | 6 | 2 | 3–4 | 4 | 7 |
| 10. | Cucurbita pepo var. cylindrica | 7 | 1 | 4 | 2 | 3–4 | 3 | 9 |
| 11. | Lagenaria siceraria (Molina) Standl. | 7 | 1 | 3–4 | 2–3 | 7–9 | 5 | 8 |
| 12. | Luffa acutangula (L.) Roxb. | 8 | 1 | 5 | 1 | 6–8 | 3 | 8 |
| 13. | Luffa acutangula var.amara C.B.Clarke | 12 | 1 | 6 | 2 | 6 | 2 | 9 |
| 14. | Luffa cylindrica (L) M.Roem | 8 | 1 | 5 | 1 | 6 | 3 | 9 |
| 15. | Momordica charantia L. | 7 | 1 | 5 | 1 | 4 | 3 | 9 |
| 16 | Momordica balsamina L. | 3 | 1 | 3–5 | 1 | 1 | 3–4 | 10 |
| 17 | Praecitrullus fistulosus (Stocks) Pangalo | 10 | 1 | 2–5 | 2 | 4 | 5 | 5 |
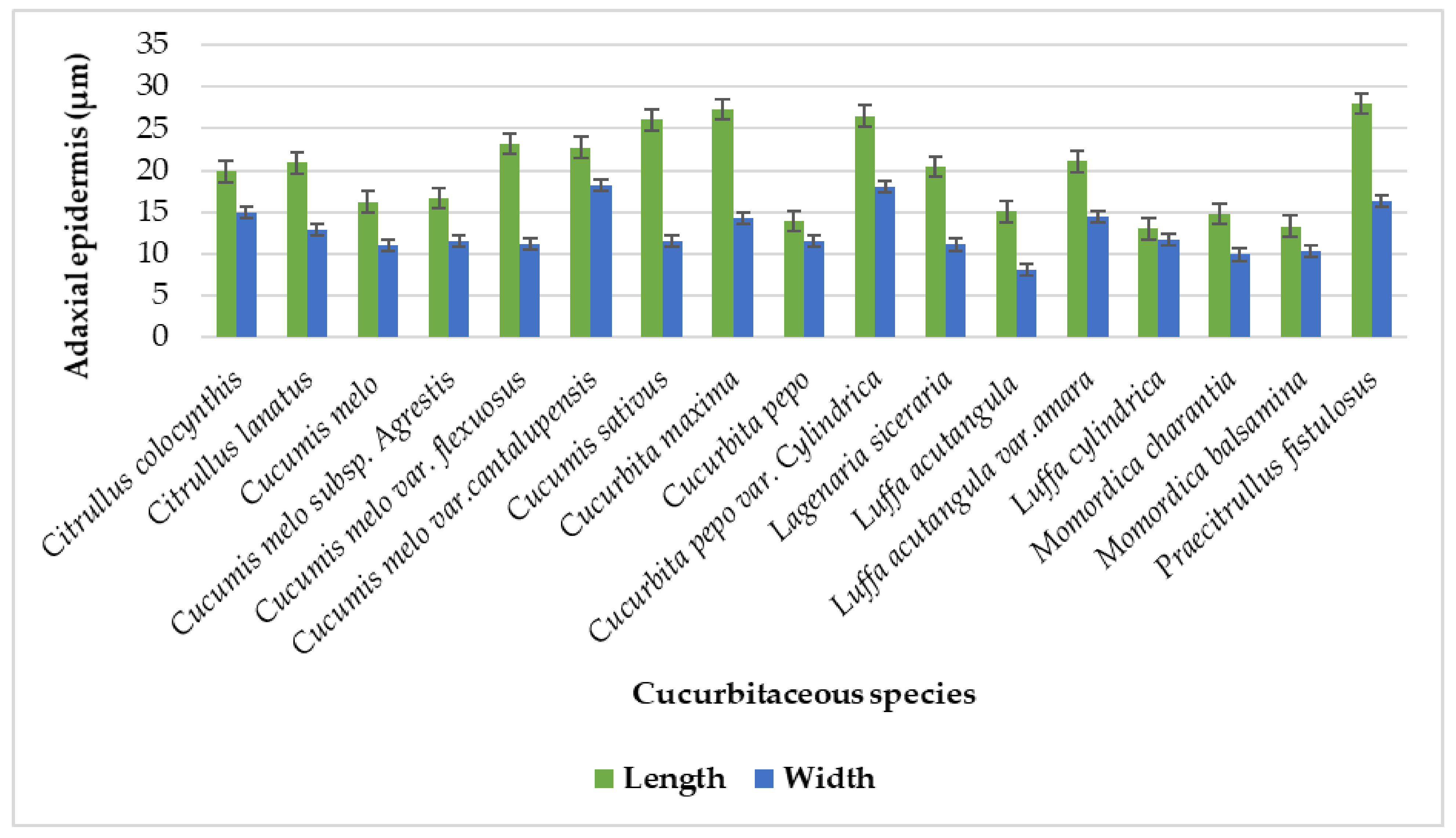
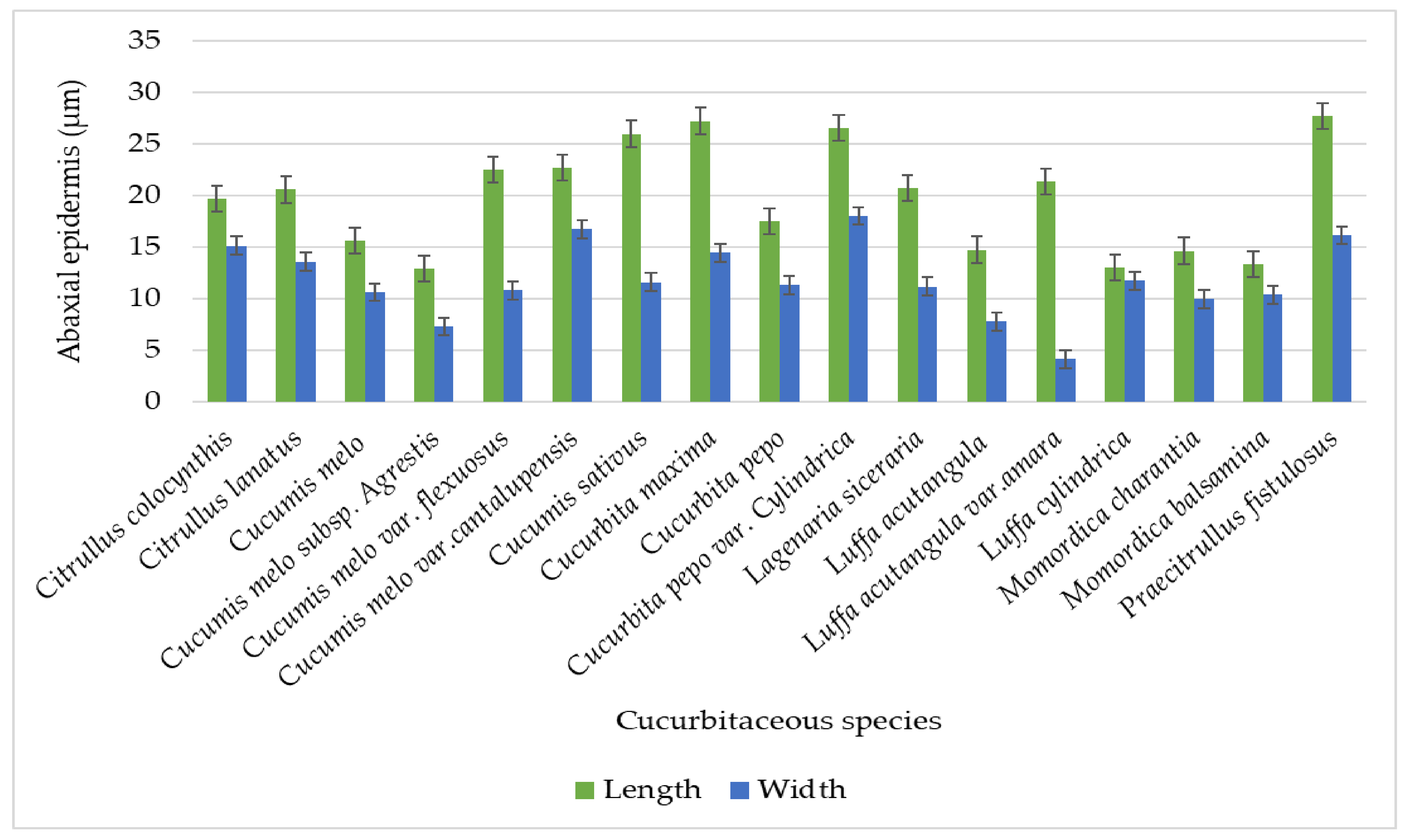
| Sr No. | Cucurbitaceous taxa | L x W | Tendril Diameter (µm) | Upper Epidermal Cell (µm) | Lower Epidermis Cell (µm) | Collenchyma Cell (µm) |
|---|---|---|---|---|---|---|
| 1. | Citrullus colocynthis (L) Schrad. | L | 314 − 326.25 = 321.65 ± 5.02 | 16.75 − 23.25 = 19.85 ± 2.57 | 16.75 − 22.50 = 19.65 ± 2.25 | 10.50 − 21.25 = 16.80 ± 4.74 |
| W | 297 − 304.75 = 301.10 ± 2.77 | 12.25 − 17.75 = 14.95 ± 2.13 | 12.75 − 18 = 15.10 ± 2.19 | 4.25 − 16.75 = 11 ± 4.72 | ||
| 2. | Citrullus lanatus (Thunb.) Matsum.& Nakai | L | 512.50 − 517.75 = 514.95 ± 2.18 | 15.75 − 25.25 = 20.90 ± 3.75 | 14.75 − 25.75 = 20.55 ± 4.93 | 16.25 − 23.25 = 19.80 ± 2.56 |
| W | 250 − 258.75 = 253.50 ± 3.24 | 10 − 20 = 12.9 ± 4.14 | 10.25 − 20.50 = 13.55 ± 4.12 | 7.50 − 12.50 = 10.05 ± 1.97 | ||
| 3. | Cucumis melo L. | L | 126.75 − 145.25 = 134.10 ± 7.44 | 10.50 − 21.75 = 16.20 ± 4.50833 | 10 − 20.75 = 15.55 ± 4.28442 | 12.00 − 21.25 = 16.20 ± 3.77 |
| W | 122.75 − 138 = 130 ± 6.38 | 8.75 − 13.75 = 11.05 ± 2.07 | 8.75 − 13 = 10.6 ± 1.85 | 7.50 − 13.75 = 10.65 ± 2.75 | ||
| 4. | Cucumis melo subsp. agrestis (Naudin) Pangalo | L | 200.50 − 212.50 = 205.35 ± 5 | 3.75 − 25.50 = 16.70 ± 9.13 | 4.75 − 25.50 = 12.85 ± 9.49 | 13.00 − 35.50 = 25.50 ± 9.78 |
| W | 183.50 − 188.50 = 186.30 ± 2.13 | 3.75 − 18 = 11.55 ± 6.18 | 2.50 − 15.25 = 7.25 ± 4.97 | 8.25 − 25.50 = 15.45 ± 6.54 | ||
| 5. | Cucumis melo var. flexuosus (L.) Naudin | L | 325.25 − 333 = 328.85 ± 3.18 | 13 − 50.25 = 23.15 ± 15.28 | 13.50 − 45.25 = 22.50 ± 12.90 | 11 − 28 = 18.50 ± 6.98 |
| W | 250.25 − 267.75 = 257.30 ± 7.73 | 7.75 − 16.75 = 11.15 ± 3.81 | 8 − 14.25 = 10.75 ± 2.97 | 2.75 − 13.25 = 8.5 ± 3.83813 | ||
| 6. | Cucumis melo var.cantalupensis Naudin | L | 250.50 − 256.50 = 253.75 ± 2.384 | 18.75 − 26.25 = 22.7500 ± ± 2.87 | 19.50 − 26.50 = 22.6500 ± 2.831 | 13.00 − 24.75 = 18.7000 ± 4.396 |
| W | 225 − 231.25 = 227.60 ± 2.36 | 13.75 − 23.25 = 18.20 ± 3.40221 | 13.25 − 19.50 = 16.70 ± 2.558 | 12 − 14.25 = 13.1500 ± 0.91 | ||
| 7. | Cucumis sativus L. | L | 524.75 − 531.25 = 527.40 ± 2.71 | 20.75 − 30.00 = 26.0500 ± 4.052 | 17.75 − 30.50 = 25.9500 ± 5.7047 | 23.25 − 33.50 = 27.8500 ± 3.7358 |
| W | 225.00 − 238.25 = 229.40 ± 5.375 | 10.25 − 13.00 = 11.5500 ± 1.242 | 10.25 − 13.00 = 11.5500 ± 1.267 | 13.75 − 21.25 = 17.7000 ± 2.808 | ||
| 8. | Cucurbita maxima Duchesne | L | 589.75 − 600.25 = 595.60 ± 3.9115 | 19.25 − 42.25 = 27.3000 ± 10.24 | 19.50 − 41.00 = 27.2000 ± 9.8256 | 12.25 − 20.00 = 16.8500 ± 3.5820 |
| W | 486.25 − 494.75 = 489.60 ± 3.3241 | 9.25 − 20.00 = 14.2500 ± 4.10 | 9.75 − 19.75 = 14.4000 ± 3.91152 | 8.75 − 13.25 = 10.7000 ± 1.68077 | ||
| 9. | Cucurbita pepo L. | L | 650.00 − 662.75 = 656.10 ± 4.735 | 8.75 − 19.75 = 13.9000 ± 4.211 | 8.00 − 20.00 = 13.5000 ± 4.53114 | 9.50 − 15.25 = 12.8500 ± 2.29538 |
| W | 315.50 − 321.25 = 317.60 ± 2.1837 | 7.50 − 17.50 = 11.4500 ± 3.8786 | 7.25 − 18.00 = 11.3000 ± 4.298 | 7.25 − 14.75 = 10.4500 ± 3.0893 | ||
| 10. | Cucurbita pepo var. cylindrica | L | 361.25 − 368.00 = 364.45 ± 2.6774 | 22.50 − 31.25 = 26.5000 ± 3.579 | 22.75 − 31.00 = 26.5500 ± 3.188 | 16.75 − 25.00 = 21.7500 ± 3.292 |
| W | 162.75 − 170.25 = 1.67.00 ± 2.93 | 13.00 − 22.50 = 18.0500 ± 3.692 | 13.25 − 22.00 = 18.0000 ± 3.592 | 10.50 − 19.25 = 14.8500 ± 3.223 | ||
| 11. | Lagenaria siceraria (Molina) Standl. | L | 471.25 − 487.75 = 477.85 ± 6.125 | 14.50 − 27.50 = 20.4500 ± 5.874 | 14.25 − 27.75 = 20.7000 ± 6.2759 | 22.50 − 30.50 = 27.7500 ± 3.172 |
| W | 210.00 − 223.75 = 217.65 ± 5.641 | 9.75 − 13.00 = 11.1000 ± 1.526 | 9.50 − 13.00 = 11.1500 ± 1.61632 | 8.75 − 12.75 = 10.7000 ± 1.71756 | ||
| 12. | Luffa acutangula (L.) Roxb. | L | 250 − 676.25 = 582.85 ± 186.37 | 8.50 − 20.50 = 15.05 ± 4.56002 | 7.25 − 20.50 = 14.7 ± 5.03550 | 13.00 − 28.50 = 21.20 ± 6.13 |
| W | 251.75 − 270.25 = 258.85 ± 7.86 | 5.25 − 10.50 = 8.05 ± 1.89 | 5.00 − 10.25 = 7.75 ± 1.87 | 7.50 − 17.50 = 12.20 ± 3.7 | ||
| 13. | Luffa acutangula var.amara C.B.Clarke | L | 451.25 − 480.50 = 472.45 ± 11.99 | 17.50 − 26 = 21.05 ± 4.19 | 17.75 − 27.50 = 21.3 ± 4.8 | 15.50 − 25 = 20.30 ± 3.83 |
| W | 219.75 − 252.75 = 229.20 ± 13.64 | 12.50 − 17 = 14.4 ± 1.82 | 12.75 − 16.25 = 4.1 ± 1.47479 | 12.75 − 22.75 = 15.9500 ± 3.98 | ||
| 14. | Luffa cylindrica (L) M.Roem | L | 401.25 − 413.75 = 406.2 ± 4.84897 | 11.25 − 15.75 = 13.0000 ± 1.677 | 11.50 − 15.75 = 13.0000 ± 1.6488 | 15.75 − 23.25 = 19.7000 ± 3.2948 |
| W | 172.25 − 180.25 = 175.75 ± 2.915 | 8.00 − 15.25 = 11.6500 ± 2.637 | 8.25 − 15.25 = 11.7000 ± 2.51496 | 13.00 − 18.00 = 15.0500 ± 2.041 | ||
| 15. | Momordica charantia L. | L | 350.00 − 357.75 = 353.25 ± 3.177 | 11.75 − 17.00 = 14.7500 ± 2.186 | 12.00 − 17.00 = 14.6000 ± 2.2262 | 16.25 − 20.00 = 18.2500 ± 1.3578 |
| W | 160.75 − 167.25 = 164.55 ± 2.7064 | 8.75 − 11.00 = 9.9000 ± 0.96177 | 9.00 − 10.75 = 9.9500 ± 0.71589 | 9.25 − 11.75 = 10.5500 ± 1.10962 | ||
| 16 | Momordica balsamina L. | L | 122.25 − 124.0 = 123.05 ± 0.81777 | 8.75 − 16.25 = 13.3000 ± 2.9811 | 8.50 − 16.00 = 13.3000 ± 3.15436 | 14.00 − 25.50 = 19.4500 ± 5.5884 |
| W | 112.25 − 114.00 = 112.95 ± 0.64711 | 7.75 − 12.25 = 10.3000 ± 1.6240 | 8.00 − 12.00 = 10.3500 ± 1.49583 | 12.75 − 14.75 = 13.8500 ± 0.89443 | ||
| 17 | Praecitrullus fistulosus (Stocks) Pangalo | L | 575.25 − 584.00 = 578.95 ± 3.49 | 22.50 − 32.75 = 28.0000 ± 4.172 | 23.50 − 33.00 = 27.6500 ± 4.207 | 17.25 − 26.25 = 22.2000 ± 3.858 |
| W | 238.00 − 244.75 = 241.90 ± 2.7477 | 13.00 − 19.25 = 16.2500 ± 2.417 | 13.25 − 18.75 = 16.1000 ± 2.043 | 10.50 − 20.00 = 14.6000 ± 3.529 |
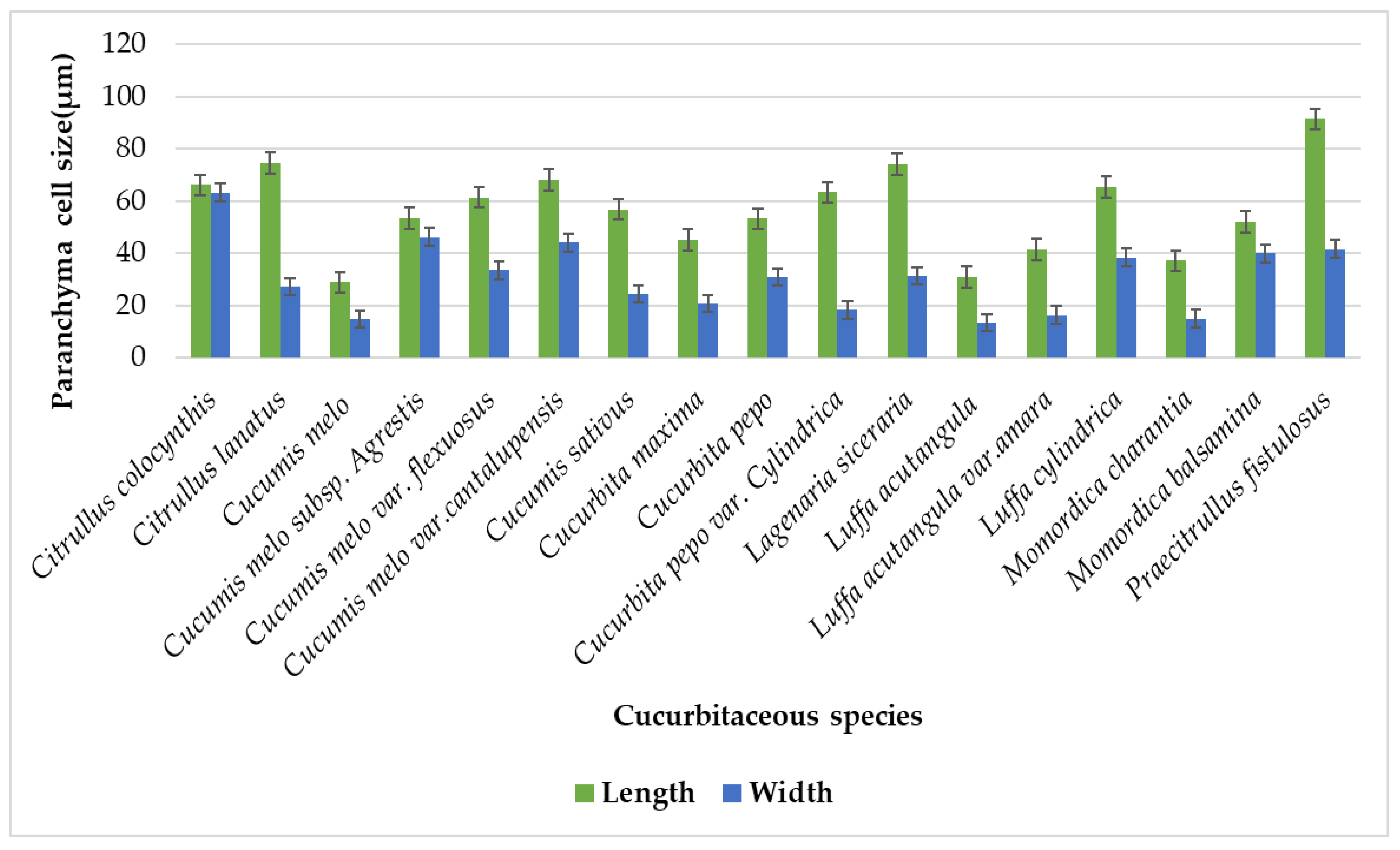
| Sr No. | Cucurbitaceous taxa | L x W | Chlorenchyma Cell (µm) | Sclerenchyma Cell (µm) | Parenchyma Cell (µm) | Vessel Elements(µm) | Vascular Bundle (µm) |
|---|---|---|---|---|---|---|---|
| 1. | Citrullus colocynthis (L) Schrad. | L | 31.75 − 50.25 = 40.50 ± 7.65 | 11.25 − 25 = 25 ± 5.76 | 50.50 − 84.75 = 66.25 ± 15.04 | 18.00 − 26.25 = 21.70 ± 3.52 | 135.50 − 188 = 161.55 ± 21.20 |
| W | 12.00 − 19.25 = 16.05 ± 3.04 | 10.50 − 22.25 = 16.10 ± 4.73 | 47.00 − 77.25 = 63.35 ± 13.40 | 5.70 − 9.90 = 7.82 ± 1.69 | 74.25 − 90.75 = 81.40 ± 6.79 | ||
| 2. | Citrullus lanatus (Thunb.) Matsum.& Nakai | L | 16.25 − 27.00 = 21 ± 4.25 | 10.75 − 27.75 = 22.40 ± 6.76 | 38.75 − 101 = 74.65 ± 28.14 | 25.50 − 37.50 = 29.05 ± 4.85 | 89.50 − 200.25 = 146.15 ± 48.33 |
| W | 8.25 − 14.75 = 10.65 ± 2.56 | 8.50 − 21 = 16.10 ± 4.84 | 26.25 − 28.75 = 27.35 ± 0.96 | 15 − 25 = 18.30 ± 4.44 | 27 − 89.75 = 64.25 ± 29.42 | ||
| 3. | Cucumis melo L. | L | 18.75 − 26.25 = 22.15 ± 2.98 | 16.75 − 28 = 22.10 ± 4.83 | 18 − 43.75 = 29 ± 11.15 | 11.25 − 16.25 = 13.75 ± 2.26 | 89 − 113.75 = 99.90 ± 10.29 |
| W | 8.75 − 11.75 = 10.35 ± 1.23 | 9.75 − 22 = 14.90 ± 5.23 | 9.75 − 22 = 14.90 ± 5.23 | 8.50 − 11.25 = 9.55 ± 1.10 | 30.50 − 55.25 = 38.20 ± 9.97 | ||
| 4. | Cucumis melo subsp. agrestis (Naudin) Pangalo | L | 15.50 − 25.25 = 20.60 ± 4.05 | 8.25 − 20.25 = 13.55 ± 4.54 | 20.50 − 77.75 = 53.55 ± 26.50 | 9.75 − 13.25 = 11.50 ± 1.47 | 47.25 − 97 = 74 ± 19.78 |
| W | 7.75 − 10.25 = 8.9000 ± 1.24 | 2.75 − 18.50 = 9 ± 6.02 | 27.75 − 62.75 = 46.25 ± 13.21 | 5.25 − 10.25 = 7.90 ± 1.78 | 22.50 − 33.75 = 28.20 ± 4.25 | ||
| 5. | Cucumis melo var. flexuosus (L.) Naudin | L | 13 − 41.25 = 25.65 ± 10.97 | 5.25 − 20.50 = 16.15 ± 6.39 | 23.25 − 112.50 = 61.50 ± 37.46 | 7.75 − 13.75 = 11.75 ± 2.60 | 75.25 − 163 = 117.10 ± 39.43 |
| W | 10.50 − 20 = 14.35 ± 4.27 | 3.75 − 15.25 = 10.15 ± 4.71 | 15.50 − 55.25 = 33.50 ± 14.27 | 5.50 − 13.25 = 9.60 ± 3.02 | 26.75 − 51.50 = 42.55 ± 9.86 | ||
| 6. | Cucumis melo var.cantalupensis Naudin | L | 42.00 − 46.25 = 43.80 ± 1.67 | 19.25 − 25.75 = 21.35 ± 2.61 | 49.25 − 86.25 = 68.10 ± 13.73 | 29.75 − 40.75 = 34.95 ± 4.48 | 77.25 − 106.25 = 89.10 ± 11.14 |
| W | 24.50 − 29.50 = 27.45 ± 2.06 | 10.50 − 14.25 = 12.20 ± 1.47 | 36.25 − 52.75 = 44.15 ± 6.78 | 21.25 − 37.75 = 30.45 ± 6.66 | 26.25 − 37.25 = 30.95 ± 4.10 | ||
| 7. | Cucumis sativus L. | L | 25.50 − 62.75 = 44.25 ± 13.87 | 10.25 − 30 = 19.05 ± 8.56 | 13.75 − 88 = 56.95 ± 32.34 | 38.25 − 47.50 = 42.55 ± 4.09 | 151.75 − 301.25 = 224.15 ± 56.12 |
| W | 13.75 − 20.25 = 17 ± 2.54 | 7.75 − 20.25 = 14.25 ± 5.60 | 11.00 − 39.25 = 24.45 ± 10.77 | 20.50 − 38 = 29.25 ± 7.35 | 101.50 − 202.25 = 125.30 ± 43.31 | ||
| 8. | Cucurbita maxima Duchesne | L | 20.50 − 51.25 = 33.60 ± 11.74 | 13.75 − 27.75 = 20.70 ± 5.67 | 24.75 − 64.50 = 45.15 ± 16.02 | 24.75 − 43 = 35.10 ± 6.83 | 30.50 − 38.75 = 34 ± 3.15 |
| W | 11.75 − 18 = 13.55 ± 2.60 | 9.50 − 12.50 = 11.45 ± 1.19 | 14.50 − 28.25 = 20.80 ± 6.11 | 18.75 − 27.75 = 22.90 ± 3.92 | 24.75 − 30.75 = 27.35 ± 2.24 | ||
| 9. | Cucurbita pepo L. | L | 21.75 − 29.25 = 26.60 ± 3.12 | 20.25 − 30 = 24.05 ± 3.68 | 30.50 − 66.25 = 53.35 ± 14.31 | 25.75 − 35.25 = 30.80 ± 3.69 | 37.25 − 44.50 = 41.55 ± 3.12 |
| W | 7.25 − 19.75 = 10.95 ± 5.05 | 9.75 − 14.25 = 11.25 ± 1.76 | 17.75 − 50.75 = 30.95 ± 12.30 | 15.25 − 33.75 = 24.75 ± 8.65 | 24.50 − 29.50 = 27.55 ± 1.93 | ||
| 10. | Cucurbita pepo var. cylindrica | L | 26.75 − 46.25 = 33.55 ± 7.57 | 19.50 − 25.25 = 23.05 ± 2.34 | 47.25 − 98.75 = 63.45 ± 20.67 | 16.25 − 22.25 = 19 ± 2.34 | 22.50 − 28.75 = 26.10 ± 2.87 |
| W | 12.50 − 28 = 19.10 ± 7.40 | 12.25 − 18 = 16.35 ± 2.32 | 12.25 − 25 = 18.45 ± 5.13 | 11.25 − 15 = 13.30 ± 1.52 | 17.25 − 21.25 = 19.35 ± 1.51 | ||
| 11. | Lagenaria siceraria (Molina) Standl. | L | 20 − 46.25 = 32.50 ± 10.08 | 22.50 − 31.25 = 27.60 ± 3.43 | 43.75 − 105.25 = 74.15 ± 22.10 | 20 − 37.50 = 28.30 ± 7.28 | 63.25 − 103.75 = 86.25 ± 17.20 |
| W | 11.75 − 17.50 = 13.40 ± 2.38 | 11.75 − 20.25 = 15.05 ± 3.65 | 26.75 − 36.50 = 31.40 ± 4.32 | 18 − 30.75 = 24.80 ± 5.14 | 25.50 − 63.50 = 38.95 ± 14.65 | ||
| 12. | Luffa acutangula (L.)Roxb. | L | 17.75 − 45.50 = 30.50 ± 10.42 | 15.75 − 35.75 = 26.70 ± 8.98 | 24.25 − 38.75 = 31 ± 5.76086 | 25.50 − 32.75 = 27.60 ± 3.04 | 76.25 − 125.25 = 109.85 ± 19.32 |
| W | 11.25 − 15.25 = 13 ± 1.60 | 11.00 − 15.50 = 13.50 ± 1.81 | 10 − 15.25 = 13.50 ± 2.05 | 13.25 − 30 = 23.95 ± 6.43 | 46.50 − 101.75 = 87.60 ± 23.57 | ||
| 13. | Luffa acutangula var. amara C.B.Clarke | L | 15.25 − 45 = 29.85 ± 10.89 | 30.75 − 46 = 38.05 ± 6.45 | 23.25 − 82.81 = 41.61 ± 24.37 | 12.75 − 25.25 = 17 ± 5.54 | 76.25 − 125.25 = 109.90 ± 19.33 |
| W | 10.25 − 15.75 = 13.45 ± 2.25 | 13.00 − 22.75 = 17.95 ±3.97 | 12.75 − 20.75 = 16.40 ± 3.83± | 10.25 − 22.75 = 15.80 ± 5.58 | 46.50 − 101.75 = 87.60 ± 23.57 | ||
| 14. | Luffa cylindrica (L) M.Roem | L | 17.50 − 37.75 = 28.80 ± 7.97 | 21.25 − 31.25 = 26.50 ± 3.99 | 53 − 75.75 = 65.34 ± 9.45473 | 10.25 − 31.25 = 21.70 ± 9.30 | 101 − 139.25 = 119.90 ± 17.32 |
| W | 11.00 − 20.50 = 15.60 ± 3.76 | 11.25 − 17.50 = 13.80 ± 2.68 | 19.50 − 45.25 = 38.50 ± 10.88 | 7.50 − 27.75 = 17.55 ± 8 | 76 − 101.75 = 84.75 ± 10.84 | ||
| 15. | Momordica charantia L. | L | 31.25 − 53.50 = 45.20 ± 9.05 | 21.25 − 26.25 = 24.45 ± 2.01 | 30.25 − 41.75 = 37.20 ± 4.36 | 22.25 − 31 = 26.85 ± 3.83 | 95.50 − 112.75 = 102.90 ± 7.33 |
| W | 11.25 − 28.50 = 19.35 ± 6.64 | 11.50 − 18 = 14.95 ± 2.63 | 11.25 − 19.50 = 15.05 ± 3.83 | 17.50 − 23.75 = 20.75 ± 2.89 | 71 − 77.25 = 73.70 ± 2.61 | ||
| 16 | Momordica balsamina L. | L | 16.25 − 28.75 = 22.45 ± 4.88 | 16.25 − 21.25 = 19.60 ± 1.94 | 38.75 − 64 = 52.05 ± 9.07 | 7.25 − 11 = 9.50 ± 1.57 | 51.25 − 68.75 = 59.15 ± 7.87 |
| W | 13.75 − 20.50 = 16.70 ± 3.10 | 11.25 − 17.75 = 13.95 ± 2.58 | 31.25 − 48.75 = 39.95 ± 7.16 | 7.75 − 9.75 = 8.60 ± 0.741 | 41.25 − 50.50 = 45.90 ± 3.90 | ||
| 17 | Praecitrullus fistulosus (Stocks) Pangalo | L | 20.00 − 47.50 = 32.70 ± 11.01 | 23.25 − 31.25 = 27 ± 3.08 | 62.50 − 126.75 = 91.55 ± 26 | 13 − 22.75 = 18 ± 4.09 | 138.25 − 153.75 = 147 ± 6.25 |
| W | 13.75 − 25.50 = 19.85 ± 4.97 | 12.25 − 17.75 = 14. ± 2.22 | 20 − 75 = 41.65 ± 27.69 | 10.25 − 18 = 14.35 ± 3 | 104.50 − 127.75 = 118 ± 11.78 |
| S/N | Species Name | Locality | GPS Coordinates | Collection Date | Voucher Number | Accession Number |
|---|---|---|---|---|---|---|
| 1. | Citrullus colocynthis (L) Schrad. | Patti Bulanda | 31°15′24.67” N 71°25′28.66” E | 7 May 2022 | QAU-NA-3 | 132045 |
| 2. | Citrullus lanatus (Thunb.) Matsum.& Nakai | Mian farm | 31°17′46.31” N 71°24′41.17” E | 19 March 2022 | QAU-NA-16 | 132053 |
| 3. | Cucumis melo L. | Mian farm | 31°17′46.31” N 71°24′41.17” E | 19 March 2022 | QAU-NA-15 | 132059 |
| 4. | Cucumis melo subsp. agrestis (Naudin) Pangalo | Rakhhonda lala | 31°19′01.56” N 71°25′50.80” E | 7 May 2022 | QAU-NA-7 | 132049 |
| 5. | Cucumis melo var. flexuosus (L.) Naudin | Patti Bulanda | 31°15′24.67” N 71°25′28.66” E | 8 May 2022 | QAU-NA-6 | 132047 |
| 6. | Cucumis melo var.cantalupensis Naudin | Rakhhonda lala | 31°19′01.56” N 71°25′50.80” E | 7 May 2022 | QAU-NA-9 | 132046 |
| 7. | Cucumis sativus L. | Mian farm | 31°17′46.31” N 71°24′41.17” E | 19 March 2022 | QAU-NA-14 | 132060 |
| 8. | Cucurbita maxima Duchesne | 47 TDA | 31°31′46.92” N 71°09′56.33” E | 22 March 2022 | QAU-NA-8 | 132052 |
| 9. | Cucurbita pepo L. | 47 TDA | 31°31′46.92” N 71°09′56.33” E | 20 March 2022 | QAU-NA-13 | 132056 |
| 10. | Cucurbita pepo var. cylindrica | Jahan khan | 31°33′09.40” N 71°10′17.71” E | 20 March 2022 | QAU-NA-11 | 132051 |
| 11. | Lagenaria siceraria (Molina) Standl. | Jahan khan | 31°33′09.40” N 71°10′17.71” E | 20 March 2022 | QAU-NA-17 | 132050 |
| 12. | Luffa acutangula (L.)Roxb | Basti Thind | 31°15′31.94” N 71°27′20.64” E | 22 March 2022 | QAU-NA-2 | 132055 |
| 13. | Luffa acutangula var.amara C.B.Clarke | Darkhana wala | 31°16′13.57” N 71°25′03.28” E | 11 May 2022 | QAU-NA-5 | 132044 |
| 14. | Luffa cylindrica (L) M.Roem | Basti Thind | 31°15′31.94” N 71°27′20.64” E | 10 May 2022 | QAU-NA-10 | 132054 |
| 15. | Momordica charantia L. | Jahan khan | 31°33′09.40” N 71°10′17.71” E | 20 March 2022 | QAU-NA-1 | 132057 |
| 16. | Momordica balsamina L. | 222 TDA | 31°09′43.53” N 71°12′38.90” E | 14 July 2022 | QAU-NA-18 | 132143 |
| 17. | Praecitrullus fistulosus (Stocks) Pangalo | Fateh pur | 31°10′28.68” N 71°13′32.71” E | 10 May 2022 | QAU-NA-4 | 132048 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abbas, N.; Zafar, M.; Ahmad, M.; Althobaiti, A.T.; Ramadan, M.F.; Makhkamov, T.; Gafforov, Y.; Khaydarov, K.; Kabir, M.; Sultana, S.; et al. Tendril Anatomy: A Tool for Correct Identification among Cucurbitaceous Taxa. Plants 2022, 11, 3273. https://doi.org/10.3390/plants11233273
Abbas N, Zafar M, Ahmad M, Althobaiti AT, Ramadan MF, Makhkamov T, Gafforov Y, Khaydarov K, Kabir M, Sultana S, et al. Tendril Anatomy: A Tool for Correct Identification among Cucurbitaceous Taxa. Plants. 2022; 11(23):3273. https://doi.org/10.3390/plants11233273
Chicago/Turabian StyleAbbas, Naveed, Muhammad Zafar, Mushtaq Ahmad, Ashwaq T. Althobaiti, Mohamed Fawzy Ramadan, Trobjon Makhkamov, Yusufjon Gafforov, Khislat Khaydarov, Muhammad Kabir, Shazia Sultana, and et al. 2022. "Tendril Anatomy: A Tool for Correct Identification among Cucurbitaceous Taxa" Plants 11, no. 23: 3273. https://doi.org/10.3390/plants11233273
APA StyleAbbas, N., Zafar, M., Ahmad, M., Althobaiti, A. T., Ramadan, M. F., Makhkamov, T., Gafforov, Y., Khaydarov, K., Kabir, M., Sultana, S., Majeed, S., & Batool, T. (2022). Tendril Anatomy: A Tool for Correct Identification among Cucurbitaceous Taxa. Plants, 11(23), 3273. https://doi.org/10.3390/plants11233273