Uncovering Pathways Highly Correlated to NUE through a Combined Metabolomics and Transcriptomics Approach in Eggplant

 , , ,
, , ,  ,
,  , ,
, ,  ,
, 

Abstract
:1. Introduction
2. Results
2.1. Metabolite Detection in Contrasting NUE Genotypes at Different Resupply Time Intervals
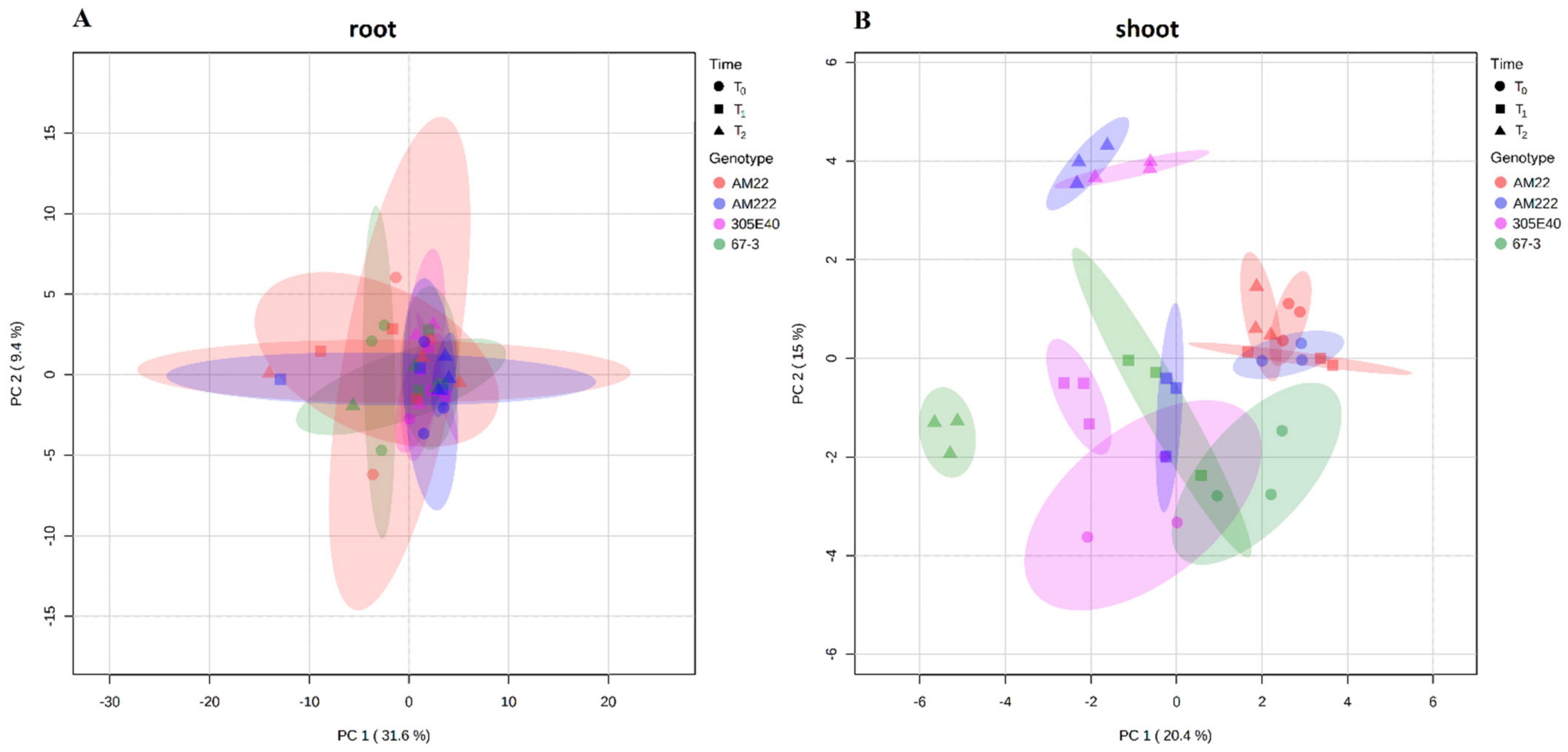
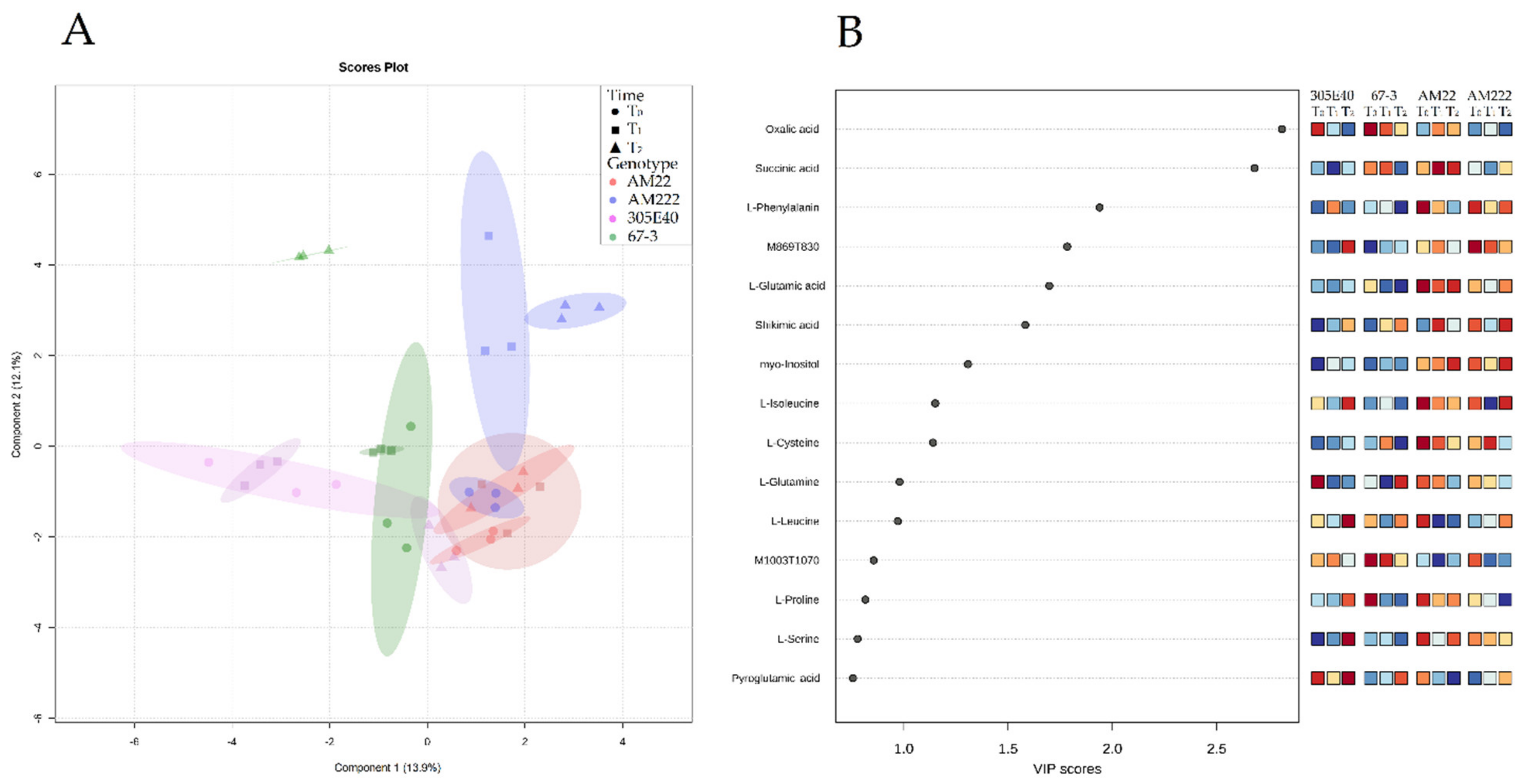
2.2. Multivariate Statistical Analysis of Eggplant Metabolite
2.3. Root and Shoot Metabolites in Eggplant Genotypes under Low N Supply
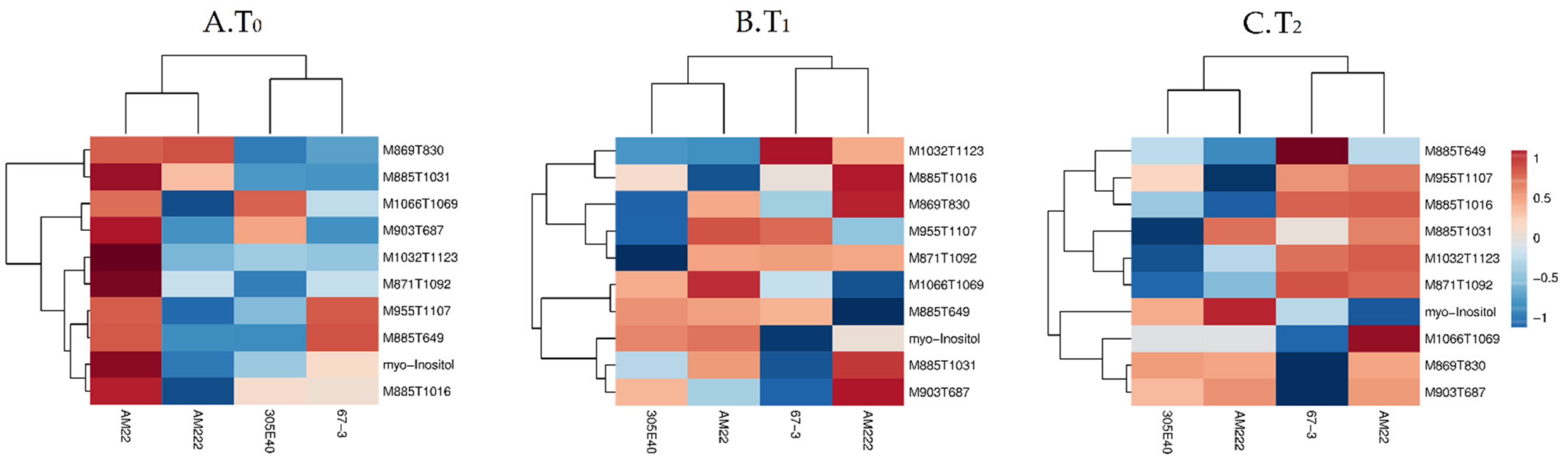
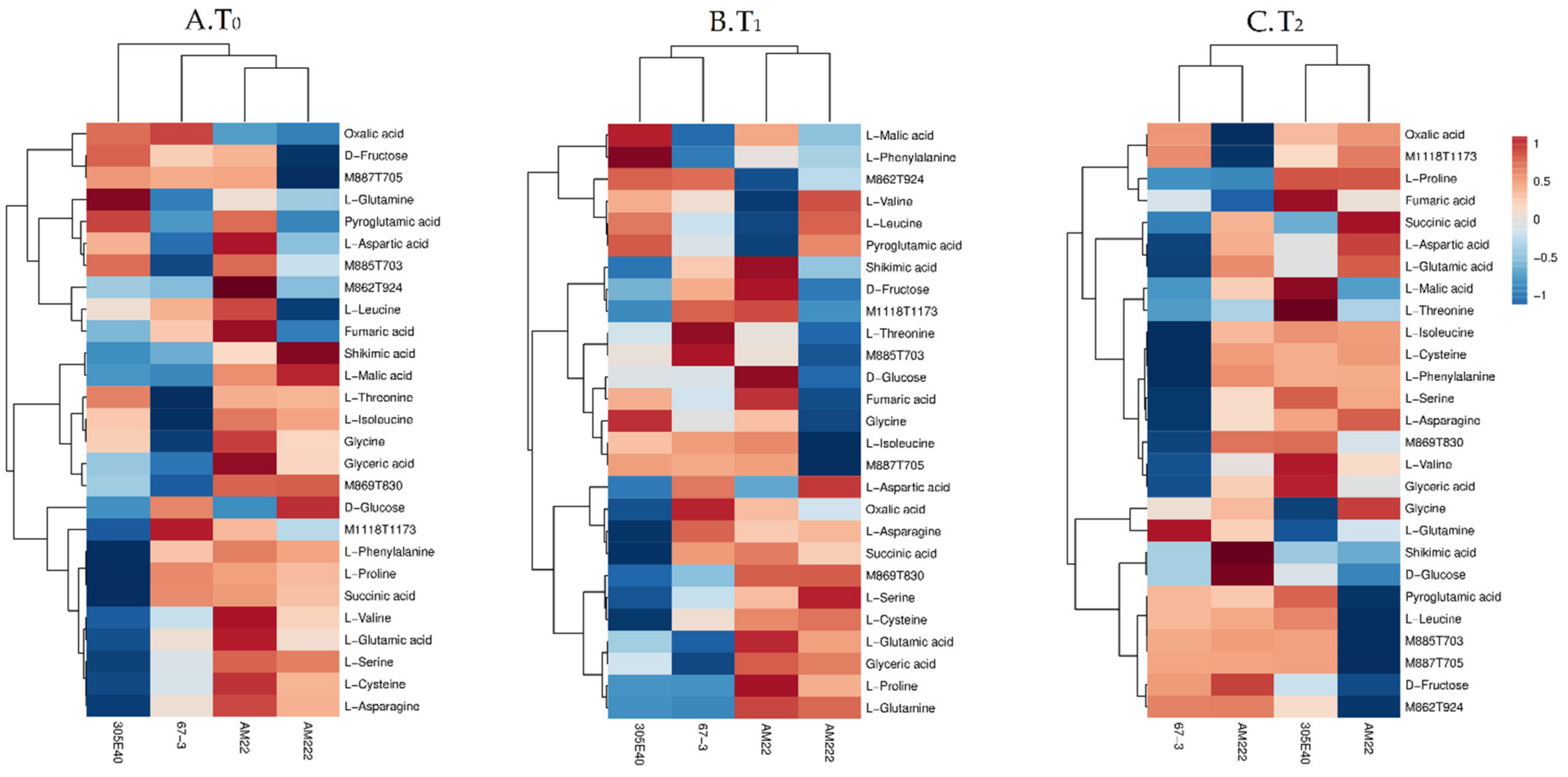
2.4. Comparative Changes in the Primary Metabolite Pathways in Shoot
2.5. Metabolite- and Transcript-Correlation Analysis
2.6. Genotype Clustering and Responses to Nitrogen Starvation
2.7. Genotype Clustering and Short-Term Responses to Low Nitrogen Supply
2.8. Genotype Clustering and Long-Term Responses to Low Nitrogen Supply
2.9. Implementing a Simplified Modeling Scheme
3. Discussion
3.1. Variance and Pathway Analysis
3.2. Metabolite and Transcript Correlation Analysis
3.3. Glycine, Serine, and Threonine Metabolism
3.4. Glyoxylate and Dicarboxylate Metabolism
3.5. Starch and Sucrose Metabolism
4. Materials and Methods
4.1. Plant Materials, Experimental Design, Tissue Sampling, and Sample Preparation
4.2. Metabolite Extraction and Annotation
4.3. RNAseq Analysis Data Validation by qRT-PCR
4.4. Statistical Analysis for Metabolite Profiling
4.5. KEGG Orthology (KO) Annotation and Transcriptomics and Metabolomics Integrated Correlation Network Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Hirel, B.; Le Gouis, J.; Ney, B.; Gallais, A. The challenge of improving nitrogen use efficiency in crop plants: Towards a more central role for genetic variability and quantitative genetics within integrated approaches. J. Exp. Bot. 2007, 58, 2369–2387. [Google Scholar] [CrossRef] [PubMed]
- Vitousek, P.M.; Mooney, H.A.; Lubchenco, J.; Melillo, J.M. Human Domination of Earth’s Ecosystems. Science 1997, 277, 494. [Google Scholar] [CrossRef] [Green Version]
- McArthur, J.W.; McCord, G.C. Fertilizing growth: Agricultural inputs and their effects in economic development. J. Dev. Econ. 2017, 127, 133–152. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Galiano, V.F.; Luque-Espinar, J.A.; Chica-Olmo, M.; Mendes, M.P. Feature selection approaches for predictive modelling of groundwater nitrate pollution: An evaluation of filters, embedded and wrapper methods. Sci. Total Environ. 2018, 624, 661–672. [Google Scholar] [CrossRef] [PubMed]
- Correia, C.M.; Brito, C.; Sampaio, A.; Dias, A.A.; Bacelar, E.; Gonçalves, B.; Ferreira, H.; Moutinho-Pereira, J.; Rodrigues, M.A. Leguminous Cover Crops Improve the Profitability and the Sustainability of Rainfed Olive (Olea europaea L.). Orchards: From Soil Biology to Physiology of Yield Determination. Proc. Environ. Sci. 2015, 29, 282–283. [Google Scholar] [CrossRef] [Green Version]
- Moll, R.H.; Kamprath, E.J.; Jackson, W.A. Analysis and Interpretation of Factors Which Contribute to Efficiency of Nitrogen Utilization. Agron. J. 1982, 74, 562–564. [Google Scholar] [CrossRef]
- Good, A.G.; Shrawat, A.K.; Muench, D.G. Can less yield more? Is reducing nutrient input into the environment compatible with maintaining crop production? Trends Plant Sci. 2004, 9, 597–605. [Google Scholar] [CrossRef]
- Hawkesford, M.J.; Griffiths, S. Exploiting genetic variation in nitrogen use efficiency for cereal crop improvement. Curr. Opin. Plant Biol. 2019, 49, 35–42. [Google Scholar] [CrossRef]
- Allwood, J.W.; Xu, Y.; Martinez-Martin, P.; Palau, R.; Cowan, A.; Goodacre, R.; Marshall, A.; Stewart, D.; Howarth, C. Rapid UHPLC-MS metabolite profiling and phenotypic assays reveal genotypic impacts of nitrogen supplementation in oats. Metabolomics 2019, 15, 42. [Google Scholar] [CrossRef] [Green Version]
- Mibei, E.A.-O.; Owino, W.O.; Ambuko, J.; Giovannoni, J.J.; Onyango, A.N. Metabolomic analyses to evaluate the effect of drought stress on selected African Eggplant accessions. J. Sci. Food Agric. 2018, 98, 205–216. [Google Scholar] [CrossRef]
- Yang, R.; Yang, J.; Yu, J.; Wang, S.; Yang, C.; Xu, F. Effects of different nitrogen application rates on the quality and metabolomics of cigar tobacco. Agron. J. 2022, 1–13. [Google Scholar] [CrossRef]
- Li, H.; Hu, B.; Chu, C. Nitrogen use efficiency in crops: Lessons from Arabidopsis and rice. J. Exp. Bot. 2017, 68, 2477–2488. [Google Scholar] [CrossRef] [PubMed]
- Hazra, P.; Rout, A.; Roy, U.; Nath, S.; Roy, T.; Dutta, R.; Acharya, S.; Mondal, A. Characterization of brinjal (Solanum melongena L.) germplasm. Veg. Sci. 2003, 30, 145–149. [Google Scholar]
- Mauceri, A.; Bassolino, L.; Lupini, A.; Badeck, F.; Rizza, F.; Schiavi, M.; Toppino, L.; Abenavoli, M.R.; Rotino, G.L.; Sunseri, F. Genetic variation in eggplant for Nitrogen Use Efficiency under contrasting NO3− supply. J. Int. Plant Biol. 2020, 62, 487–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mauceri, A.; Abenavoli, M.R.; Toppino, L.; Panda, S.; Mercati, F.; Aci, M.M.; Aharoni, A.; Sunseri, F.; Rotino, G.L.; Lupini, A. Transcriptomic insights on molecular regulation of Solanum melongena L. N-Use Efficiency. J. Exp. Bot. 2021, 72, 4237–4253. [Google Scholar] [CrossRef] [PubMed]
- Hameed, A.; Ijaz, S.; Mohammad, I.S.; Muhammad, K.S.; Akhtar, N.; Khan, H.M.S. Aglycone solanidine and solasodine derivatives: A natural approach towards cancer. Biomed. Pharmacother. 2017, 94, 446–457. [Google Scholar] [CrossRef] [PubMed]
- Lelario, F.; De Maria, S.; Rivelli, A.R.; Russo, D.; Milella, L.; Bufo, S.A.; Scrano, L. A Complete Survey of Glycoalkaloids Using LC-FTICR-MS and IRMPD in a Commercial Variety and a Local Landrace of Eggplant (Solanum melongena L.) and their Anticholinesterase and Antioxidant Activities. Toxins 2019, 11, 230. [Google Scholar] [CrossRef] [Green Version]
- Krapp, A.; Berthomé, R.; Orsel, M.; Mercey-Boutet, S.; Yu, A.; Castaings, L.; Elftieh, S.; Major, H.; Renou, J.-P.; Daniel-Vedele, F. Arabidopsis roots and shoots show distinct temporal adaptation patterns toward nitrogen starvation. Plant Physiol. 2011, 157, 1255–1282. [Google Scholar] [CrossRef] [Green Version]
- Ganie, A.H.; Pandey, R.; Kumar, M.N.; Chinnusamy, V.; Iqbal, M.; Ahmad, A. Metabolite Profiling and Network Analysis Reveal Coordinated Changes in Low-N Tolerant and Low-N Sensitive Maize Genotypes under Nitrogen Deficiency and Restoration Conditions. Plants 2020, 11, 1459. [Google Scholar] [CrossRef]
- Lei, B.; Chang, W.; Zhao, H.; Zhang, K.; Yu, J.; Yu, S.; Cai, K.; Zhang, J.; Lu, K. Nitrogen application and differences in leaf number retained after topping affect the tobacco (Nicotiana tabacum) transcriptome and metabolome. BMC Plant Biol. 2022, 22, 38. [Google Scholar] [CrossRef]
- Amiour, N.; Imbaud, S.; Clément, G.; Agier, N.; Zivy, M.; Valot, B.; Balliau, T.; Armengaud, P.; Quilleré, I.; Cañas, R.; et al. The use of metabolomics integrated with transcriptomic and proteomic studies for identifying key steps involved in the control of nitrogen metabolism in crops such as maize. J. Exp. Bot. 2012, 63, 5017–5033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Hu, J.; Wu, Y.; Wang, J.; Song, H.; Chai, M.; Cong, L.; Miao, F.; Ma, L.; Tang, W.; et al. Integrative analysis of the metabolome and transcriptome reveal the phosphate deficiency response pathways of alfalfa. Plant Physiol. Biochem. 2022, 170, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Lea, P.J.; Sodek, L.; Parry, M.A.J.; Shewry, P.R.; Halford, N.G. Asparagine in plants. Ann. Appl. Biol. 2007, 150, 1–26. [Google Scholar] [CrossRef]
- Braun, H.; Fontes, P.C.R.; Silva, T.P.D.; Finger, F.L.; Cecon, P.R.; Ferreira, A.P.S. Carbohydrates Concentration in leaves of potato plants affected by nitrogen fertilization rates. Rev. Ceres 2016, 63, 241–248. [Google Scholar] [CrossRef]
- Renau-Morata, B.; Molina, R.-V.; Minguet, E.G.; Cebolla-Cornejo, J.; Carrillo, L.; Martí, R.; García-Carpintero, V.; Jiménez-Benavente, E.; Yang, L.; Cañizares, J.; et al. Integrative Transcriptomic and Metabolomic Analysis at Organ Scale Reveals Gene Modules Involved in the Responses to Suboptimal Nitrogen Supply in Tomato. Agronomy 2021, 11, 1320. [Google Scholar] [CrossRef]
- Tahjib-Ul-Arif, M.; Zahan, M.I.; Karim, M.M.; Imran, S.; Hunter, C.T.; Islam, M.S.; Mia, M.A.; Hannan, M.A.; Rhaman, M.S.; Hossain, M.A.; et al. Citric Acid-Mediated Abiotic Stress Tolerance in Plants. Int. J. Mol. Sci. 2021, 22, 7235. [Google Scholar] [CrossRef]
- Atanasova, E. Effect of nitrogen sources on the nitrogenous forms and accumulation of amino acid in head cabbage. Plant Soil Environ. 2008, 54, 66–71. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Yan, L.; Riaz, M.; White, P.J.; Yi, C.; Wang, S.L.; Shi, L.; Xu, F.S.; Wang, C.; Cai, H.M.; et al. Integrated transcriptome and metabolome analysis reveals the physiological and molecular responses of allotetraploid rapeseed to ammonium toxicity. Environ. Exp. Bot. 2021, 189, 104550. [Google Scholar] [CrossRef]
- Chourasia, K.N.; More, S.J.; Kumar, A.; Kumar, D.; Singh, B.; Bhardwaj, V.; Kumar, A.; Das, S.K.; Singh, R.K.; Zinta, G.; et al. Salinity responses and tolerance mechanisms in underground vegetable crops: An integrative review. Planta 2022, 255, 68. [Google Scholar] [CrossRef]
- Khan, S.; Pinto, V.B.; do Amaral Júnior, A.T.; Gonçalves, G.M.B.; Corrêa, C.C.G.; Ferreira, F.R.A.; de Souza, G.A.R.; Campostrini, E.; Freitas, M.S.M.; Vieira, M.E.; et al. Revealing the differential protein profiles behind the nitrogen use efficiency in popcorn (Zea mays var. everta). Sci. Rep. 2022, 12, 1521. [Google Scholar] [CrossRef]
- McAllister, C.H.; Beatty, P.H.; Good, A.G. Engineering nitrogen use efficient crop plants: The current status. Plant Biotechnol. J. 2012, 10, 1011–1025. [Google Scholar] [CrossRef] [PubMed]
- Teng, W.; He, X.; Tong, Y.P. Transgenic approaches for improving use efficiency of nitrogen, phosphorus and potassium in crops. J. Int. Agric. 2017, 16, 2657–2673. [Google Scholar] [CrossRef]
- Thomsen, H.C.; Eriksson, D.; Møller, I.S.; Schjoerring, J.K. Cytosolic glutamine synthetase: A target for improvement of crop nitrogen use efficiency? Trends Plant Sci. 2014, 19, 656–663. [Google Scholar] [CrossRef]
- Cavill, R.; Jennen, D.; Kleinjans, J.; Briedé, J.J. Transcriptomic and metabolomic data integration. Brief Bioinform. 2016, 17, 891–901. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, A.; Dong, Q.; Wang, X.; Gui, H.; Zhang, H.; Zhang, X.; Song, M. Transcriptome Analysis Reveals Differences in Key Genes and Pathways Regulating Carbon and Nitrogen Metabolism in Cotton Genotypes under N Starvation and Resupply. Int. J. Mol. Sci. 2020, 21, 1500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nunes-Nesi, A.; Fernie, A.R.; Stitt, M. Metabolic and signaling aspects underpinning the regulation of plant carbon nitrogen interactions. Mol. Plant 2010, 3, 973–996. [Google Scholar] [CrossRef]
- Fürtauer, L.; Pschenitschnigg, A.; Scharkosi, H.; Weckwerth, W.; Nägele, T. Combined multivariate analysis and machine learning reveals a predictive module of metabolic stress response in Arabidopsis thaliana. Mol. Omics 2018, 14, 437–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pracharoenwattana, I.; Zhou, W.; Keech, O.; Francisco, P.B.; Udomchalothorn, T.; Tschoep, H.; Stitt, M.; Gibon, Y.; Smith, S.M. Arabidopsis has a cytosolic fumarase required for the massive allocation of photosynthate into fumaric acid and for rapid plant growth on high nitrogen. Plant J. 2010, 62, 785–795. [Google Scholar] [CrossRef] [PubMed]
- Araji, S.; Grammer, T.A.; Gertzen, R.; Anderson, S.D.; Mikulic-Petkovsek, M.; Veberic, R.; Phu, M.L.; Solar, A.; Leslie, C.A.; Dandekar, A.M.; et al. Novel roles for the polyphenol oxidase enzyme in secondary metabolism and the regulation of cell death in walnut. Plant Physiol. 2014, 164, 1191–1203. [Google Scholar] [CrossRef] [Green Version]
- Jukanti, A. Distribution, localization, and structure of plant polyphenol oxidases (PPOs). In Polyphenol Oxidases (PPOs) in Plants; Springer: Singapore, 2017; pp. 11–32. [Google Scholar]
- Annunziata, M.G.; Ciarmiello, L.F.; Woodrow, P.; Dell’Aversana, E.; Carillo, P. Spatial and Temporal Profile of Glycine Betaine Accumulation in Plants Under Abiotic Stresses. Front. Plant Sci. 2019, 10, 230. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Mauve, C.; Lamothe-Sibold, M.; Guérard, F.; Glab, N.; Hodges, M.; Jossier, M. Photorespiratory serine hydroxymethyltransferase 1 activity impacts abiotic stress tolerance and stomatal closure. Plant Cell Environ. 2019, 42, 2567–2583. [Google Scholar] [CrossRef] [PubMed]
- Xiaochuang, C.; Chu, Z.; Lianfeng, Z.; Junhua, Z.; Hussain, S.; Lianghuan, W.; Qianyu, J. Glycine increases cold tolerance in rice via the regulation of N uptake, physiological characteristics, and photosynthesis. Plant Physiol. Biochem. 2017, 112, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Yuenyong, W.; Sirikantaramas, S.; Qu, L.J.; Buaboocha, T. Isocitrate lyase plays important roles in plant salt tolerance. BMC Plant Biol. 2019, 19, 472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlüter, U.; Mascher, M.; Colmsee, C.; Scholz, U.; Bräutigam, A.; Fahnenstich, H.; Sonnewald, U. Maize source leaf adaptation to nitrogen deficiency affects not only nitrogen and carbon metabolism but also control of phosphate homeostasis. Plant Physiol. 2012, 160, 1384–1406. [Google Scholar] [CrossRef] [Green Version]
- Moreno, J.I.; Martín, R.; Castresana, C. Arabidopsis SHMT1, a serine hydroxymethyl-transferase that functions in the photorespiratory pathway influences resistance to biotic and abiotic stress. Plant J. 2005, 41, 451–463. [Google Scholar] [CrossRef]
- Moeder, W.; del Pozo, O.; Navarre, D.A.; Martin, G.B.; Klessig, D.F. Aconitase plays a role in regulating resistance to oxidative stress and cell death in Arabidopsis and Nicotiana benthamiana. Plant Mol. Biol. 2007, 63, 273–287. [Google Scholar] [CrossRef]
- Lindén, P.; Keech, O.; Stenlund, H.; Gardeström, P.; Moritz, T. Reduced mitochondrial malate dehydrogenase activity has a strong effect on photorespiratory metabolism as revealed by 13C labelling. J. Exp. Bot. 2016, 67, 3123–3135. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, K.; Komae, K. A rice family 9 glycoside hydrolase isozyme with broad substrate specificity for hemicelluloses in type II cell walls. Plant Cell Physiol. 2006, 47, 1541–1554. [Google Scholar] [CrossRef] [Green Version]
- Seung, D.; Soyk, S.; Coiro, M.; Maier, B.A.; Eicke, S.; Zeeman, S.C. Protein targeting to starch is required for localising granule-bound starch synthase to starch granules and for normal amylose synthesis in Arabidopsis. PLoS Biol. 2015, 13, e1002080. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.-H.P.; Lynd, L.R. Toward an aggregated understanding of enzymatic hydrolysis of cellulose: Non complexed cellulase system. Biotechnol. Bioeng. 2004, 88, 798–842. [Google Scholar] [CrossRef]
- Sarkar, A.K.; Sadhukhan, S. Imperative role of trehalose metabolism and trehalose-6-phosphate signalling on salt stress responses in plants. Physiol. Plant. 2022, e13647. [Google Scholar] [CrossRef]
- Korenblum, E.; Dong, Y.; Szymanski, J.; Panda, S.; Jozwiak, A.; Massalha, H.; Meir, S.; Rogachev, I.; Aharoni, A. Rhizosphere microbiome mediates systemic root metabolite exudation by root-to-root signaling. Proc. Natl. Acad. Sci. USA 2020, 117, 3874–3883. [Google Scholar] [CrossRef] [PubMed]
- Itkin, M.; Heinig, U.; Tzfadia, O.; Bhide, A.J.; Shinde, B.; Cardenas, P.D.; Bocobza, S.E.; Unger, T.; Malitsky, S.; Finkers, R.; et al. Biosynthesis of antinutritional alkaloids in solanaceous crops is mediated by clustered genes. Science 2013, 341, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Tzfadia, O.; Bocobza, S.; Defoort, J.; Almekias-Siegl, E.; Panda, S.; Levy, M.; Storme, V.; Rombauts, S.; Jaitin, D.A.; Keren-Shaul, H.; et al. The ‘TranSeq’ 3′-end sequencing method for high-throughput transcriptomics and gene space refinement in plant genomes. Plant J. 2018, 96, 223–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barchi, L.; Pietrella, M.; Venturini, L.; Minio, A.; Toppino, L.; Acquadro, A.; Andolfo, G.; Aprea, G.; Avanzato, C.; Bassolino, L.; et al. A chromosome-anchored eggplant genome sequence reveals key events in Solanaceae evolution. Sci. Rep. 2019, 9, 11769. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Xia, J.; Psychogios, N.; Young, N.; Wishart, D.S. MetaboAnalyst: A web server for metabolomic data analysis and interpretation. Nucleic Acids Res. 2009, 37, W652–W660. [Google Scholar] [CrossRef] [Green Version]
- Van den Berg, R.A.; Hoefsloot, H.C.; Westerhuis, J.A.; Smilde, A.K.; van der Werf, M.J. Centering, scaling, and transformations: Improving the biological information content of metabolomics data. BMC Genom. 2006, 7, 142. [Google Scholar] [CrossRef] [Green Version]
- Kolde, R. Pheatmap: Pretty Heatmaps. R Package Version 1.0.10. 2012. Available online: https://CRAN.R-project.org/package=pheatmap (accessed on 30 December 2021).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pairwise Comparison in Shoot | Pathway Analysis | Total Cmpd | Hits | Raw p | −log(p) | Holm Adjust | FDR | Impact |
|---|---|---|---|---|---|---|---|---|
| T0_67-3_vs_AM22 | Alanine, aspartate, and glutamate metabolism | 22 | 7 | 0.020155 | 1.6956 | 0.50388 | 0.047029 | 0.64748 |
| T0_305E40_vs_AM22 | 0.002014 | 2.6959 | 0.068486 | 0.0094 | ||||
| T1_AM222_vs_AM22 | 0.00278 | 2.5559 | 0.088969 | 0.010616 | ||||
| T1_67-3_vs_AM22 | 1.99 × 10−5 | 4.7012 | 0.00077607 | 0.000209 | ||||
| T1_305E40_vs_AM22 | 2.3 × 10−5 | 4.6388 | 0.00094188 | 0.00029 | ||||
| T2_AM222_vs_AM22 | Starch and sucrose metabolism | 22 | 2 | 0.003484 | 2.458 | 0.13935 | 0.038299 | 0.39104 |
| T2_67-3_vs_AM22 | Alanine, aspartate, and glutamate metabolism | 22 | 7 | 0.000618 | 3.2088 | 0.021642 | 0.003246 | 0.64748 |
| T2_305E40_vs_AM22 | Glycine, serine, and threonine metabolism | 33 | 5 | 0.000335 | 3.4751 | 0.013731 | 0.00559 | 0.53598 |
| T0_305E40_vs_AM222 | Aminoacyl-tRNA biosynthesis | 46 | 14 | 0.00032 | 3.4948 | 0.013443 | 0.013443 | 0.11111 |
| T1_305E40_vs_AM222 | Alanine, aspartate, and glutamate metabolism | 22 | 7 | 4.37 × 10−5 | 4.3594 | 0.0016609 | 0.000367 | 0.64748 |
| T2_305E40_vs_AM222 | 0.007058 | 2.1513 | 0.26116 | 0.049408 | ||||
| T1_67-3_vs_AM222 | Alanine, aspartate, and glutamate metabolism | 22 | 7 | 0.001201 | 2.9203 | 0.043252 | 0.007209 | 0.64748 |
| T2_67-3_vs_AM222 | Phenylalanine metabolism | 11 | 1 | 9.78 × 10−5 | 4.0098 | 0.0040085 | 0.000851 | 0.47059 |
| T0_67-3_vs_305E40 | Glyoxylate and dicarboxylate metabolism | 29 | 9 | 0.002673 | 2.5731 | 0.10691 | 0.037417 | 0.28209 |
| T1_67-3_vs_305E40 | Alanine, aspartate, and glutamate metabolism | 22 | 7 | 2.06 × 10−5 | 4.6871 | 0.00078109 | 0.000173 | 0.64748 |
| T2_67-3_vs_305E40 | 0.001274 | 2.8949 | 0.033116 | 0.003147 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mauceri, A.; Aci, M.M.; Toppino, L.; Panda, S.; Meir, S.; Mercati, F.; Araniti, F.; Lupini, A.; Panuccio, M.R.; Rotino, G.L.; et al. Uncovering Pathways Highly Correlated to NUE through a Combined Metabolomics and Transcriptomics Approach in Eggplant. Plants 2022, 11, 700. https://doi.org/10.3390/plants11050700
Mauceri A, Aci MM, Toppino L, Panda S, Meir S, Mercati F, Araniti F, Lupini A, Panuccio MR, Rotino GL, et al. Uncovering Pathways Highly Correlated to NUE through a Combined Metabolomics and Transcriptomics Approach in Eggplant. Plants. 2022; 11(5):700. https://doi.org/10.3390/plants11050700
Chicago/Turabian StyleMauceri, Antonio, Meriem Miyassa Aci, Laura Toppino, Sayantan Panda, Sagit Meir, Francesco Mercati, Fabrizio Araniti, Antonio Lupini, Maria Rosaria Panuccio, Giuseppe Leonardo Rotino, and et al. 2022. "Uncovering Pathways Highly Correlated to NUE through a Combined Metabolomics and Transcriptomics Approach in Eggplant" Plants 11, no. 5: 700. https://doi.org/10.3390/plants11050700
APA StyleMauceri, A., Aci, M. M., Toppino, L., Panda, S., Meir, S., Mercati, F., Araniti, F., Lupini, A., Panuccio, M. R., Rotino, G. L., Aharoni, A., Abenavoli, M. R., & Sunseri, F. (2022). Uncovering Pathways Highly Correlated to NUE through a Combined Metabolomics and Transcriptomics Approach in Eggplant. Plants, 11(5), 700. https://doi.org/10.3390/plants11050700