
Modeling Bark Thickness and Bark Biomass on Stems of Four Broadleaved Tree Species



Abstract
:1. Introduction
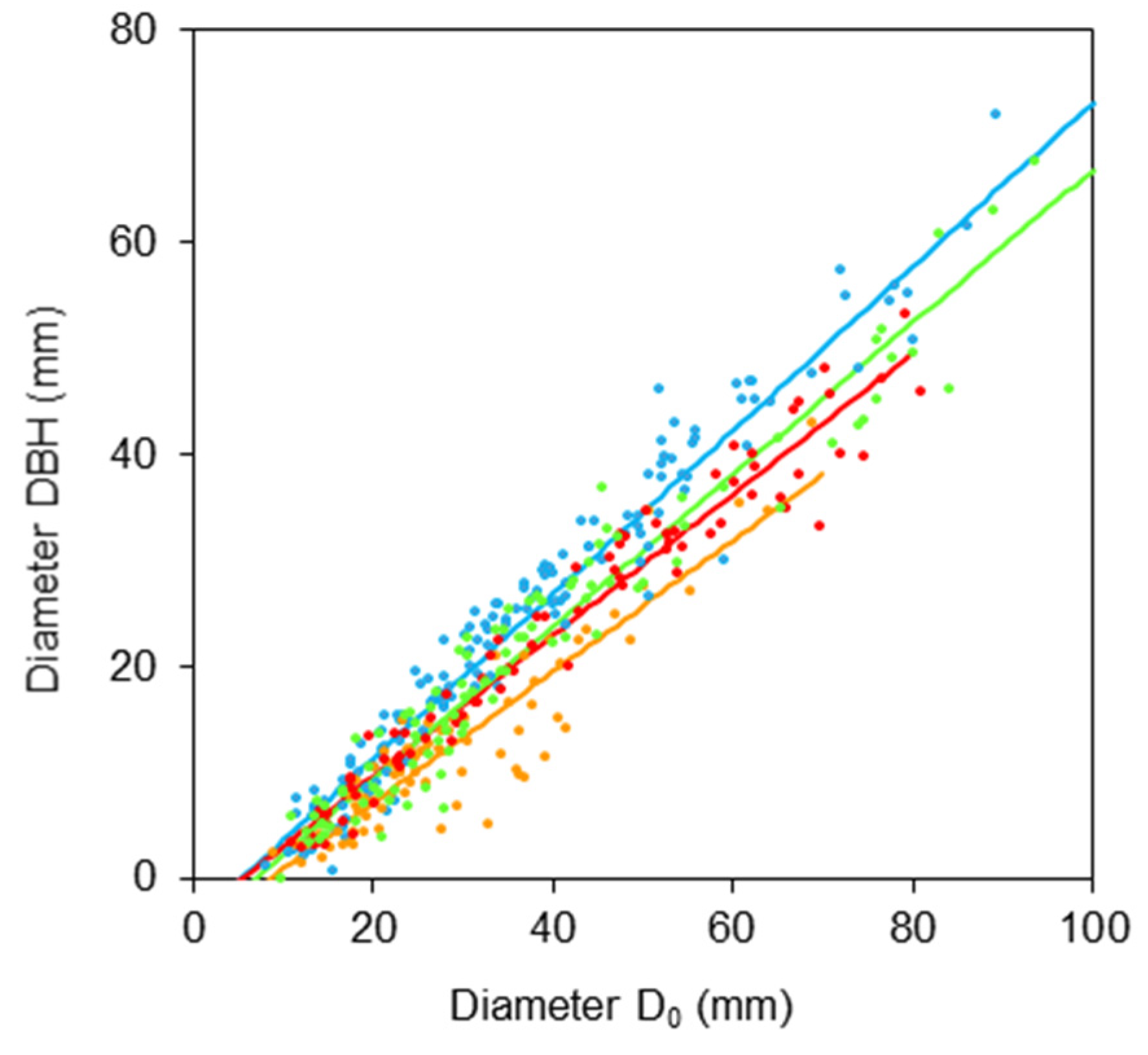
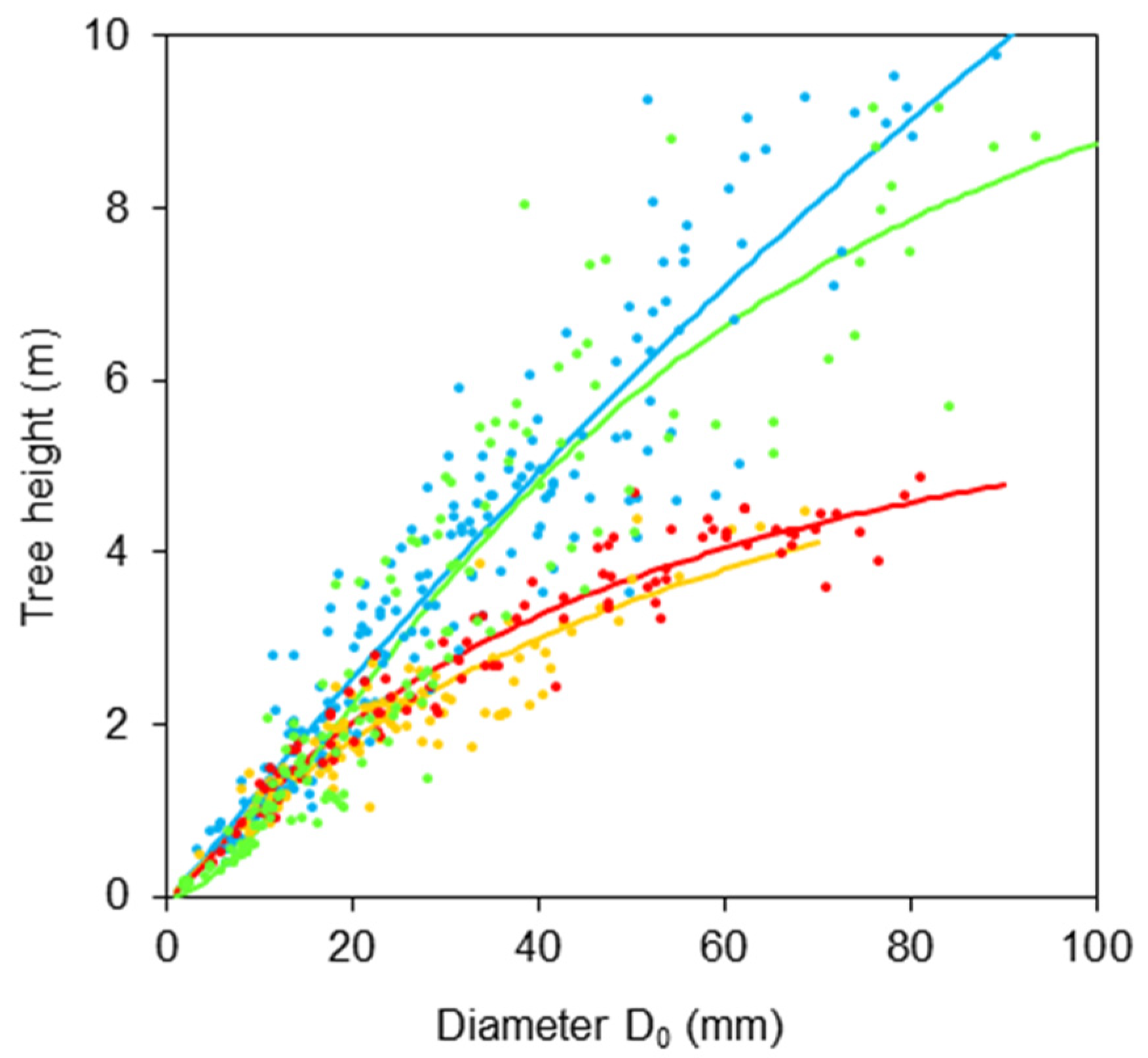
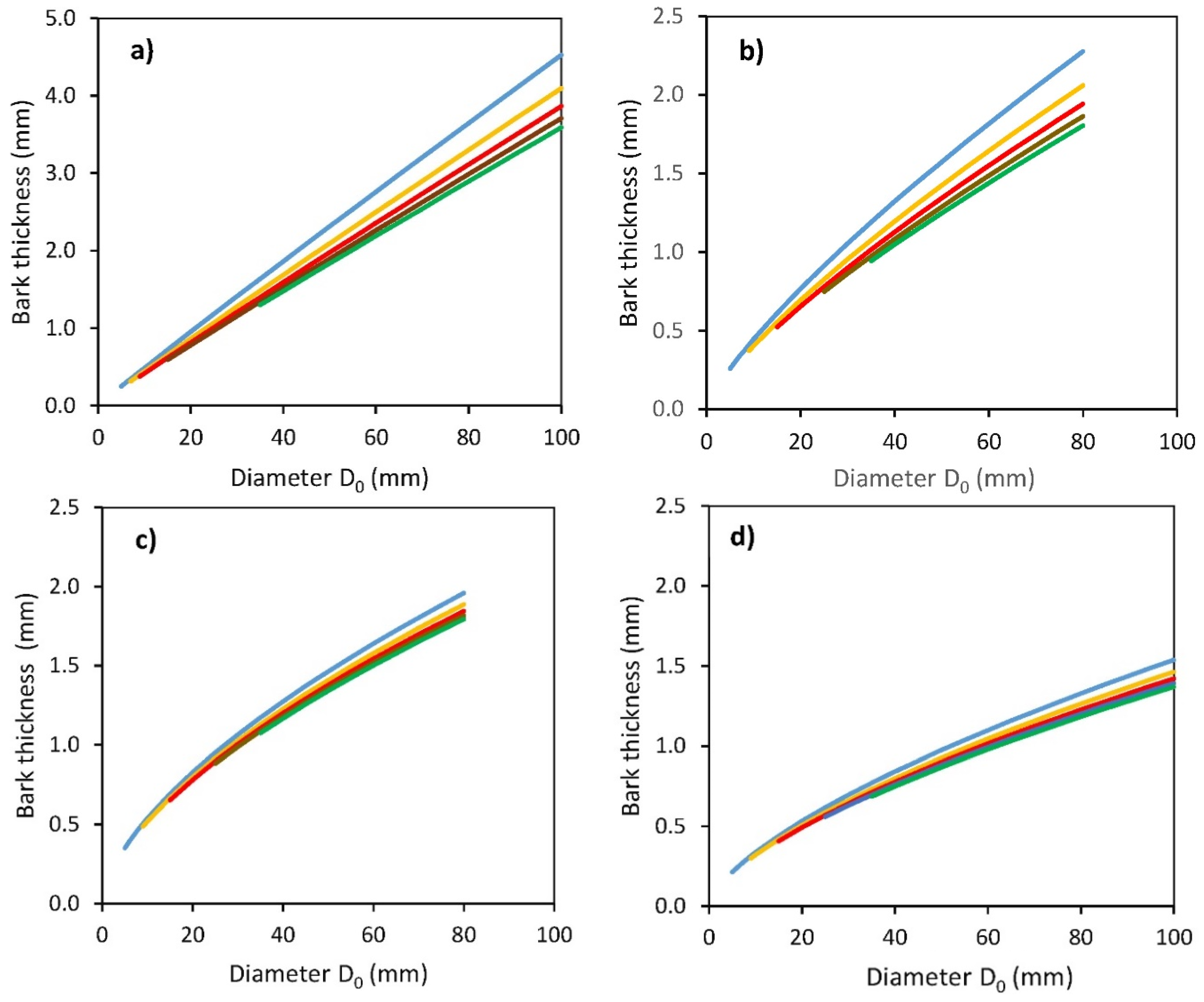
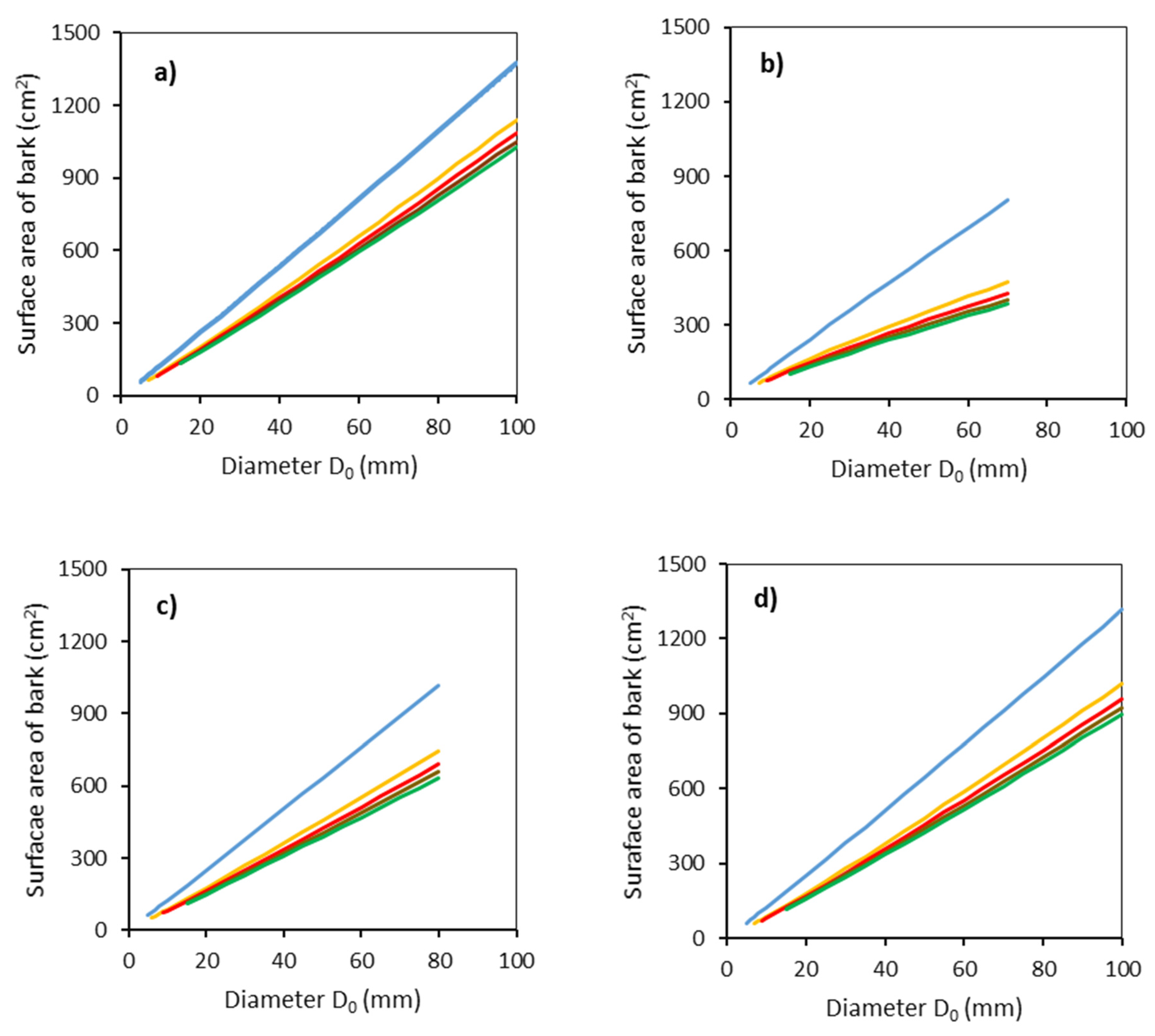
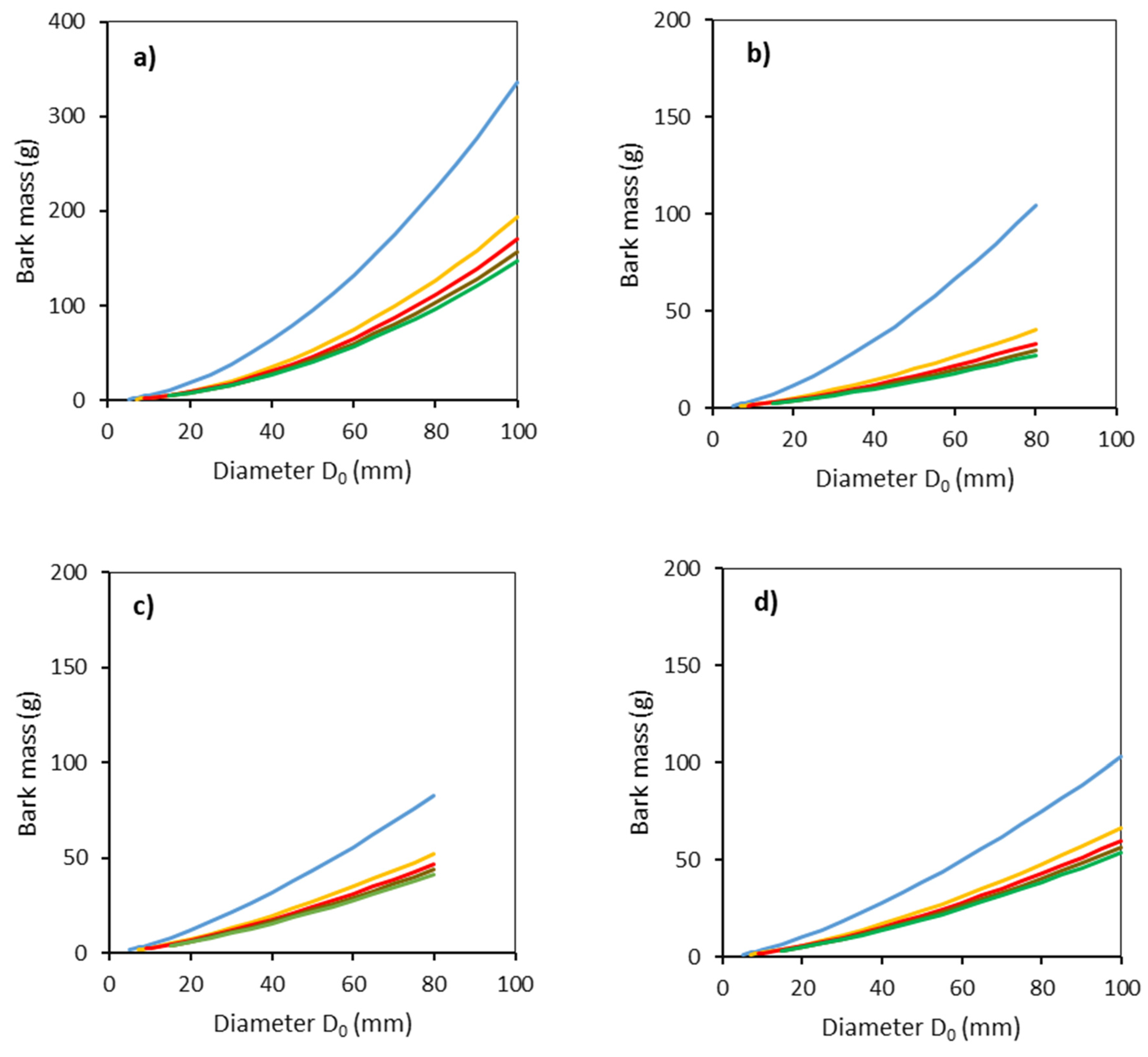
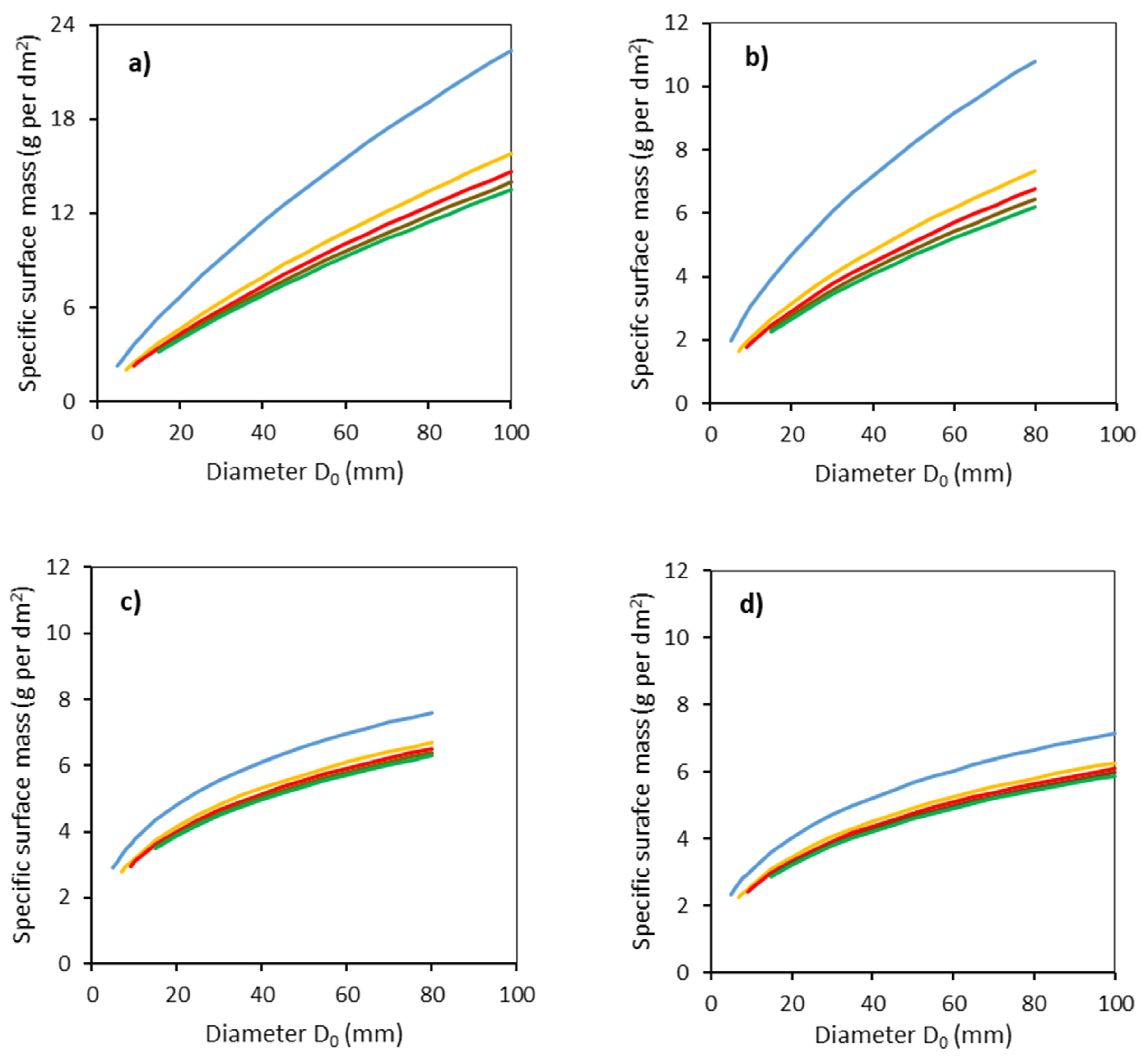
2. Results
3. Discussion
4. Materials and Methods
4.1. Sampling and Data Collection
4.2. Data Processing and Modeling
- H is a tree height (m);
- D0 is a diameter at stem base (mm);
- b0, b1, b2 are regression coefficients.
- DBH is a diameter at breast height (mm);
- D0 is a diameter at stem base (mm);
- b0, b1 are regression coefficients.
- Tb is bark thickness (mm);
- D0 is a diameter at stem base (mm);
- Hg is a distance from the ground level (cm);
- b0, b1, b2 are regression coefficients.
- Sb is bark surface (cm2);
- r1 is a radius of the bottom end (cm);
- r2 is a radius of the top end (cm);
- ls is the length of the section (cm).
- r is a stem radius (mm);
- D0 is a diameter at stem base (mm);
- Hg is a distance from the ground level (cm);
- b0, b1, b2 are regression coefficients.
- Wb is bark mass weight (g);
- Vb is bark volume (cm3);
- ρb is bark density (kg per m3).
- SPH is specific surface mass of bark (g per dm2);
- Wb is bark mass (g);
- Sb is bark surface (dm2).
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Crang, R.; Lyons-Sobaski, S.; Wise, R. Plant Anatomy; Springer International Publishing: Berlin/Heidelberg, Germany, 2018; p. 725. [Google Scholar]
- Kozlowski, T.T.; Pallardy, S.G. Physiology of Woody Plants, 2nd ed.; Academic Press, Inc.: London, UK, 1997; p. 411. [Google Scholar]
- Harkin, J.M.; Rowe, J.W. Bark and Its Possible Uses; Research Note Forest Products Laboratory 091: Madison, WI, USA, 1971; p. 76. [Google Scholar]
- Ryan, M.G.; Asao, S. Phloem transport in trees. Tree Physiol. 2013, 34, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolfe, B.T. Bark eater vapour conductance is associate with drought performance in tropical trees. Biol. Lett. 2020, 16, 20200263. [Google Scholar] [CrossRef] [PubMed]
- Biggs, A.R.; Merrill, W.; Davis, D.D. Discussion: Response of bark tissues to injury and infection. Can. J. For. Res. 1984, 14, 351–356. [Google Scholar] [CrossRef]
- Teskey, T.; Wertin, T.; Bauweraerts, I.; Ameye, M.; McGuire, M.A.; Steppe, K. Responses of tree species to heat waves and extreme heat events. Plant Cell Environ. 2015, 38, 1699–1712. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, A.F.A.; Anderegg, W.R.; Paine, C.E.; Hoffmann, W.A.; Kartzinel, T.; Rabin, S.S.; Sheil, D.; Franco, A.C.; Pacala, S.W. Convergence of bark investment according to fire and climate structures ecosystem vulnerability to future change. Ecol. Lett. 2017, 20, 307–316. [Google Scholar] [CrossRef] [Green Version]
- Christiainsen, E.; Krokene, P.; Berryman, A.A.; Franceschi, V.R.; Krekling, T.; Lieutier, F.; Lönneborg, A.; Solheim, H. Mechanical injury and fungal infection induce acquired resistance in Norway spruce. Tree Physiol. 1999, 19, 399–409. [Google Scholar] [CrossRef] [Green Version]
- Boland, J.M.; Woodward, D.L. Thick bark can protect trees from a severe ambrosia beetle attack. PeerJ 2021, 9, e10755. [Google Scholar] [CrossRef]
- Romero, C. Bark: Structure and functional ecology. In Bark: Use, Management, and Commerce in Africa; Cunningham, A.B., Campbell, B.M., Luckert, M.K., Eds.; New York Botanical Garden Press: New York, NY, USA, 2013; Volume 17, pp. 5–25. [Google Scholar]
- Ilek, A.; Van Stan, J.T.; Morkisz, K.; Kucza, J. Vertical Variability in Bark Hydrology for Two Coniferous Tree Species. Front. For. Glob. Change 2021, 4, 687907. [Google Scholar] [CrossRef]
- Williams, V.L.; Witkowski, E.T.F.; Balkwill, K. Relationship between bark thickness and diameter at breast height for six tree species used medicinally in South Africa. S. Afr. J. Bot. 2007, 73, 449–465. [Google Scholar] [CrossRef] [Green Version]
- Paine, C.E.T.; Stahl, C.; Courtois, E.A.; Patino, S.; Sarmiento, C.; Baraloto, C. Functional explanations for variation in bark thickness in tropical rain forest trees. Funct. Ecol. 2010, 24, 1202–1210. [Google Scholar] [CrossRef]
- Schafer, J.J.; Breslow, B.P.; Hohmann, G.; Hoffmann, A. Relative bark thickness is correlated with tree species distributions along a fire frequency gradient. Fire Ecol. 2015, 11, 74–87. [Google Scholar] [CrossRef]
- Zeibig-Kichas, N.E.; Ardis, C.W.; Berrill, J.-P.; King, J.P. Bark Thickness Equations for Mixed-Coniferous Forest Type in Klamath and Sierra Nevada Mountains of California. Int. J. For. Res. 2016, 2016, 1864039. [Google Scholar]
- Stängle, S.M.; Dormann, C.F. Modelling the variation of bark thickness within and between European silver fir (Abies alba Mill.) trees in southwest Germany. Forestry 2018, 91, 283–294. [Google Scholar] [CrossRef]
- Doruska, P.F.; Patterson, D.; Hartley, J.; Hurd, M.; Hart, T. Newer technologies and bioenergy bring focus back to bark factor equations. J. For. 2009, 107, 38–43. [Google Scholar]
- Pizzi, A. Wood products and green chemistry. Ann. For. Sci. 2016, 73, 185–203. [Google Scholar] [CrossRef] [Green Version]
- Poeiras, A.P.; Oliveira, T.; Reis, J.; Surový, P.; Silva, M.E. Influence of water supply on cork increment and quality in Quercus suber L. Cent. Eur. For. J. 2021, 1, 3–14. [Google Scholar] [CrossRef]
- Morhart, C.; Sheppard, J.P.; Schuler, J.K.; Spiecker, H. Above-ground woody biomass allocation and within tree carbon and nutrient distribution of wild cherry (Prunus aviunm L.)—A case study. For. Ecosyst. 2016, 3, 4. [Google Scholar] [CrossRef] [Green Version]
- Temesgen, H.; Affleck, D.; Poudel, K.; Gray, A.; Sessions, J. A review of the challenges and opportunities in estimating ground forest biomass using tree-level models. Scand. J. For. Res. 2015, 30, 326–335. [Google Scholar] [CrossRef]
- Pajtík, J.; Konôpka, B.; Šebeň, B. Mathematical Biomass Models for Young Individuals of Forest Tree Species in the Region of the Western Carpathians; National Forest Centre: Zvolen, Slovakia, 2018; p. 89. [Google Scholar]
- Myking, T.; Bohler, F.; Austrheim, G.; Solberg, E.J. Life history strategies of aspen (Populus tremula L.) and browsing effects: A literature review. Forestry 2011, 84, 61–71. [Google Scholar] [CrossRef] [Green Version]
- Myking, T.; Solberg, E.J.; Austrheim, G.; Speed, J.D.M.; Bohler, F.; Astrup, R.; Eriksen, R. Browsing of sallow (Salix caprea L.) and rowan (Sorbus aucuparia L.) in the context of life history strategy: A literature review. Eur. J. Forest Res. 2013, 132, 399–409. [Google Scholar] [CrossRef]
- Edenius, L.; Ericsson, G. Effects of ungulate browsing on recruitment of aspen and rowan: A demographic approach. Scand. J. For. Res. 2015, 30, 283–288. [Google Scholar] [CrossRef]
- Konôpka, B.; Pajtík, J.; Shipley, L.A. Intensity of red deer browsing on young rowans differs between freshly-felled and standing individuals. For. Ecol. Manag. 2018, 429, 511–519. [Google Scholar] [CrossRef]
- Borowski, Z.; Gill, W.; Barton, K.; Zajaczkowski, G.; Lukaszewicz, J.; Tittenbrun, A.; Radlisnki, B. Density-related effect of red deer browsing on palatable and unpalatable tree species and forest regeneration dynamics. For. Ecol. Manag. 2021, 496, 119442. [Google Scholar] [CrossRef]
- Konôpka, B.; Šebeň, V.; Pajtík, J.; Shipley, L. Excluding large wild herbivores reduced Norway spruce dominance and supported tree species richness in a young, naturally regenerated stand. Forests 2021, 12, 737. [Google Scholar] [CrossRef]
- Ligot, G.; Gheysen, T.; Lehaire, F.; Hébert, J.; Licoppe, A.; Lejeune, P.; Brostaux, Y. Modelling recent bark stripping by red deer (Cervus elaphus) in South Belgium coniferous stands. Ann. For. Sci. 2012, 70, 309–318. [Google Scholar] [CrossRef] [Green Version]
- Valente, A.M. Overabundant wild ungulate populations in Europe: Management with consideration of socio-ecological consequences. Mamm. Rev. 2020, 50, 353–366. [Google Scholar] [CrossRef]
- Vospernik, S. Probability of Bark Stripping Damage by Red Deer (Cervus elaphus) in Austria. Silva Fenn. 2006, 40, 589–601. [Google Scholar] [CrossRef] [Green Version]
- Speer, H.J. The Fundamentals of Tree-Ring Research; University of Arizona: Terre Haute, Indiana, 2012; p. 510. [Google Scholar]
- Ayanz, J.S.; de Rigo, D.; Caudullo, G.; Durrant, T.H.; Mauri, A. European Atlas of Forest Tree Species; European Commission, Official Journal of European Union: Luxembourg, 2006; Volume 54, p. 200. [Google Scholar]
- Zhao, Q.; Mäkinen, M.; Haapala, A.; Jänis, J. Variation of Bark from Short Rotation Trees by Temperature-Programmed Slow Pyrolysis. ACS Omega 2021, 6, 9771–9779. [Google Scholar] [CrossRef]
- Graves, S.J.; Rifai, S.W.; Putz, F.E. Outer bark thickness decreases more with height on stems of fire-resistant than fire-sensitive Floridian oaks (Quercus spp.; Fagaceae). Am. J. Bot. 2014, 101, 2183–2188. [Google Scholar] [CrossRef] [Green Version]
- Johansson, T. Site Index Curves for European aspen (Populus tremula L.) Growing on Forest Land of Different Soils in Sweden. Silva Fenn. 1996, 30, 437–458. [Google Scholar] [CrossRef] [Green Version]
- Smith, H.C. Bark thickness related to tree diameter in sugar maple (Acer saccharum Marsh.). USDA For. Serv. Res. Note 1967, 107, 1–4. [Google Scholar]
- Li, R.; Weiskittel, A.R. Estimating and predicting bark thickness for seven conifer species in the Acadian Region of North America using a mixed effects modelling approach: Comparison of model forms and subsampling strategies. Eur. J. For. Res. 2011, 130, 219–233. [Google Scholar] [CrossRef]
- Kurt, Y.; Calikoglu, M.; Isik, K. Relationship between bark thickness, tree age and tree diameter in Pinus brutia ten. plantations. Fresenius Environ. Bull. 2021, 30, 3122–3129. [Google Scholar]
- Sommez, T.; Keles, S.; Tilki, F. Effect of aspect, tree age and tree diameter on bark thickness of Picea orientalis. Scand. J. For. Res. 2007, 22, 193–197. [Google Scholar] [CrossRef]
- Berrill, J.-P.; O’Hara, K.L.; Kichas, N.E. Bark Thickness in Coat Redwood (Sequoia sempervivens (D.Don) Endl.) Varies According to Tree- and Crown Size, Stand Structure, Latitude and Genotype. Forests 2020, 11, 637. [Google Scholar] [CrossRef]
- Wilson, B.G.; Witkowski, E.T.F. Seed banks, bark thickness and change in age and size structure (1978–1999) of the African savanna tree, Burkea africana. Plant Ecol. 2003, 167, 151–162. [Google Scholar] [CrossRef]
- Konôpka, B.; Pajtík, J.; Kaštier, P.; Šebeň, V. Estimates of young ash trees dendromass eaten by red deer using allometric models. Zprávy Lesníckého Výzkumu 2012, 57, 283–294. [Google Scholar]
- Rautio, P.; Bergvall, U.A.; Tuomi, J.; Kesti, K.; Leimar, O. Food selection by herbivores and neighbourhood effects in the evolution if plant defences. Ann. Zool. Fenn. 2012, 49, 45–57. [Google Scholar] [CrossRef] [Green Version]
- Standgard, M.; Walsh, D. Improving harvester estimates of bark thickness for radiate pine (Pinus radiata D.Don). South. For. 2011, 73, 101–108. [Google Scholar] [CrossRef]
- Vaško, M.; Garčár, M. Lesnícka typológia a jej význam pri plánovaní drevinového zloženia lesov Slovenska. In Výstupy NLC Pre Lesnícku Prax III; National Forest Centre: Zvolen, Slovakia, 2021; pp. 56–70. (In Slovak) [Google Scholar]
- Konôpka, B.; Pajtík, J.; Šebeň, V.; Bošeľa, M.; Máliš, F.; Priwitzer, T.; Pavlenda, P. The Research Site Vrchslatina—An experimental design and the main aims. Lesn. Cas. For. J. 2013, 59, 203–213. [Google Scholar] [CrossRef] [Green Version]
- Marklund, L.G. Biomass Functions for Norway Spruce (Picea abies (L.) Karst.) in Sweden; Dep. Forest Surwey, Rep. 43; Swedish University of Agricultural Sciences: Umea, Sweden, 1987. [Google Scholar]
- Ledermann, T.; Neuman, M. Biomass equations from data of old long-term experimental plots. Austrian J. For. Sci. 2006, 123, 47–64. [Google Scholar]
- Cienciala, E.; Černý, M.; Apltauer, J.; Exnerová, Z. Biomass functions applicable to European beech. J. For. Sci. 2005, 51, 147–154. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tree Species | b0 | S.E. | b1 | S.E. | R2 | MSE |
|---|---|---|---|---|---|---|
| Common aspen | −3.422 | 0.911 | 0.763 | 0.022 | 0.890 | 29.46 |
| Goat willow | −5.285 | 0.971 | 0.620 | 0.031 | 0.838 | 11.60 |
| Rowan | −3.629 | 0.711 | 0.665 | 0.016 | 0.958 | 7.83 |
| Sycamore | −4.948 | 0.732 | 0.718 | 0.017 | 0.950 | 12.38 |
| Tree Species | b0 | S.E. | P | b1 | S.E. | P | b2 | S.E. | P | R2 | MSE |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Common aspen | 8.221 | 11.812 | 0.487 | 7.077 | 0.693 | <0.001 | 0.021 | 0.009 | 0.017 | 0.897 | 1218.609 |
| Goat willow | 6.921 | 4.691 | 0.143 | 8.127 | 0.542 | <0.001 | 0.126 | 0.016 | <0.001 | 0.812 | 237.939 |
| Rowan | 14.640 | 13.260 | 0.273 | 6.486 | 0.969 | <0.001 | 0.135 | 0.014 | <0.001 | 0.933 | 287.626 |
| Sycamore | 84.237 | 25.897 | 0.001 | 3.273 | 1.311 | 0.014 | 0.073 | 0.014 | <0.001 | 0.882 | 723.968 |
| Tree Species | b0 | S.E. | P | b1 | S.E. | P | b2 | S.E. | P | R2 | MSE |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Common aspen | 0.038 | 0.002 | <0.001 | 1.076 | 0.010 | <0.001 | −0.092 | 0.002 | <0.001 | 0.916 | 0.057 |
| Goat willow | 0.088 | 0.010 | <0.001 | 0.847 | 0.034 | <0.001 | −0.179 | 0.007 | <0.001 | 0.594 | 0.107 |
| Rowan | 0.045 | 0.004 | <0.001 | 1.038 | 0.020 | <0.001 | −0.139 | 0.003 | <0.001 | 0.886 | 0.075 |
| Sycamore | 0.036 | 0.002 | <0.001 | 1.079 | 0.014 | <0.001 | −0.110 | 0.004 | <0.003 | 0.851 | 0.089 |
| Tree Species | b0 | S.E. | b1 | S.E. | b2 | S.E. | R2 | MSE |
|---|---|---|---|---|---|---|---|---|
| Common aspen | 0.092 | 0.016 | 0.968 | 0.016 | −0.144 | 0.004 | 0.786 | 0.287 |
| Goat willow | 0.129 | 0.010 | 0.784 | 0.023 | −0.145 | 0.004 | 0.671 | 0.097 |
| Rowan | 0.160 | 0.011 | 0.620 | 0.018 | −0.055 | 0.005 | 0.681 | 0.089 |
| Sycamore | 0.098 | 0.006 | 0.659 | 0.016 | −0.072 | 0.005 | 0.587 | 0.059 |
| Tree Species | Altitude Range (m a.s.l.) | Number of Stands | Mean Stand Ages | Number of Sampled Trees | Mean Tree Height (Standard Deviation) (m) | Mean Diameter D0 (Standard Deviation) (mm) |
|---|---|---|---|---|---|---|
| Common aspen | 335–870 | 7 | 2–11 | 180 | 3.84 (2.45) | 31.9 (21.1) |
| Goat willow | 750–1030 | 5 | 2–12 | 120 | 2.04 (0.85) | 25.0 (13.2) |
| Rowan | 941–1122 | 5 | 2–12 | 100 | 2.82 (1.21) | 36.7 (21.4) |
| Sycamore | 415–970 | 10 | 2–12 | 200 | 2.85 (2.30) | 25.8 (13.2) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Konôpka, B.; Pajtík, J.; Šebeň, V.; Merganičová, K. Modeling Bark Thickness and Bark Biomass on Stems of Four Broadleaved Tree Species. Plants 2022, 11, 1148. https://doi.org/10.3390/plants11091148
Konôpka B, Pajtík J, Šebeň V, Merganičová K. Modeling Bark Thickness and Bark Biomass on Stems of Four Broadleaved Tree Species. Plants. 2022; 11(9):1148. https://doi.org/10.3390/plants11091148
Chicago/Turabian StyleKonôpka, Bohdan, Jozef Pajtík, Vladimír Šebeň, and Katarína Merganičová. 2022. "Modeling Bark Thickness and Bark Biomass on Stems of Four Broadleaved Tree Species" Plants 11, no. 9: 1148. https://doi.org/10.3390/plants11091148
APA StyleKonôpka, B., Pajtík, J., Šebeň, V., & Merganičová, K. (2022). Modeling Bark Thickness and Bark Biomass on Stems of Four Broadleaved Tree Species. Plants, 11(9), 1148. https://doi.org/10.3390/plants11091148