
Phenotypic Variation in European Wild Pear (Pyrus pyraster (L.) Burgsd.) Populations in the North-Western Part of the Balkan Peninsula


 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Study Area
2.2. Sampling Method
2.3. Studied Phenotypic Traits
2.4. Environmental Data
2.5. Population Phenotypic Diversity
2.6. Population Structure
2.7. Correlation between Environmental, Geographic and Phenotypic Data
3. Results
3.1. Climate Differences among Sampling Sites
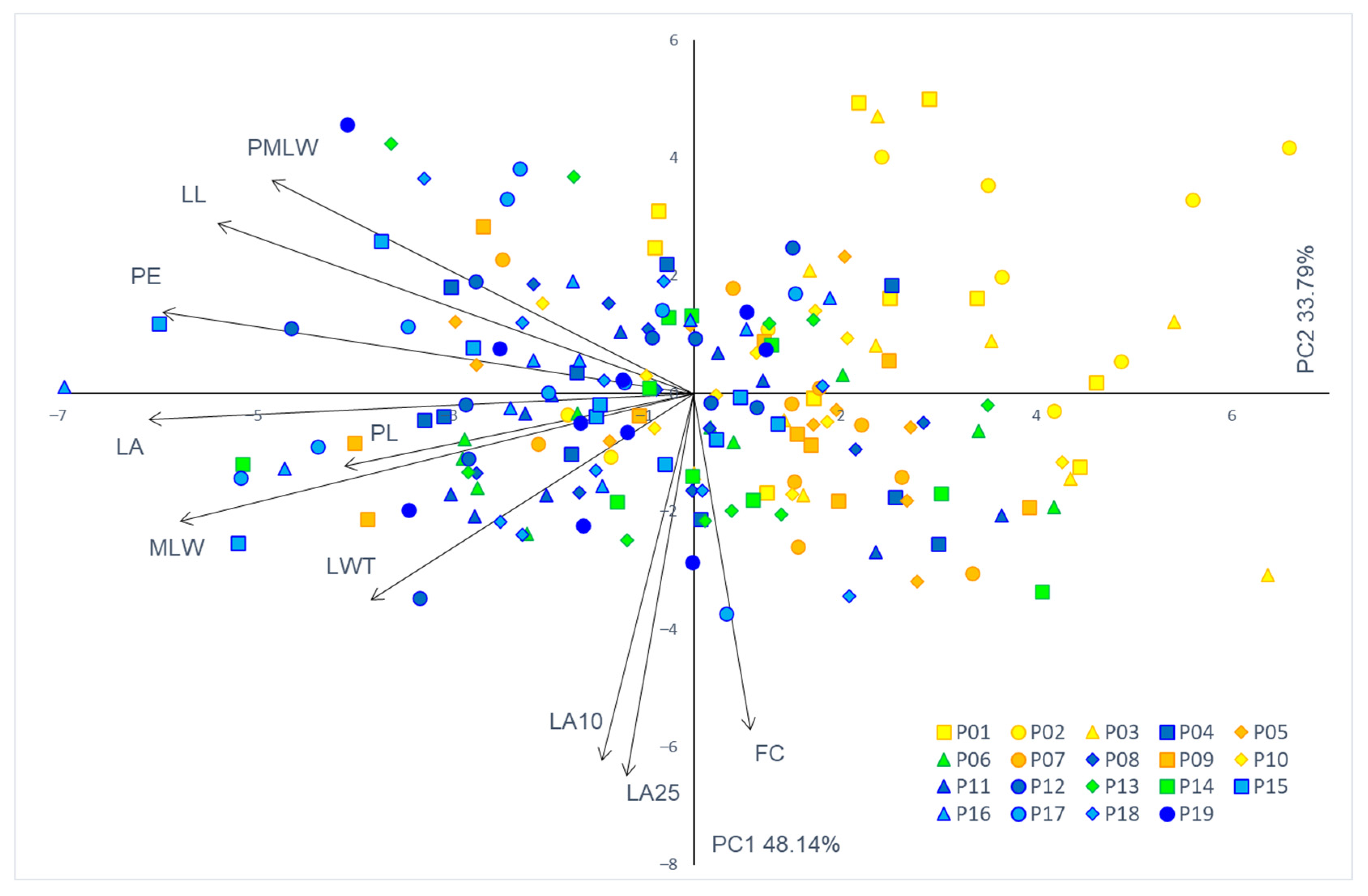
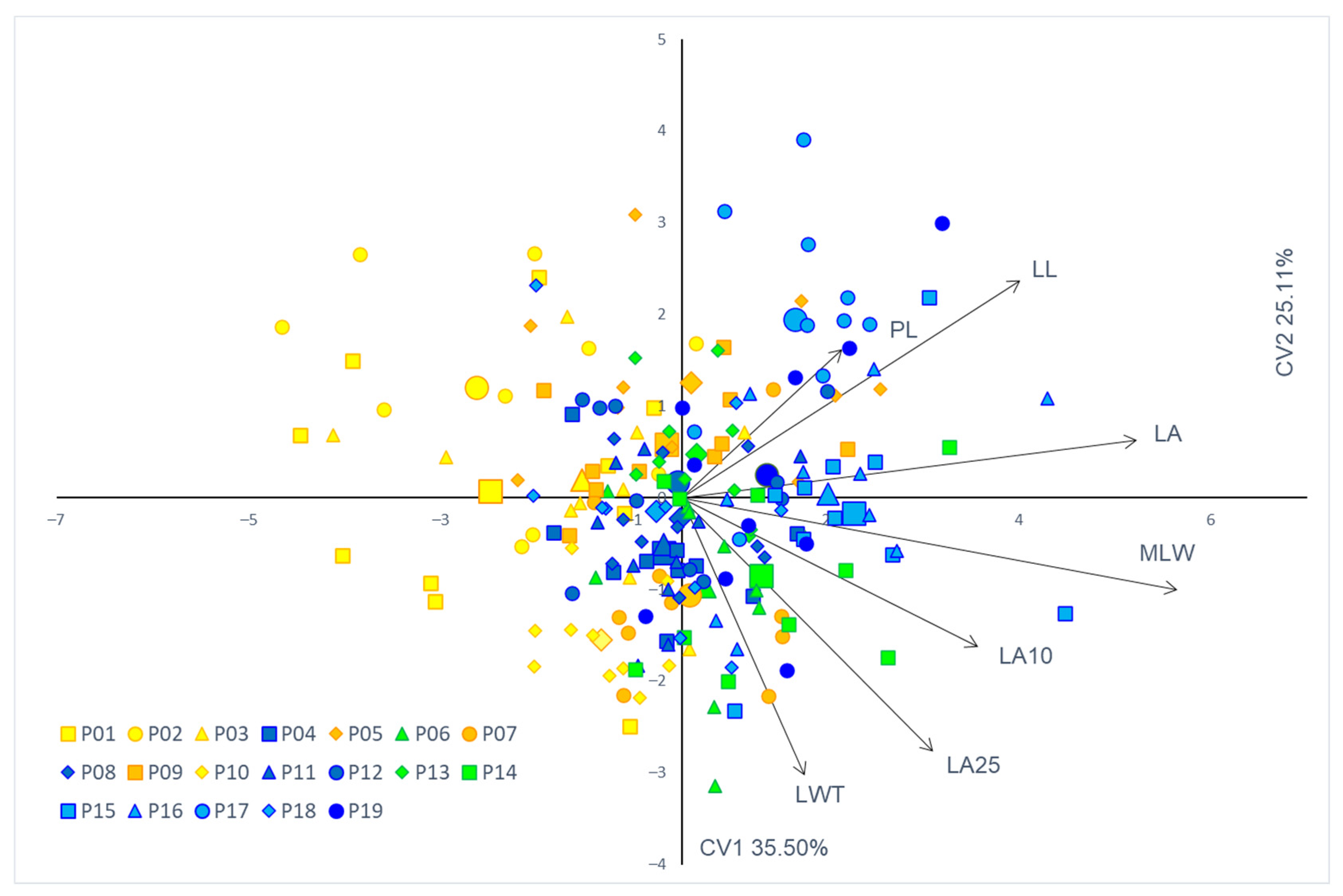
3.2. Phenotypic Traits and Populations’ Diversity
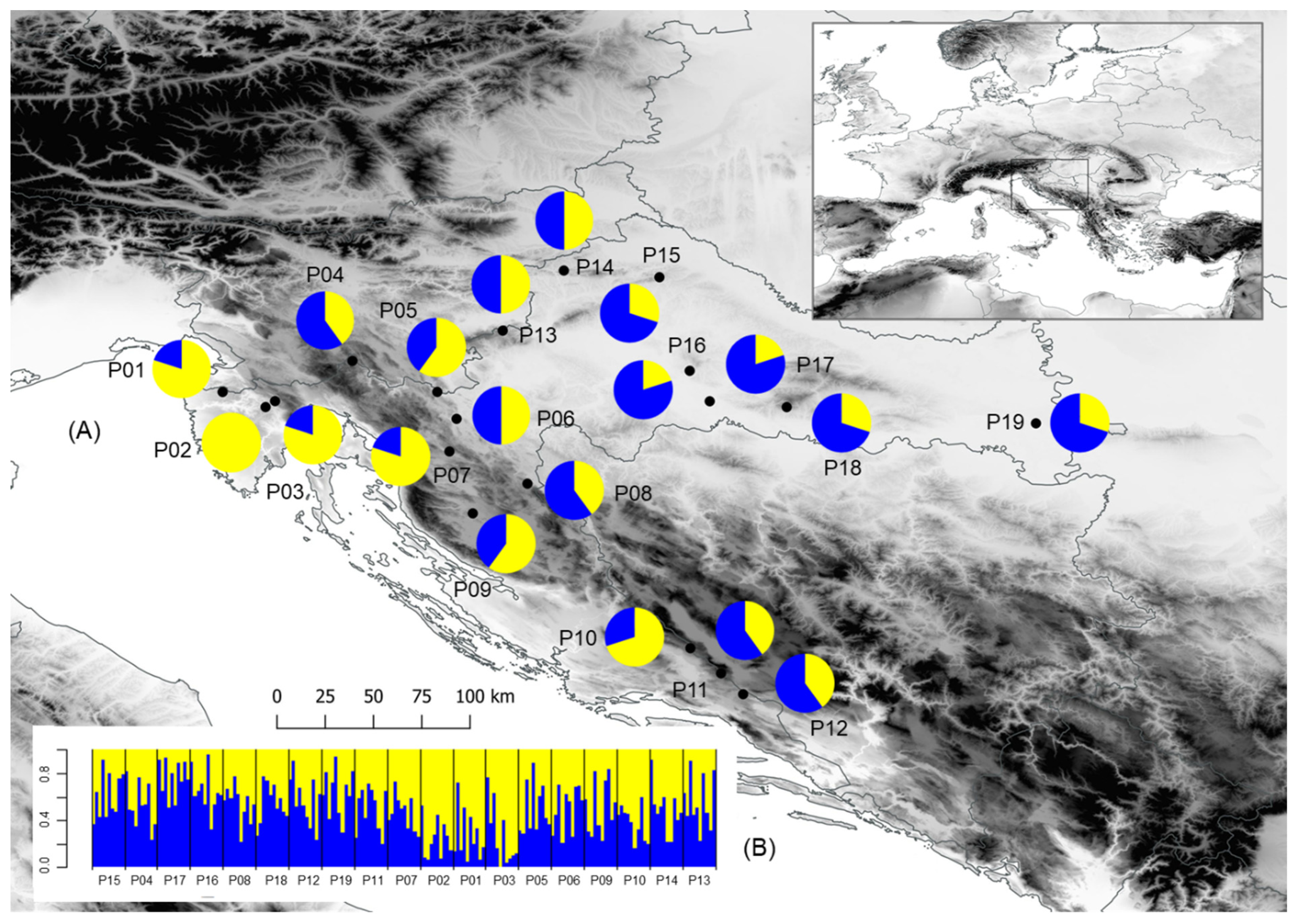
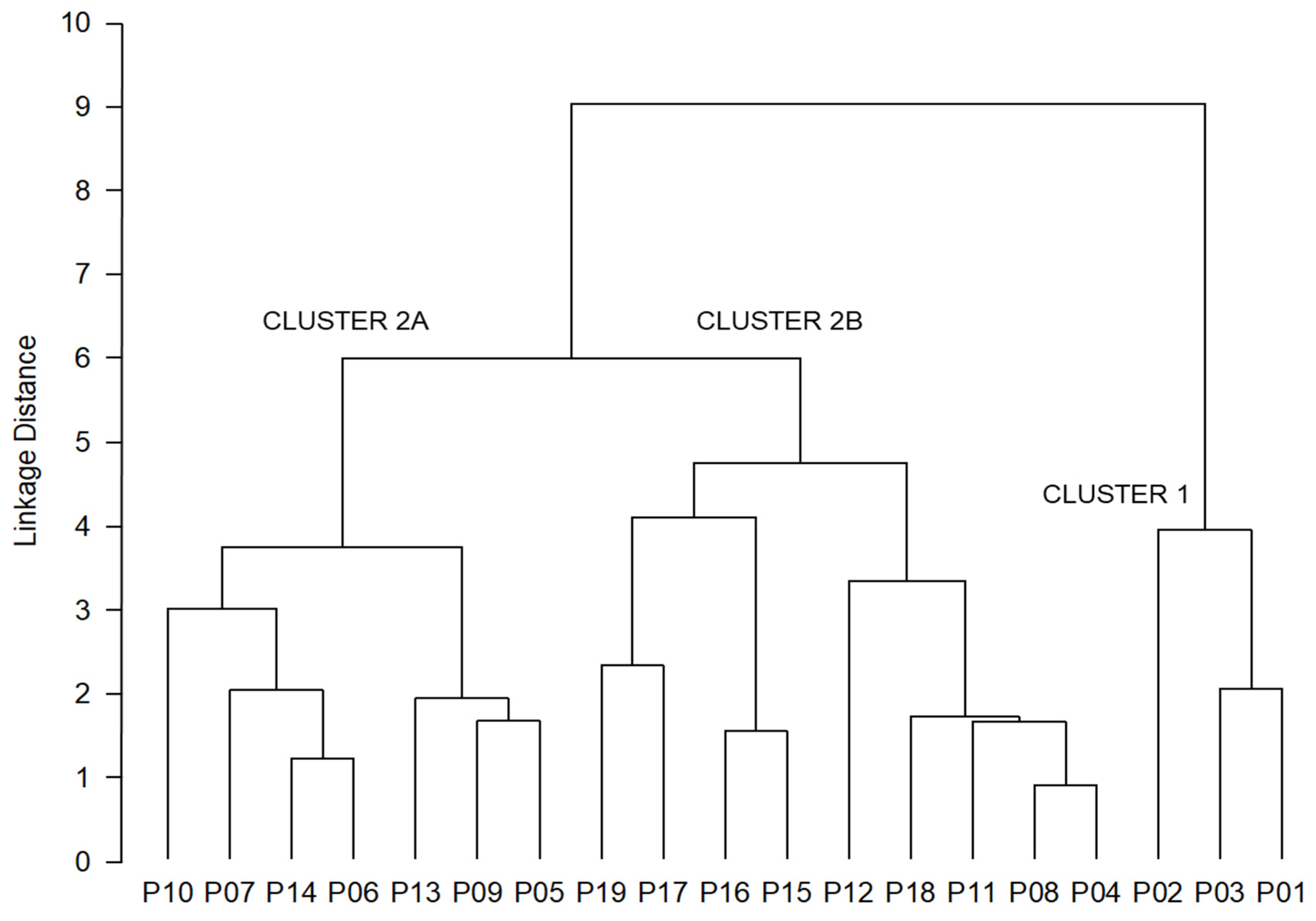
3.3. Population Structure
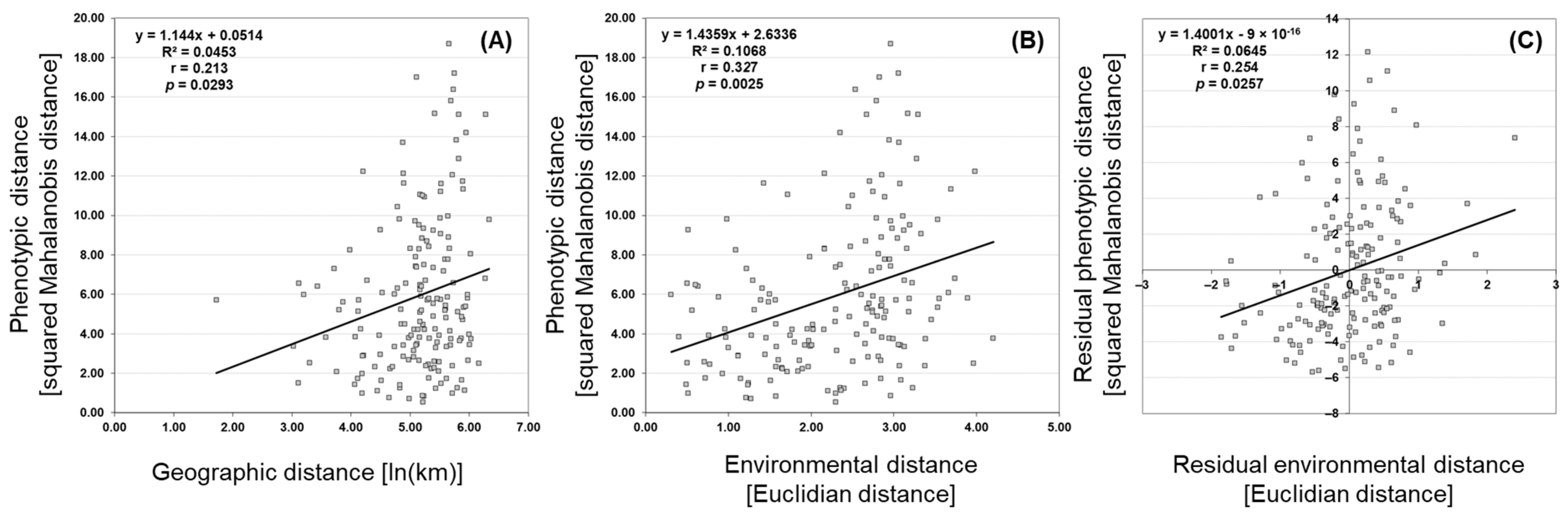
3.4. Isolation by Distance (IBD) and Isolation by Environment (IBE)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Matesanz, S.; Gianoli, E.; Valladares, F. Global change and the evolution of phenotypic plasticity in plants. Ann. N. Y. Acad. Sci. 2010, 1206, 35–55. [Google Scholar] [CrossRef] [PubMed]
- Palacio-López, K.; Beckage, B.; Scheiner, S.; Molofsky, J. The ubiquity of phenotypic plasticity in plants: A synthesis. Ecol. Evol. 2015, 5, 3389–3400. [Google Scholar] [CrossRef] [PubMed]
- Sultan, S.E. Phenotypic plasticity and plant adaptation. Acta Bot. Neerl. 1995, 44, 363–383. [Google Scholar] [CrossRef]
- Vázquez, D.P.; Gianoli, E.; Morris, W.F.; Bozinovic, F. Ecological and evolutionary impacts of changing climatic variability. Biol. Rev. 2015, 92, 22–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlichting, C.D.; Smith, H. Phenotypic plasticity: Linking molecular mechanisms with evolutionary outcomes. Evol. Ecol. 2002, 16, 189–211. [Google Scholar] [CrossRef]
- Stotz, G.C.; Salgado-Luarte, C.; Escobedo, V.M.; Valladares, F.; Gianoli, E. Global trends in phenotypic plasticity of plants. Ecol. Lett. 2021, 24, 2267–2281. [Google Scholar] [CrossRef] [PubMed]
- Sultan, S.E. Phenotypic plasticity for plant development, function and life history. Trends Plant Sci. 2000, 5, 537–542. [Google Scholar] [CrossRef]
- Valladares, F.; Gianoli, E.; Gómez, J.M. Ecological limits to plant phenotypic plasticity. New Phytol. 2007, 176, 749–763. [Google Scholar] [CrossRef]
- Chitwood, D.H.; Sinha, N.R. Evolutionary and environmental forces sculpting leaf development. Curr. Biol. 2016, 26, R297–R306. [Google Scholar] [CrossRef] [Green Version]
- Tsukaya, H. Leaf shape diversity with an emphasis on leaf contour variation, developmental background, and adaptation. Semin. Cell Dev. Biol. 2018, 79, 48–57. [Google Scholar] [CrossRef]
- Atkin, O.K.; Loveys, B.R.; Atkinson, L.J.; Pons, T.L. Phenotypic plasticity and growth temperature: Understanding interspecific variability. J. Exp. Bot. 2005, 57, 267–281. [Google Scholar] [CrossRef] [PubMed]
- Royer, D.L.; Meyerson, L.A.; Robertson, K.M.; Adams, J.M. Phenotypic plasticity of leaf shape along a temperature gradient in Acer rubrum. PLoS ONE 2009, 4, e7653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosbakh, S.; Römermann, C.; Poschlod, P. Specific leaf area correlates with temperature: New evidence of trait variation at the population, species and community levels. Alp. Bot. 2015, 125, 79–86. [Google Scholar] [CrossRef]
- McKee, M.L.; Royer, D.L.; Poulos, H.M. Experimental evidence for species-dependent responses in leaf shape to temperature: Implications for paleoclimate inference. PLoS ONE 2019, 14, e0218884. [Google Scholar] [CrossRef] [PubMed]
- McDonald, P.G.; Fonseca, C.R.; Overton, J.M.; Westoby, M. Leaf-size divergence along rainfall and soil-nutrient gradients: Is the method of size reduction common among clades? Funct. Ecol. 2003, 17, 50–57. [Google Scholar] [CrossRef] [Green Version]
- Leuschner, C.; Voß, S.; Foetzki, A.; Clases, Y. Variation in leaf area index and stand leaf mass of European beech across gradients of soil acidity and precipitation. Plant Ecol. 2006, 186, 247–258. [Google Scholar] [CrossRef]
- Meier, I.C.; Leuschner, C. Leaf size and leaf area index in Fagus sylvatica forests: Competing effects of precipitation, temperature, and nitrogen availability. Ecosystems 2008, 11, 655–669. [Google Scholar] [CrossRef] [Green Version]
- Grassi, G.; Bagnaresi, U. Foliar morphological and physiological plasticity in Picea abies and Abies alba saplings along a natural light gradient. Tree Physiol. 2001, 21, 959–967. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.; Guo, W.; Xu, W.; Wei, Y.; Wang, R. Leaf morphology correlates with water and light availability: What consequences for simple and compound leaves? Prog. Nat. Sci. 2009, 19, 1789–1798. [Google Scholar] [CrossRef]
- Coble, A.P.; Cavaleri, M.A. Light drives vertical gradients of leaf morphology in a sugar maple (Acer saccharum) forest. Tree Physiol. 2014, 34, 146–158. [Google Scholar] [CrossRef]
- Pearcy, R.W.; Muraoka, H.; Valladares, F. Crown architecture in sun and shade environments: Assessing function and trade-offs with a three-dimensional simulation model. New Phytol. 2005, 166, 791–800. [Google Scholar] [CrossRef] [PubMed]
- Niinemets, Ü.; Valladares, F. Tolerance to shade, drought, and waterlogging of temperate northern hemisphere trees and shrubs. Ecol. Monogr. 2006, 76, 521–547. [Google Scholar] [CrossRef]
- Fritz, M.A.; Rosa, S.; Sicard, A. Mechanisms underlying the environmentally induced plasticity of leaf morphology. Front. Genet. 2018, 9, 478. [Google Scholar] [CrossRef]
- Miljković, D.; Čortan, D. Morphometric and morphological analysis of Populus nigra L. leaves in flooded regions. Šumar. List. 2020, 144, 139–147. [Google Scholar] [CrossRef]
- Hegi, G. Illustrierte Flora von Mitteleuropa; Blackwell Wissenschaft-Verlag: Berlin, Germany, 1995; pp. 284–286. [Google Scholar]
- Cedro, A.; Antkowiak, W. Dendroclimatological analysis of wild pear Pyrus pyraster (L.) Burgsd. from Biedrusko Military area (West Poland)—Preliminary study. Geochronometria 2016, 43, 18–23. [Google Scholar] [CrossRef] [Green Version]
- Paganová, V. Taxonomic reliability of leaf and fruit morphological characteristics of the Pyrus L. taxa in Slovakia. Hortic. Sci. 2011, 30, 98–107. [Google Scholar] [CrossRef] [Green Version]
- Idžojtić, M. Dendrology: Cones, Flowers, Fruits and Seeds; Elsevier-Academic Press: London/San Diego/Cambridge/Oxford, UK, 2019; p. 800. [Google Scholar]
- Fedriani, J.M.; Wiegand, T.; Delibes, M. Spatial pattern of adult trees and the mammal-generated seed rain in the Iberian pear. Ecography 2010, 33, 545–555. [Google Scholar] [CrossRef]
- Reim, S.; Lochschmidt, F.; Proft, A.; Wolf, H.; Wolf, H. Species delimitation, genetic diversity and structure of the European indigenous wild pear (Pyrus pyraster) in Saxony, Germany. Genet. Resour. Crop. Evol. 2016, 64, 1075–1085. [Google Scholar] [CrossRef]
- Kleinschmit, J.; Soppa, B.; Wagner, I.; Fellenberg, U.; Schmidt, J.; Brötje, H.; Schüte, G.; Meier-Dinkel, A. Die Wildbirne–Baum des Jahres 1998. Forst. Und Holz. 1998, 53, 35–39. [Google Scholar]
- Kleinschmit, J.; Stephan, R.; Wagner, I. Conservation of genetic resources of wild fruit species (Prunus avium, Malus sylvestris and Pyrus pyraster). In Noble Hardwoods Network Report of the Second Meeting; Turok, J., Collin, E., Demesure, B., Erikson, G., Kleinschmit, J., Rusanen, M., Stephan, R., Eds.; International Plant Genetic Resources Institute: Rome, Italy, 1998; pp. 51–57. [Google Scholar]
- Wolko, Ł.; Bocianowski, J.; Antkowiak, W.; Słomski, R. Genetic diversity and population structure of wild pear (Pyrus pyraster (L.) Burgsd.) in Poland. Open Life Sci. 2014, 10, 19–29. [Google Scholar] [CrossRef] [Green Version]
- Terpó, A. Comprehensive survey of taxonomy of species Pyrus. Acta Hortic. 1984, 161, 117–132. [Google Scholar] [CrossRef]
- Terpó, A. Pyrus taxa in Hungary, and their practical importance. Thaiszia 1992, 2, 41–57. [Google Scholar]
- Paganová, V. The occurrence and morphological characteristics of the wild pear lower taxa in Slovakia. Hortic. Sci. 2009, 36, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Rouhan, G.; Gaudeul, M. Plant taxonomy: A historical perspective, current challenges, and perspectives. In Molecular Plant Taxonomy. Methods in Molecular Biology (Methods and Protocols); Besse, P., Ed.; Humana Press: Totowa, NJ, USA, 2014; Volume 1115, p. 37. [Google Scholar] [CrossRef]
- Hofmann, H. Zur Verbreitung und Ökologie der Wildbirne (Pyrus communis L.) in Süd-Niedersachen und Nordhessen sowie ihrer Abgrenzung von verwilderten Kulturbirnen (Pyrus domestica MED.). Mitt. Dtsch. Dendrol. Ges. 1993, 81, 27–69. [Google Scholar]
- Wagner, I. Identifikation von Wildapfel (Malus sylvestris (L.) Mill.) und Wildbirne (Pyrus pyraster (L.) Burgsd.) Voraussetzung zur Generhaltung des einheimischen Wildobstes. Forstarchiv 1995, 66, 39–47. [Google Scholar]
- Wagner, I. Zusammenstellung morphologischer Merkmale und ihrer Ausprägungen zur Unterscheidung von Wild-und Kulturformen der Apfel und Birnbaumes. Mitt. Dtsch. Dendrol. Ges. 1996, 82, 87–108. [Google Scholar]
- WinFolia TM, Version PRO 2005b; Regent Instruments Inc.: Quebec, ON, Canada, 2001.
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- O’Donnell, M.S.; Ignizio, D.A. Bioclimatic predictors for supporting ecological applications in the conterminous United States. In Geological Survey Data Series 691; United States Geological Survey: Reston, VA, USA, 2012; p. 10. [Google Scholar]
- Sokal, R.R.; Rohlf, F.J. Biometry: The Principles and Practice of Statistics in Biological Research, 4th ed.; W.H. Freeman and Co.: New York, NY, USA, 2012; p. 937. [Google Scholar]
- Douaihy, B.; Sobierajska, K.; Jasińska, A.K.; Boratyńska, K.; Ok, T.; Romo, A.; Machon, N.; Didukh, Y.; Dagher-Kharrat, M.B.; Boratyński, A. Morphological versus molecular markers to describe variability in Juniperus excelsa subsp. excelsa (Cupressaceae). AoB PLANTS 2012, 2012, pls013. [Google Scholar] [CrossRef] [Green Version]
- Poljak, I.; Idžojtić, M.; Šapić, I.; Korijan, P.; Vukelić, J. Diversity and structure of Croatian continental and Alpine-Dinaric populations of grey alder (Alnus incana/L./Moench subsp. incana); Isolation by distance and environment explains phenotypic divergence. Šumar. List 2018, 142, 19–31. [Google Scholar] [CrossRef]
- Statistica (Data Analysis Software System), version 13; TIBCO Software Inc.: Palo Alto, CA, USA, 2018. Available online: http://www.statsoft.com (accessed on 15 November 2021).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016; Available online: https://www.R-project.org/ (accessed on 17 July 2019).
- Sokal, R.R. Testing statistical significance of geographic variation patterns. Syst. Zool. 1979, 28, 227–232. [Google Scholar] [CrossRef]
- Sokal, R.R.; Rohlf, F.J. Biometry–The Principles and Practice of Statistics in Biological Research, 3rd ed.; W.H. Freeman and Co.: New York, NY, USA, 1995. [Google Scholar]
- Rohlf, F.J. NTSYS-pc: Numerical Taxonomy and Multivariate Analysis System, version 2.2; Applied Biostatistics Inc.: New York, NY, USA, 2009; p. 44. [Google Scholar]
- Boratyńska, K. 1990 Systematics and geographic distribution: European pear Pyrus communis L. In Wild Fruit Trees. Bird Cherry Cerasus avium (L.) Moench, Wild Apple Malus sylvestris (L.) Miller, Common pear Pyrus communis L.; Białobok, S., Ed.; Nasze Drzewa Leśne, Monografie Popularnonaukowe: Poznań, Poland, 1990; pp. 81–91. [Google Scholar]
- Idžojtić, M. Dendrologija–List; Faculty of Forestry and Wood Technology, University of Zagreb: Zagreb, Croatia, 2009; p. 635. [Google Scholar]
- Korotkova, N.; Parolly, G.; Khachatryan, A.; Ghulikyan, L.; Sargsyan, H.; Akopian, J.; Borsch, T.; Gruenstaeudl, M. Towards resolving the evolutionary history of Caucasian pears (Pyrus, Rosaceae)-Phylogenetic relationships, divergence times and leaf trait evolution. J. Syst. Evol. 2017, 56, 35–47. [Google Scholar] [CrossRef]
- Fedorov, A.A. Caucasian Representatives of the Genus Pyrus L. (Morphological-Geographical Investigation); The Armenian Branch of the USSR Academy of Sciences: Yerevan, Armenia, 1943. [Google Scholar]
- Krüssmann, G. Handbuch der Laubgehölze; Paul Parkey in Berlin und Hamburg, Verlag für Landwirtschaft, Veterinärmedizin, Gartenbau und Forstwesen: Berlin, Germany, 1976; Volume 3. [Google Scholar]
- Nicotra, A.B.; Leigh, A.; Boyce, C.K.; Jones, C.S.; Niklas, K.J.; Royer, D.L.; Tsukaya, H. The evolution and functional significance of leaf shape in the angiosperms. Funct. Plant Biol. 2011, 38, 535–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radersma, R.; Noble, D.W.; Uller, T. Plasticity leaves a phenotypic signature during local adaptation. Evol. Lett. 2020, 4, 360–370. [Google Scholar] [CrossRef] [PubMed]
- Kidner, C.A.; Umbreen, S. Why is leaf shape so variable? Int. J. Plant Dev. Biol. 2010, 4, 64–75. [Google Scholar]
- Poljak, I. Morfološka i Genetska Raznolikost Populacija i Kemijski Sastav Plodova Europskog Pitomog Kestena (Castanea Sativa Mill.) u Hrvatskoj. Ph.D. Thesis, Faculty of Forestry and Wood Technology, University of Zagreb, Zagreb, Croatia, 5 December 2014. [Google Scholar]
- Stojnić, S.; Orlović, S.; Miljković, D.; Von, W.G. Intra- and interprovenance variations in leaf morphometric traits in European beech (Fagus sylvatica L.). Arch. Biol. Sci. 2016, 68, 781–788. [Google Scholar] [CrossRef]
- Stojnić, S.; Viscosi, V.; Marković, M.; Ivanković, M.; Orlović, S.; Tognetti, R.; Cocozza, C.; Vasić, V.; Loy, A. Spatial patterns of leaf shape variation in European beech (Fagus sylvatica L.) provenances. Trees 2021, 1–15. [Google Scholar] [CrossRef]
- Miljković, D.; Stefanović, M.; Orlović, S.; Neđić, M.S.; Kesić, L.; Stojnić, S. Wild cherry (Prunus avium (L.) L.) leaf shape and size variations in natural populations at different elevations. Alp. Bot. 2019, 129, 163–174. [Google Scholar] [CrossRef]
- Said, A.A.; Oukabli, A.; Gaboun, F.; Simard, M.H.; El Modafar, C. Phenotypic biodiversity of an endemic wild pear, Pyrus mamorensis Trab., in North-Western Morocco using morphological descriptors. Genet. Resour. Crop Evol. 2012, 60, 927–938. [Google Scholar] [CrossRef]
- Vidaković, A.; Liber, Z.; Šatović, Z.; Idžojtić, M.; Volenec, I.; Zegnal, I.; Pintar, V.; Radunić, M.; Poljak, I. Phenotypic diversity of almond-leaved pear (Pyrus spinosa Forssk.) along eastern Adriatic coast. Forests 2021, 12, 1630. [Google Scholar] [CrossRef]
- Bednorz, L. Morphological variability of leaves of Sorbus torminalis (L.) Crantz in Poland. Acta Soc. Bot. Pol. 2011, 75, 233–243. [Google Scholar] [CrossRef] [Green Version]
- Zebec, M. Morfologija i Varijabilnost Nizinskog Brijesta (Ulmus minor Mill. sensu latissimo) u Hrvatskoj. Ph.D. Thesis, Faculty of Forestry and Wood Technology, University of Zagreb, Zagreb, Croatia, 19 April 2010. [Google Scholar]
- Zarei, A.; Erfani-Moghadam, J.; Jalilian, H. Assessment of variability within and among four Pyrus species using multivariate analysis. Flora-Morphol. Distrib. Funct. Ecol. Plants 2018, 250, 27–36. [Google Scholar] [CrossRef]
- Matthies, D.; Bräuer, I.; Maibom, W.; Tscharntke, T. Population size and the risk of local extinction: Empirical evidence from rare plants. Oikos 2004, 105, 481–488. [Google Scholar] [CrossRef]
- Stebbins, G.L. Self fertilization and population variability in the higher plants. Am. Nat. 1957, 91, 337–354. [Google Scholar] [CrossRef]
- Cornille, A.; Giraud, T.; Bellard, C.; Tellier, A.; Le Cam, B.; Smulders, M.J.M.; Kleinschmit, J.; Roldan-Ruiz, I.; Gladieux, P. Postglacial recolonization history of the European crabapple (Malus sylvestris Mill.), a wild contributor to the domesticated apple. Mol. Ecol. 2013, 22, 2249–2263. [Google Scholar] [CrossRef] [PubMed]
- Alcántara-Ayala, O.; Oyama, K.; Ríos-Muñoz, C.A.; Rivas, G.; Ramirez-Barahona, S.; Luna-Vega, I. Morphological variation of leaf traits in the Ternstroemia lineataspecies complex (Ericales: Penthaphylacaceae) in response to geographic and climatic variation. PeerJ 2020, 8, e8307. [Google Scholar] [CrossRef] [Green Version]
- Kramer, K. Phenotypic plasticity of the phenology of seven European tree species in relation to climatic warming. Plant Cell Environ. 1995, 18, 93–104. [Google Scholar] [CrossRef]
- Balaguer, L.; Martinez-Ferri, E.; Valladares, F.; Corona, M.E.P.; Baquedano, F.J.; Castillo, F.J.; Manrique, E. Population divergence in the plasticity of the response of Quercus coccifera to the light environment. Funct. Ecol. 2001, 15, 124–135. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Valiente, J.A.; Sanchez-Gomez, D.; Aranda, I.; Valladares, F. Phenotypic plasticity and local adaptation in leaf ecophysiological traits of 13 contrasting cork oak populations under different water availabilities. Tree Physiol. 2010, 30, 618–627. [Google Scholar] [CrossRef] [Green Version]
- Joshi, J.; Schmid, B.; Caldeira, M.C.; Dimitrakopoulos, P.G.; Good, J.; Harris, R.; Hector, A.; Huss-Danell, K.; Jumpponen, A.; Minns, A.; et al. Local adaptation enhances performance of common plant species. Ecol. Lett. 2001, 4, 536–544. [Google Scholar] [CrossRef] [Green Version]
- Bradburd, G.S.; Ralph, P.L.; Coop, G.M. Disentangling the effects of geographic and ecological isolation on genetic differentiation. Evolution 2013, 67, 3258–3273. [Google Scholar] [CrossRef] [Green Version]
- Morente-López, J.; García, C.; Lara-Romero, C.; García-Fernández, A.; Draper, D.; Iriondo, J.M. Geography and Environment Shape Landscape Genetics of Mediterranean Alpine Species Silene ciliata Poiret. (Caryophyllaceae). Front. Plant Sci. 2018, 9, 1698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, S. Isolation by distance. Genetics 1943, 28, 114–138. [Google Scholar] [CrossRef] [PubMed]
- Wang, I.J.; Bradburd, G.S. Isolation by environment. Mol. Ecol. 2014, 23, 5649–5662. [Google Scholar] [CrossRef]
- Sexton, J.P.; Hangartner, S.B.; Hoffmann, A. Genetic isolation by environment or distance: Which pattern of gene flow is most common? Evolution 2013, 68, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Temunović, M.; Franjić, J.; Šatović, Z.; Grgurev, M.; Frascaria-Lacoste, N.; Fernández-Manjarrés, J.F. Environmental heterogeneity explains the genetic structure of continental and Mediterranean populations of Fraxinus angustifolia Vahl. PLoS ONE 2012, 7, e42764. [Google Scholar] [CrossRef] [Green Version]
- Veljković, B.; Šoštarić, I.; Dajić-Stevanović, Z.; Liber, Z.; Šatović, Z. Genetic structure of wild raspberry populations in the Central Balkans depends on their location and on their relationship to commercial cultivars. Sci. Hortic. 2019, 256. [Google Scholar] [CrossRef]
- Gao, J.; Liu, Z.; Zhao, W.; Tomlinson, K.W.; Xia, S.; Zeng, Q.; Wang, X.; Chen, J. Combined genotype and phenotype analyses reveal patterns of genomic adaptation to local environments in the subtropical oak Quercus acutissima. J. Syst. Evol. 2020, 59, 541–556. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Acronyms | M | SD | Min | Max | CV (%) |
|---|---|---|---|---|---|---|
| Leaf area (cm2) | LA | 9.88 | 3.56 | 1.56 | 27.97 | 36.06 |
| Perimeter (cm) | PE | 11.64 | 2.33 | 5.14 | 21.10 | 20.05 |
| Form coefficient | FC | 0.90 | 0.12 | 0.51 | 1.44 | 13.70 |
| Leaf length (cm) | LL | 3.89 | 0.79 | 1.56 | 6.88 | 20.17 |
| Maximal leaf width (cm) | MLW | 3.27 | 0.63 | 1.10 | 5.68 | 19.18 |
| Position of maximal leaf width (cm) | PMLW | 1.56 | 0.37 | 0.13 | 2.95 | 23.84 |
| Leaf width top (cm) | LWT | 1.40 | 0.47 | 0.22 | 3.08 | 33.92 |
| Leaf angle 10 (°) | LA10 | 69.45 | 5.58 | 34.00 | 79.00 | 8.04 |
| Leaf angle 25 (°) | LA25 | 56.36 | 4.74 | 37.00 | 68.00 | 8.40 |
| Petiole length (cm) | PL | 3.54 | 1.15 | 0.61 | 7.95 | 32.50 |
| Trait | Components of the Variance | df | F | Percent of Variability | p-Value |
| Leaf area (LA) | Among populations | 18 | 5.11 | 14.91 | <0.01 |
| Within populations | 171 | 21.56 | 34.61 | <0.01 | |
| Error | 50.48 | ||||
| Perimeter (PE) | Among populations | 18 | 4.44 | 12.58 | <0.01 |
| Within populations | 171 | 20.88 | 34.84 | <0.01 | |
| Error | 52.58 | ||||
| Form coefficient (FC) | Among populations | 18 | 1.37 | 1.64 | 0.15 |
| Within populations | 171 | 23.82 | 42.49 | <0.01 | |
| Error | 55.87 | ||||
| Leaf length (LL) | Among populations | 18 | 3.94 | 11.23 | <0.01 |
| Within populations | 171 | 21.86 | 36.41 | <0.01 | |
| Error | 52.36 | ||||
| Maximal leaf width (MLW) | Among populations | 18 | 5.83 | 17.88 | <0.01 |
| Within populations | 171 | 23.83 | 35.49 | <0.01 | |
| Error | 46.63 | ||||
| Position of maximal leaf width (PMLW) | Among populations | 18 | 1.52 | 2.16 | 0.09 |
| Within populations | 171 | 21.59 | 39.82 | <0.01 | |
| Error | 58.02 | ||||
| Leaf width top (LWT) | Among populations | 18 | 3.03 | 9.68 | <0.01 |
| Within populations | 171 | 32.51 | 46.27 | <0.01 | |
| Error | 44.05 | ||||
| Leaf angle 10 (LA10) | Among populations | 18 | 2.70 | 8.92 | <0.01 |
| Within populations | 171 | 39.66 | 51.28 | <0.01 | |
| Error | 39.80 | ||||
| Leaf angle 25 (LA25) | Among populations | 18 | 2.88 | 9.38 | <0.01 |
| Within populations | 171 | 35.35 | 48.37 | <0.01 | |
| Error | 42.25 | ||||
| Petiole length (PL) | Among populations | 18 | 3.62 | 6.86 | <0.01 |
| Within populations | 171 | 11.35 | 23.90 | <0.01 | |
| Error | 69.24 |
| Trait | PC—Principal Component | ||
|---|---|---|---|
| PC1 | PC2 | PC3 | |
| Leaf area (LA) | −0.979 | −0.065 | −0.008 |
| Perimeter (PE) | −0.955 | 0.203 | −0.088 |
| Form coefficient (FC) | 0.101 | −0.841 | 0.270 |
| Leaf length (LL) | −0.855 | 0.427 | 0.134 |
| Maximal leaf width (MLW) | −0.923 | −0.319 | −0.119 |
| Position of maximal leaf width (PMLW) | −0.758 | 0.534 | −0.112 |
| Leaf width top (LWT) | −0.581 | −0.517 | −0.360 |
| Leaf angle 10 (LA10) | −0.166 | −0.917 | −0.002 |
| Leaf angle 25 (LA25) | −0.120 | −0.955 | −0.161 |
| Petiole length (PL) | −0.629 | −0.195 | 0.682 |
| Eigenvalue | 4.81 | 3.38 | 0.75 |
| % Total variance | 48.14 | 33.79 | 7.45 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vidaković, A.; Šatović, Z.; Tumpa, K.; Idžojtić, M.; Liber, Z.; Pintar, V.; Radunić, M.; Runjić, T.N.; Runjić, M.; Rošin, J.; et al. Phenotypic Variation in European Wild Pear (Pyrus pyraster (L.) Burgsd.) Populations in the North-Western Part of the Balkan Peninsula. Plants 2022, 11, 335. https://doi.org/10.3390/plants11030335
Vidaković A, Šatović Z, Tumpa K, Idžojtić M, Liber Z, Pintar V, Radunić M, Runjić TN, Runjić M, Rošin J, et al. Phenotypic Variation in European Wild Pear (Pyrus pyraster (L.) Burgsd.) Populations in the North-Western Part of the Balkan Peninsula. Plants. 2022; 11(3):335. https://doi.org/10.3390/plants11030335
Chicago/Turabian StyleVidaković, Antonio, Zlatko Šatović, Katarina Tumpa, Marilena Idžojtić, Zlatko Liber, Valentino Pintar, Mira Radunić, Tonka Ninčević Runjić, Marko Runjić, Jakša Rošin, and et al. 2022. "Phenotypic Variation in European Wild Pear (Pyrus pyraster (L.) Burgsd.) Populations in the North-Western Part of the Balkan Peninsula" Plants 11, no. 3: 335. https://doi.org/10.3390/plants11030335
APA StyleVidaković, A., Šatović, Z., Tumpa, K., Idžojtić, M., Liber, Z., Pintar, V., Radunić, M., Runjić, T. N., Runjić, M., Rošin, J., Gaunt, D., & Poljak, I. (2022). Phenotypic Variation in European Wild Pear (Pyrus pyraster (L.) Burgsd.) Populations in the North-Western Part of the Balkan Peninsula. Plants, 11(3), 335. https://doi.org/10.3390/plants11030335