

Chemical Survey of Three Species of the Genus Rauhia Traub (Amaryllidaceae)

 ,
,  ,
, 

 ,
, 
Abstract
:1. Introduction
2. Results and Discussion
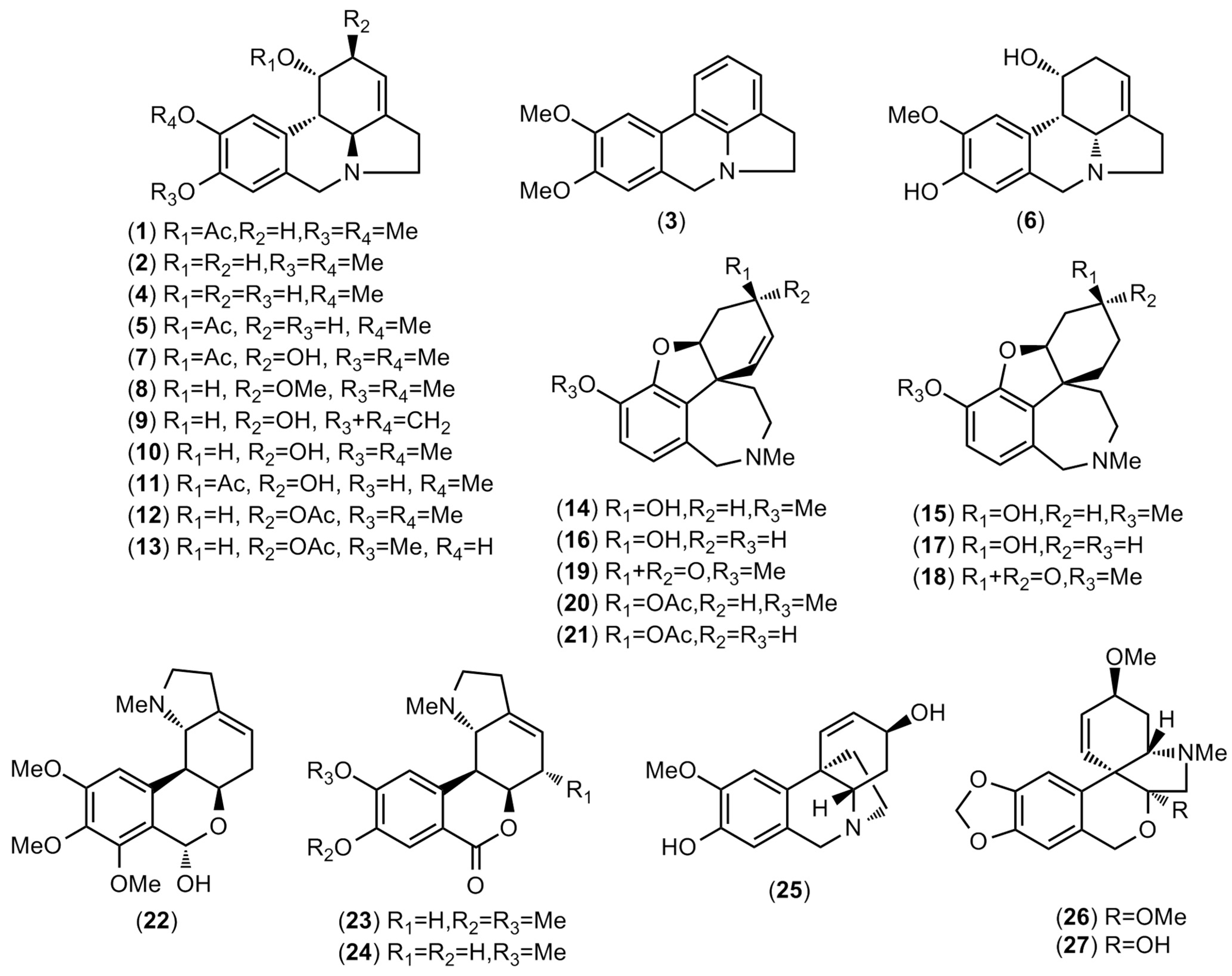
2.1. Alkaloid Profiling
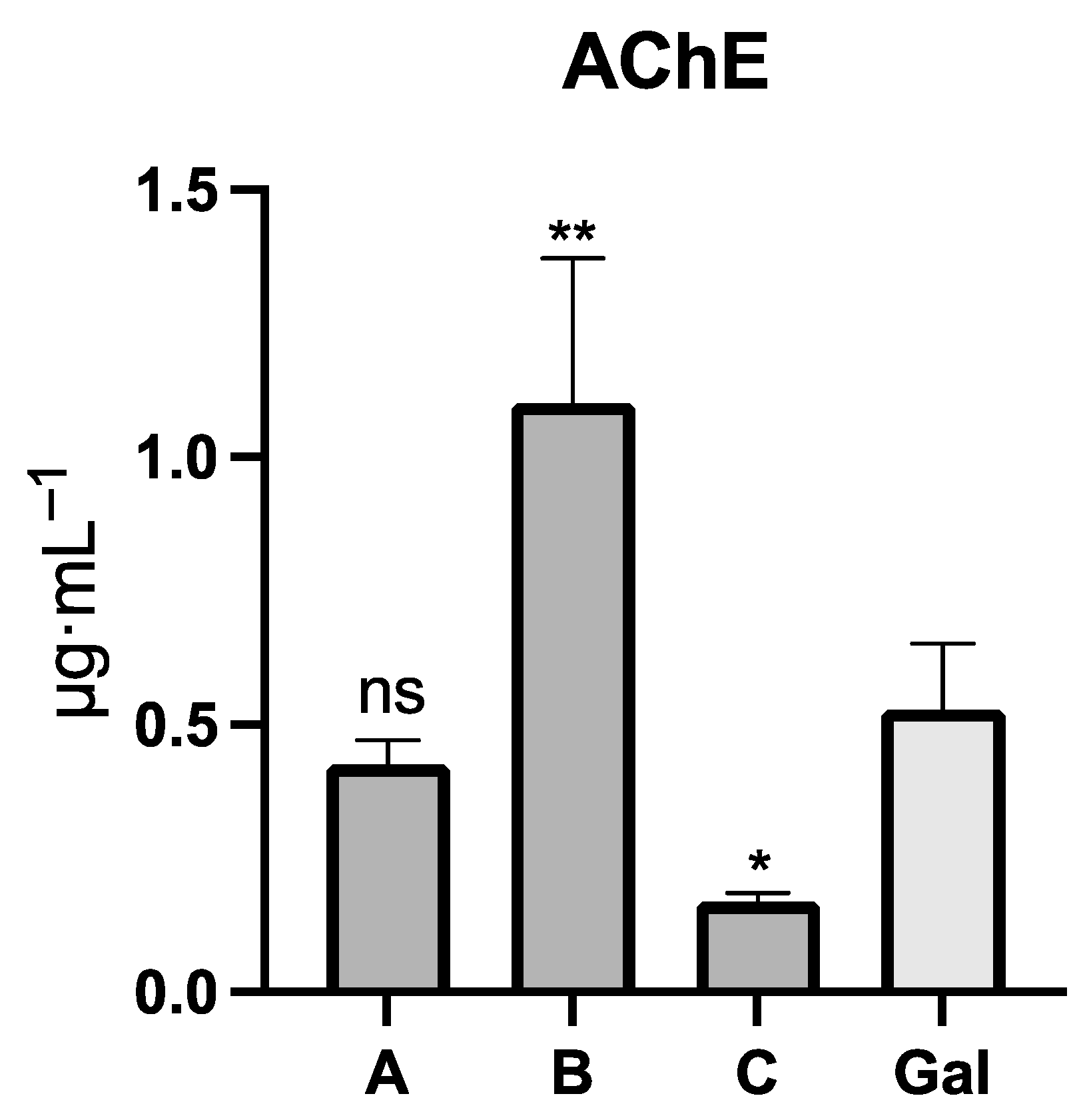
2.2. Acetylcholinesterase Inhibition
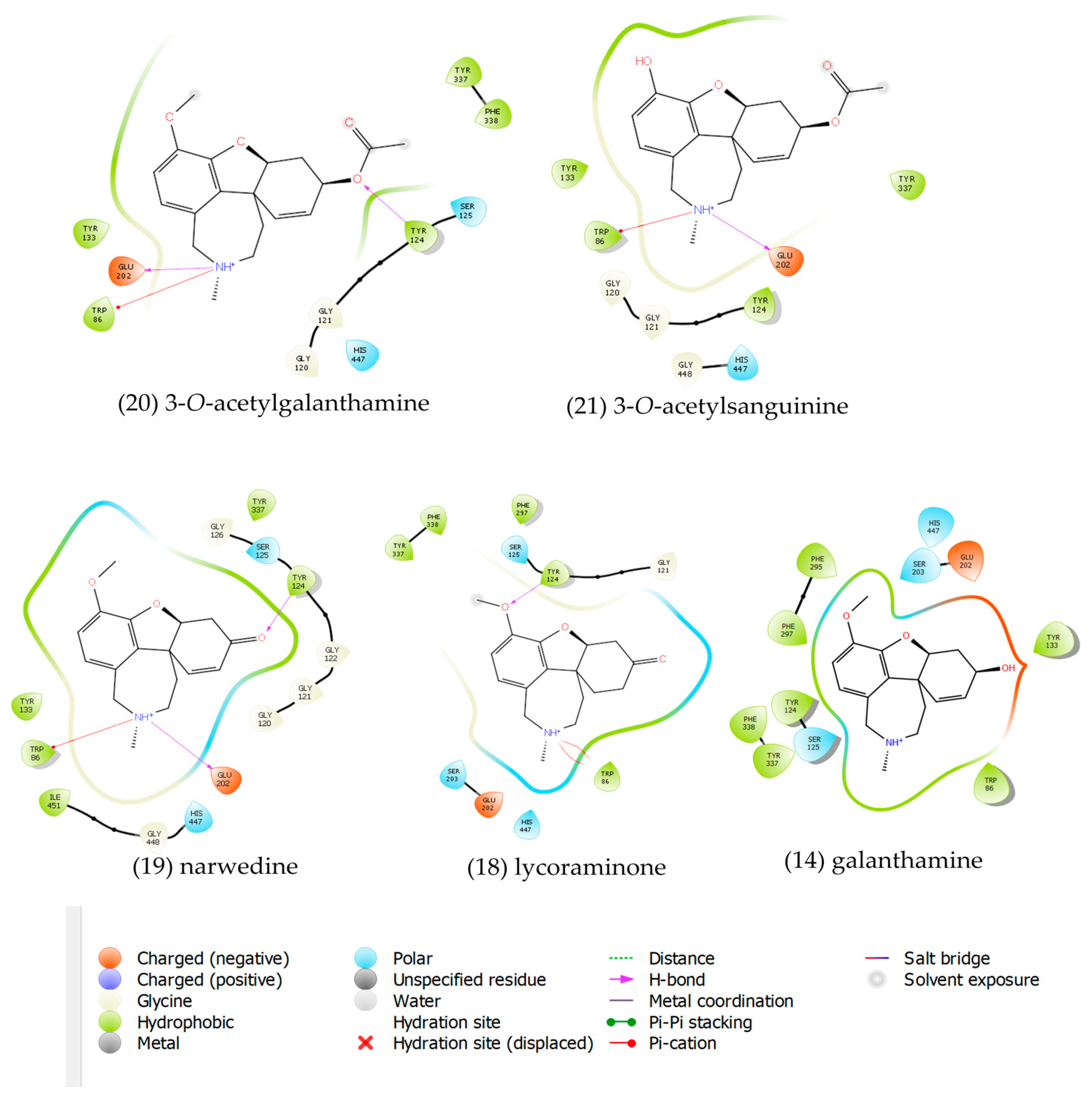
2.3. Molecular Docking
3. Materials and Methods
3.1. Plant Material Voucher
3.2. Extraction
3.3. GC-MS Analysis
3.4. Alkaloid Identification and Quantification
3.5. Enzymatic Assay
3.6. Statistical Analysis
3.7. Molecular Docking
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization–Biodiversity and Health. Available online: https://www.who.int/news-room/fact-sheets/detail/biodiversity-and-health (accessed on 25 July 2022).
- Newman, D.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feher, M.; Schimidt, J.M. Property distributions: Differences between drugs, natural products, and molecules from combinatorial chemistry. J. Chem. Inf. Comput. Sci. 2003, 43, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.-J.; Bao, J.-L.; Chen, X.-P.; Huang, M.; Wang, Y.-T. Alkaloids isolated from natural herbs as the anticancer agents. Evid. Based. Compl. Alt. 2012, 2012, 485042. [Google Scholar] [CrossRef] [Green Version]
- Bastida, J.; Lavilla, R.; Viladomat, F. Chemical and biological aspects of Narcissus alkaloids. In The Alkaloids: Chemistry and Physiology; Cordell, G.A., Ed.; Elsevier: Amsterdam, The Netherlands, 2006; Volume 63, pp. 87–179. [Google Scholar] [CrossRef]
- Meerow, A.W.; Snijman, D.A. Amaryllidaceae. In Families and Genera of Vascular Plants; Kubitzki, K., Ed.; Springer: Berlin, Germany, 1998; Volume 3, pp. 83–110. [Google Scholar]
- Konrath, E.L.; Passos, C.D.S.; Klein-Júnior, L.C.; Henriques, A.T. Alkaloids as a source of potential anticholinesterase inhibitors for the treatment of Alzheimer’s disease. J. Pharm. Pharmacol. 2013, 65, 1701–1725. [Google Scholar] [CrossRef]
- Traub, H.P. Genus Rauhia and R. peruviana, gen. & sp. nov. Plant Life 1957, 13, 73–75. [Google Scholar]
- Ravenna, P. Contribution to South American Amaryllidaceae II. Plant Life 1969, 25, 55–76. [Google Scholar]
- Traub, H.P. Amaryllid notes, 1966. Plant Life 1966, 22, 11. [Google Scholar]
- Ravenna, P. Contributions to South American Amaryllidaceae VII. Plant Life 1978, 34, 69–91. [Google Scholar]
- Ravenna, P. Contribution to South American Amaryllidaceae VII [VIII]. Plant Life 1981, 37, 57–83. [Google Scholar]
- Ravenna, P. New Rauhia species from northern Peru. Onira 2002, 7, 11–12. [Google Scholar]
- Meerow, A.W.; Nakamura, K. Two new species of Peruvian Amaryllidaceae, an expanded concept of the genus Paramongaia, and taxonomic notes in Stenomesson. Phytotaxa 2019, 416, 184–196. [Google Scholar] [CrossRef]
- Meerow, A.W.; Gardner, E.M.; Nakamura, K. Phylogenomics of the Andean tetraploid clade of the American Amaryllidaceae (subfamily Amaryllidoideae): Unlocking a polyploid generic radiation abetted by continental geodynamics. Front. Plant Sci. 2020, 11, 582422. [Google Scholar] [CrossRef] [PubMed]
- Meerow, A.W.; Guy, C.L.; Li, Q.B.; Yang, S.L. Phylogeny of the American Amaryllidaceae based on nrDNA ITS sequences. Syst. Bot. 2000, 25, 708–726. [Google Scholar] [CrossRef]
- Berkov, S.; Osorio, E.; Viladomat, F.; Bastida, J. Chemodiversity, chemotaxonomy and chemoecology of Amaryllidaceae alkaloids. In The Alkaloids: Chemistry and Biology; Knölker, H.-J., Ed.; Elsevier: Amsterdam, The Netherlands, 2020; Volume 83, pp. 113–185. [Google Scholar] [CrossRef]
- Heinrich, M.; Teoh, H.L. Galanthamine from snowdrop—The development of a modern drug against Alzheimer’s disease from local Caucasian knowledge. J. Ethnopharmacol. 2004, 92, 147–162. [Google Scholar] [CrossRef] [PubMed]
- Maelicke, A.; Samochocki, M.; Jostock, R.; Fehrenbacher, A.; Ludwig, J.; Albuquerque, E.X.; Zerlin, M. Allosteric sensitization of nicotinic receptors by galanthamine, a new treatment strategy for Alzheimer’s disease. Biol. Psychiatry 2001, 49, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Berkov, S.; Georgieva, L.; Boriana, S.; Bastida, J. Evaluation of Hippeastrum papilio (Ravenna) Van Scheepen potencial as a new industrial source of galanthamine. Ind. Crops Prod. 2022, 178, 114619. [Google Scholar] [CrossRef]
- Berkov, S.; Bastida, J.; Codina, C.; de Andrade, J.P.; Berbee, R.L.M. Extract of Hippeastrum papilio rich in galanthamine. EP2999480B1, 7 March 2013. Available online: https://patents.google.com/patent/EP2999480B1/en (accessed on 28 July 2022).
- Chang, X. Lycoris, the basis of the galanthamine industry in China. Res. Rev. J. Agric. Allied Sci. 2015, 4, 1–8. [Google Scholar]
- Nair, J.J.; Van Staden, J. Cytotoxicity studies of lycorine alkaloids of the Amaryllidaceae. Nat. Prod. Commun. 2014, 9, 1193–1210. [Google Scholar] [CrossRef] [Green Version]
- Nair, J.J.; Rárová, L.; Strnad, M.; Bastida, J.; Van Staden, J. Mechanistic insights to the cytotoxicity of Amaryllidaceae alkaloids. Nat. Prod. Commun. 2015, 10, 171–182. [Google Scholar] [CrossRef] [Green Version]
- Kaur, H.; Chahal, S.; Jha, P.; Lekhak, M.M.; Shekhawat, M.S.; Naidoo, D.; Arencibia, A.D.; Ochatt, S.J.; Kumar, V. Harnessing plant biotechnology-based strategies for in vitro galanthamine (GAL) biosynthesis: A potent drug against Alzheimer’s disease. Plant Cell. Tiss. Org. 2022, 149, 81–103. [Google Scholar] [CrossRef]
- Ortiz, J.E.; Garro, A.; Pigni, N.B.; Agüero, M.B.; Roitman, G.; Slanis, A.; Enriz, R.D.; Feresin, G.E.; Bastida, J.; Tapia, A. Cholinesterase-inhibitory effect and in silico analysis of alkaloids from bulbs of Hieronymiella species. Phytomedicine 2018, 39, 66–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šafratová, M.; Hošt’álková, A.; Hulcová, D.; Breiterová, K.; Hrabcová, V.; Machado, M.; Fontinha, D.; Prudêncio, M.; Kuneš, J.; Chlebek, J.; et al. Alkaloids from Narcissus poeticus cv. Pink Parasol of various structural types and their biological activity. Arch. Pharm. Res. 2018, 41, 208–218. [Google Scholar] [CrossRef] [PubMed]
- Hulcová, D.; Maříková, J.; Korábečný, J.; Hošťálková, A.; Jun, D.; Kuneš, J.; Chlebek, J.; Opletal, L.; De Simone, A.; Nováková, L.; et al. Amaryllidaceae alkaloids from Narcissus pseudonarcissus L. cv. Dutch Master as potential drugs in treatment of Alzheimer’s disease. Phytochemistry 2019, 165, 112055. [Google Scholar] [CrossRef] [PubMed]
- Cortes, N.; Alvarez, R.; Osorio, E.H.; Alzate, F.; Berkov, S.; Osorio, E. Alkaloid metabolite profiles by GC/MS and acetylcholinesterase inhibitory activities with binding-mode predictions of five Amaryllidaceae plants. J. Pharmaceut. Biomed. 2015, 102, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Cortes, N.; Posada-Duque, R.A.; Alvarez, R.; Alzate, F.; Berkov, S.; Cardona-Gómez, G.P.; Osorio, E. Neuroprotective activity and acetylcholinesterase inhibition of five Amaryllidaceae species: A comparative study. Life Sci. 2015, 122, 42–50. [Google Scholar] [CrossRef]
- Cortes, N.; Sierra, K.; Alzate, F.; Osorio, E.H.; Osorio, E. Alkaloids of Amaryllidaceae as inhibitors of cholinesterases (AChEs and BChEs): An integrated bioguided study. Phytochem. Anal. 2018, 29, 217–227. [Google Scholar] [CrossRef]
- Trujillo-Chacón, L.M.; Alarcón-Enos, J.E.; Céspedes-Acuña, C.L.; Bustamante, L.; Baeza, M.; López, M.G.; Fernández-Mendívil, C.; Cabezas, F.; Pastene—Navarrete, E.R. Neuroprotective activity of isoquinoline alkaloids from Chilean Amaryllidaceae plants against oxidative stress-induced cytotoxicity on human neuroblastoma SH-SY5Y cells and mouse hippocampal slice culture. Food Chem. Toxicol. 2019, 132, 110665. [Google Scholar] [CrossRef]
- Moreno, R.; Tallini, L.R.; Salazar, C.; Osorio, E.H.; Montero, E.; Bastida, J.; Oleas, N.H.; León, K.A. Chemical profiling and cholinesrerase inhibitory activity of five Phaedranassa Herb. (Amaryllidaceae) species from Ecuador. Molecules 2020, 25, 2092. [Google Scholar] [CrossRef]
- Acosta, K.L.; Inca, A.; Tallini, L.R.; Osorio, E.H.; Robles, J.; Bastida, J.; Oleas, N.H. Alkaloids of Phaedranassa dubia (Kunth) J.F. Macbr. and Phaedranassa brevifolia Meerow (Amaryllidaceae) from Ecuador and its cholinesterase-inhibitory activity. S. Afr. J. Bot. 2021, 136, 91–99. [Google Scholar] [CrossRef]
- Tallini, L.R.; Carrasco, A.; Acosta, K.L.; Vinueza, D.; Bastida, J.; Oleas, N.H. Alkaloid profiling and cholinesterase inhibitory potential of Crinum x amabile Donn. (Amaryllidaceae) collected in Ecuador. Plants 2021, 10, 2686. [Google Scholar] [CrossRef]
- Soto-Vásquez, M.R.; Rodríguez-Muñoz, C.A.; Tallini, L.R.; Bastida, J. Alkaloid composition and biological activities of the Amaryllidaceae species Ismene amancaes (Ker Gawl.) Herb. Plants 2022, 11, 1906. [Google Scholar] [CrossRef] [PubMed]
- Tallini, L.R.; Bastida, J.; Cortes, N.; Osorio, E.H.; Theoduloz, C.; Schmeda-Hirschmann, G. Cholinesterase inhibition activity, alkaloid profiling, and molecular docking of Chilean Rhodophiala (Amaryllidaceae). Molecules 2018, 23, 1532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moraga-Nicolás, F.; Jara, C.; Godoy, R.; Iturriaga-Vásquez, P.; Venthur, H.; Quiroz, A.; Becerra, J.; Mutis, A.; Hormazábal, E. Rhodolirium andicola: A new renewable source of alkaloids with acetylcholinesterase inhibitory activity, a study from nature to molecular docking. Rev. Bras. Farmacogn. 2018, 28, 34–43. [Google Scholar] [CrossRef]
- Fernández-Galleguillos, C.; Romero-Parra, J.; Puerta, A.; Padrón, J.M.; Simirgiotis, M.J. Alkaloid profiling, anti-enzymatic and antiproliferative activity of the endemic Chilean Amaryllidaceae Phycella cyrtanthoides. Metabolites 2022, 12, 188. [Google Scholar] [CrossRef] [PubMed]
- Del Rojas-Vera, J.C.; Buitrago-Díaz, A.A.; Possamai, L.M.; Timmers, L.F.S.M.; Tallini, L.R.; Bastida, J. Alkaloid profile and cholinesterase inhibition activity of five species of Amaryllidaceae family collected from Mérida state-Venezuela. S. Afri. J. Bot. 2021, 136, 126–136. [Google Scholar] [CrossRef]
- De Andrade, J.P.; Giordani, R.B.; Torras-Claveria, L.; Pigni, N.B.; Berkov, S.; Font-Bardia, M.; Calvet, T.; Konrath, E.; Bueno, K.; Sachett, L.G.; et al. The Brazilian Amaryllidaceae as a source of aceylcholinesterase inhibitoy alkaloids. Phytochem. Rev. 2016, 15, 147–160. [Google Scholar] [CrossRef]
- Gasca, C.A.; Moreira, N.C.S.; de Almeida, F.C.; Gomes, J.V.D.; Castillo, W.O.; Fagg, C.W.; Magalhaes, P.O.; Fonseca-Bazzo, Y.M.; Sakamoo-Hojo, E.; de Medeiros, Y.K.; et al. Aceylcholinesterase inhibitory activity, anti-inflammaory, and neuroprotective potential of Hippeastrum psittacinum (Ker Gawl.) Herb (Amaryllidaceae). Food Chem. Toxicol. 2000, 145, 111703. [Google Scholar] [CrossRef]
- Ortiz, J.E.; Pigni, N.B.; Andujar, S.A.; Roitman, G.; Suvire, F.D.; Enriz, R.D.; Tapia, A.; Basida, J.; Feresin, G.E. Alkaloids from Hippeastrum argentinum and their cholinesterase-inhibitory activities: An in vitro and in silico study. J. Nat. Prod. 2016, 79, 1241–1248. [Google Scholar] [CrossRef]
- Zaragoza-Puchol, D.; Ortiz, J.E.; Orden, A.A.; Sanchez, M.; Palermo, J.; Tapia, A.; Bastida, J.; Feresin, G.E. Alkaloids analysis of Habranthus cardanasianus (Amaryllidaceae), anti-cholinesterase activity and biomass production by propagation strategies. Molecules 2021, 26, 192. [Google Scholar] [CrossRef]
- Ortiz, J.E.; Berkov, S.; Pigni, N.B.; Theoduloz, C.; Roitman, G.; Tapia, A.; Bastida, J.; Feresin, G.E. Wild Argentinian Amaryllidaceae, a new renewable source of the acetylcholinesterase inhibitor galanthamine and other alkaloids. Molecules 2012, 17, 13473–13482. [Google Scholar] [CrossRef] [Green Version]
- García, N.; Meerow, A.W.; Arroyo-Leuenberger, S.; Oliveira, R.S.; Dutilh, J.H.; Soltis, P.S.; Judd, W.S. Generic classification of Amaryllidaceae tribe Hippeastreae. Taxon 2019, 68, 481–498. [Google Scholar] [CrossRef]
- Cheung, J.; Rudolph, M.J.; Burshteyn, F.; Cassidy, M.S.; Gary, E.N.; Love, J.; Franklin, M.C.; Height, J.J. Structures of human acetylcholinesterase in complex with pharmacologically important ligands. J. Med. Chem. 2012, 55, 10282–10286. [Google Scholar] [CrossRef] [PubMed]
- Cheung, J.; Gary, E.N.; Shiomi, K.; Rosenberry, T.L. Structures of human acetylcholinesterase bound to dihydrotanshinone I and territrem B show peripheral site flexibility. ACS Med. Chem. Lett. 2013, 4, 1091–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sierra, K.; de Andrade, J.P.; Tallini, L.R.; Osorio, E.H.; Yañéz, O.; Osorio, M.I.; Oleas, N.H.; García-Beltrán, O.; de Borges, W.S.; Bastida, J.; et al. In vitro and in silico analysis of galanthine from Zephyranthes carinata as an inhibitor of acetylcholinesterase. Biomed. Pharmacother. 2022, 150, 113016. [Google Scholar] [CrossRef] [PubMed]
- Torras-Claveria, L.; Berkov, S.; Codina, C.; Viladomat, F.; Bastida, J. Daffodils as potential crops of galanthamine. Assessment of more than 100 ornamental varieties for their alkaloid content and acetylcholinesterase inhibitory activity. Ind. Crops Prod. 2013, 43, 237–244. [Google Scholar] [CrossRef]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [Green Version]
- Schrödinger Release 2022-3: Maestro; Schrödinger, Inc.: New York, NY, USA, 2021.
- Morris, G.M.; Goodsell, D.S.; Halliday, R.S.; Huey, R.; Hart, W.E.; Belew, R.K.; Olson, A.J. Automated docking using a Lamarckinan genetic algorithm and an empirical binding free energy function. J. Comput. Chem. 1999, 19, 1639–1662. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Alkaloid | [M]+ | m/z | RI | A 1 | A 2 | B 1 | B 2 | C 1 | C 2 |
|---|---|---|---|---|---|---|---|---|---|
| Lycorine-type | 317.5 | 314.5 | |||||||
| 1-O-acetylpluviine (1) | 329 (80) | 268 (85), 242 (100) | 2598.0 | - | - | 10.1 | 0.1 | - | - |
| pluviine (2) | 287 (78) | 286 (52), 268 (55), 243 (61), 242 (100) | 2608.2 | 16.1 | 1.4 | 69.2 | 11.9 | - | - |
| assoanine (3) | 267 (57) | 266 (100), 250 (28), 222 (12), 180 (13) | 2622.3 | 21.6 | 3.3 | 24.0 | 2.9 | - | - |
| norpluviine (4) | 273 (80) | 254 (60), 228 (100) | 2635.7 | - | - | 10.3 | 0.3 | - | - |
| 1-O-acetylnorpluviine (5) | 315 (80) | 254 (90), 228 (100) | 2641.5 | - | - | 16.0 | 2.4 | - | - |
| kirkine (6) | 273 (<1) | 253 (55), 252 (100), 237 (21), 209 (22) | 2642.2 | 13.9 | 1.4 | - | - | - | - |
| 1-O-acetyl-9-O-methylpseudolycorine (7) | 345 (30) | 284 (25), 242 (100) | 2769.3 | - | - | 10.5 | 0.3 | - | - |
| galanthine (8) | 317 (20) | 298 (10), 268 (15), 242 (100), 228 (5) | 2775.9 | 21.6 | 3.3 | - | - | - | - |
| lycorine (9) | 287 (30) | 268 (27), 250 (15), 226 (100), 147 (15) | 2789.3 | - | - | 10.1 | 0.1 | - | - |
| 9-O-methylpseudolycorine (10) | 303 (33) | 302 (22), 284 (14), 243 (78), 242 (100) | 2830.1 | 11.4 | 1.1 | 17.9 | 2.4 | - | - |
| sternbergine (11) | 331 (41) | 270 (32), 252 (14), 229 (72), 228 (100) | 2844.1 | 25.8 | 17.4 | 10.8 | 0.6 | - | - |
| 2-O-acetyl-9-O-methylpseudolycorine (12) | 345 (30) | 284 (100), 268 (40), 242 (40) | 2907.3 | 168.9 | 32.5 | 135.6 | 16.6 | - | - |
| 2-O-acetylpseudolycorine (13) | 331 (30) | 270 (100), 254 (75), 228 (80) | 2945.1 | 38.2 | 15.4 | - | - | - | - |
| Galanthamine-type | 10.0 | 10.8 | 278.7 | ||||||
| galanthamine (14) | 287 (94) | 286 (100), 270 (25), 244 (42), 216 (49) | 2437.0 | - | - | - | - | 103.6 | 48.3 |
| lycoramine (15) | 289 (78) | 288 (100), 232 (14), 202 (22), 187 (18) | 2459.4 | 10.0 | 0.1 | 10.8 | 0.4 | 73.1 | 10.3 |
| sanguinine (16) | 273 (100) | 272 (81), 256 (23), 230 (16), 202 (44) | 2476.2 | - | - | - | - | 21.7 | 5.3 |
| O-demethyllycoramine (17) | 275 (67) | 274 (100), 218 (8), 174 (13), 173 (17) | 2487.6 | - | - | - | - | 23.4 | 5.9 |
| lycoraminone (18) | 287 (68) | 286 (100), 244 (5), 218 (17), 202 (23) | 2491.6 | - | - | - | - | 10.3 | 0.3 |
| narwedine (19) | 285 (86) | 284 (100), 216 (25), 199 (24), 174 (43) | 2517.5 | - | - | - | - | 17.3 | 1.8 |
| 3-O-acetylgalanthamine (20) | 329 (34) | 328 (31), 270 (100), 216 (31), 165 (17) | 2577.2 | - | - | - | - | 13.0 | 0.9 |
| 3-O-acetylsanguinine (21) | 315 (46) | 256 (100), 255 (59), 254 (40), 212 (29) | 2584.6 | - | - | - | - | 16.3 | 1.4 |
| Homolycorine-type | 57.0 | 42.0 | |||||||
| nerinine (22) | 347 (<1) | 110 (8), 109 (100), 108 (18) | 2511.4 | 11.0 | 0.2 | 18.8 | 1.6 | - | - |
| homolycorine (23) | 315 (<1) | 110 (11), 109 (100), 108 (30) | 2785.4 | 19.2 | 2.5 | 10.0 | 0.1 | - | - |
| 8-O-demethylhomolycorine (24) | 301 (<1) | 110 (23), 109 (100), 108 (53) | 2847.6 | 26.8 | 4.1 | 13.2 | 1.3 | - | - |
| Haemanthamine -type | 11.0 | ||||||||
| 8-O-demethylmaritidine (25) | 273 (100) | 230 (24), 202 (27), 201 (93), 189 (60) | 2549.8 | - | - | 11.0 | 0.4 | - | - |
| Pretazettine-type | 11.7 | 9.9 | |||||||
| O-methyltazettine (26) | 345 (30) | 330 (30), 314 (25), 261 (100), 239 (25) | 2643.2 | - | - | - | - | 9.9 | 0.2 |
| tazettine (27) | 331 (24) | 316 (13), 298 (20), 247 (100), 70 (26) | 2686.1 | - | - | 11.7 | 1.2 | - | - |
| Unidentified | 24.8 | 11.2 | |||||||
| UI 1 (HLY type) (28) | 329 (<1) | 221 (<1), 109 (100) | 2510.8 | - | - | 11.2 | 1.1 | - | - |
| UI 2 (HLY type) (29) | 330 (<1) | 221 (<1), 109 (100) | 2555.9 | 13.4 | 1.2 | - | - | - | - |
| UI 3 (30) | 325 (40) | 282 (100), 266 (10), 139 (60) | 2989.5 | 11.4 | 0.3 | - | - | - | - |
| Total: | 409.3 | 401.2 | 288.6 |
| alkaloid | 4EY5 | 4EY6 | 4EY7 | 4M0E | 4M0F |
|---|---|---|---|---|---|
| 3-O-acetylgalanthamine (20) | −9.08 | −9.77 | −11.25 | −8.57 | −9.93 |
| 3-O-acetylsanguinine (21) | −8.75 | −9.76 | −10.55 | −8.42 | −10.11 |
| narwedine (19) | −9.15 | −9.70 | −10.41 | −8.69 | −9.72 |
| lycoraminone (18) | −9.70 | −9.48 | −9.37 | −9.10 | −9.25 |
| lycoramine (15) | −8.84 | −9.08 | −8.87 | −8.64 | −8.41 |
| O-demethyllycoramine (17) | −8.74 | −9.08 | −8.91 | −8.66 | −8.40 |
| sanguinine (16) | −8.13 | −8.54 | −9.14 | −8.50 | −9.12 |
| galanthamine (14) | −8.59 | −8.75 | −9.83 | −7.90 | −8.74 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tallini, L.R.; Osorio, E.H.; Berkov, S.; Torras-Claveria, L.; Rodríguez-Escobar, M.L.; Viladomat, F.; Meerow, A.W.; Bastida, J. Chemical Survey of Three Species of the Genus Rauhia Traub (Amaryllidaceae). Plants 2022, 11, 3549. https://doi.org/10.3390/plants11243549
Tallini LR, Osorio EH, Berkov S, Torras-Claveria L, Rodríguez-Escobar ML, Viladomat F, Meerow AW, Bastida J. Chemical Survey of Three Species of the Genus Rauhia Traub (Amaryllidaceae). Plants. 2022; 11(24):3549. https://doi.org/10.3390/plants11243549
Chicago/Turabian StyleTallini, Luciana R., Edison H. Osorio, Strahil Berkov, Laura Torras-Claveria, María L. Rodríguez-Escobar, Francesc Viladomat, Alan W. Meerow, and Jaume Bastida. 2022. "Chemical Survey of Three Species of the Genus Rauhia Traub (Amaryllidaceae)" Plants 11, no. 24: 3549. https://doi.org/10.3390/plants11243549
APA StyleTallini, L. R., Osorio, E. H., Berkov, S., Torras-Claveria, L., Rodríguez-Escobar, M. L., Viladomat, F., Meerow, A. W., & Bastida, J. (2022). Chemical Survey of Three Species of the Genus Rauhia Traub (Amaryllidaceae). Plants, 11(24), 3549. https://doi.org/10.3390/plants11243549