
The Effect of the Height of Coppicing and Harvest Season on the Yield and Quality of the Essential Oil of Kunzea ambigua


Abstract
:1. Introduction
2. Results
2.1. Growth Rates and Above-Ground Biomass following Harvest of Kunzea Plants According to Harvest Intensity and Season
2.2. The Levels of Starch, Soluble Sugars and NSCs in Coppiced Kunzea Plants Influence on Essential Oils Biosynthesis
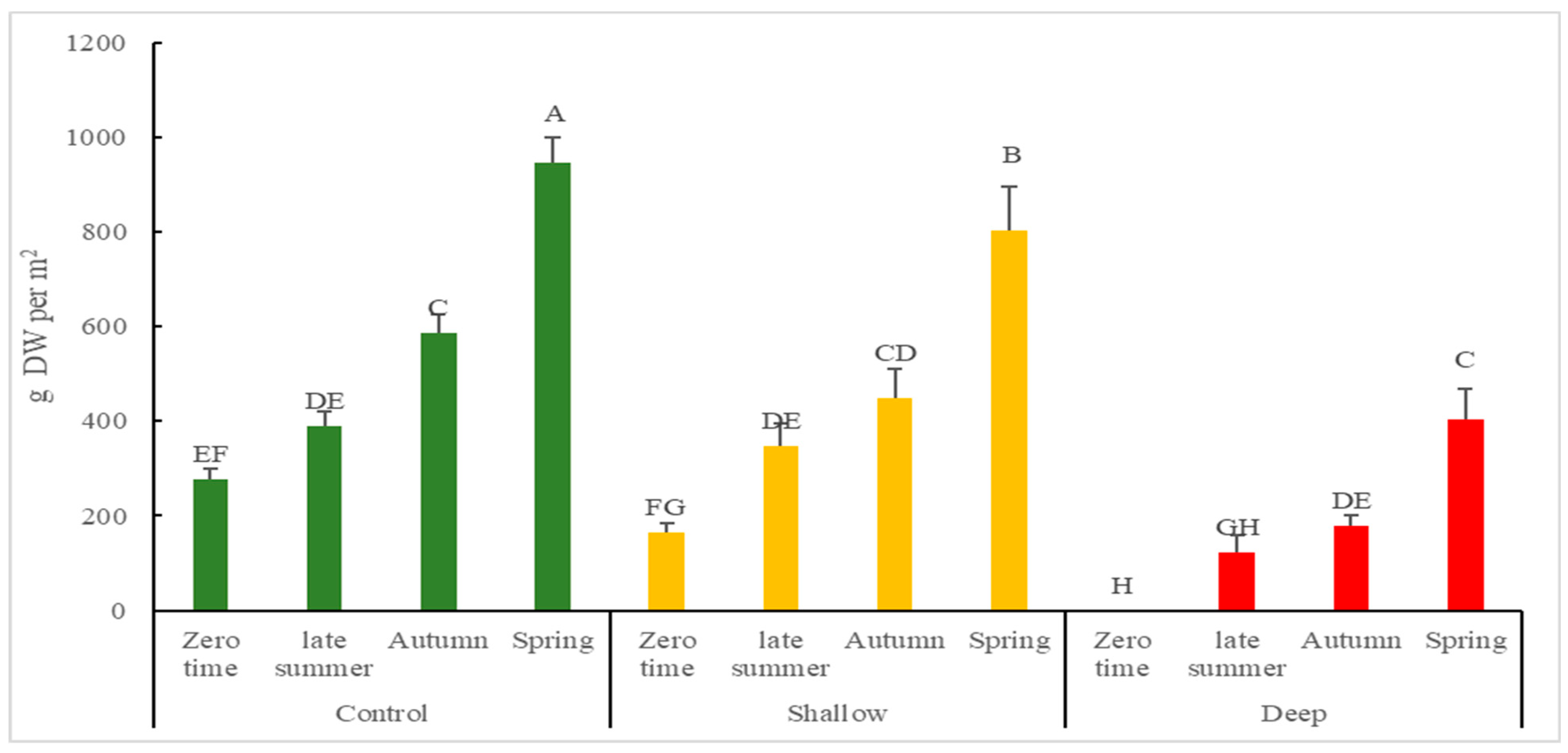
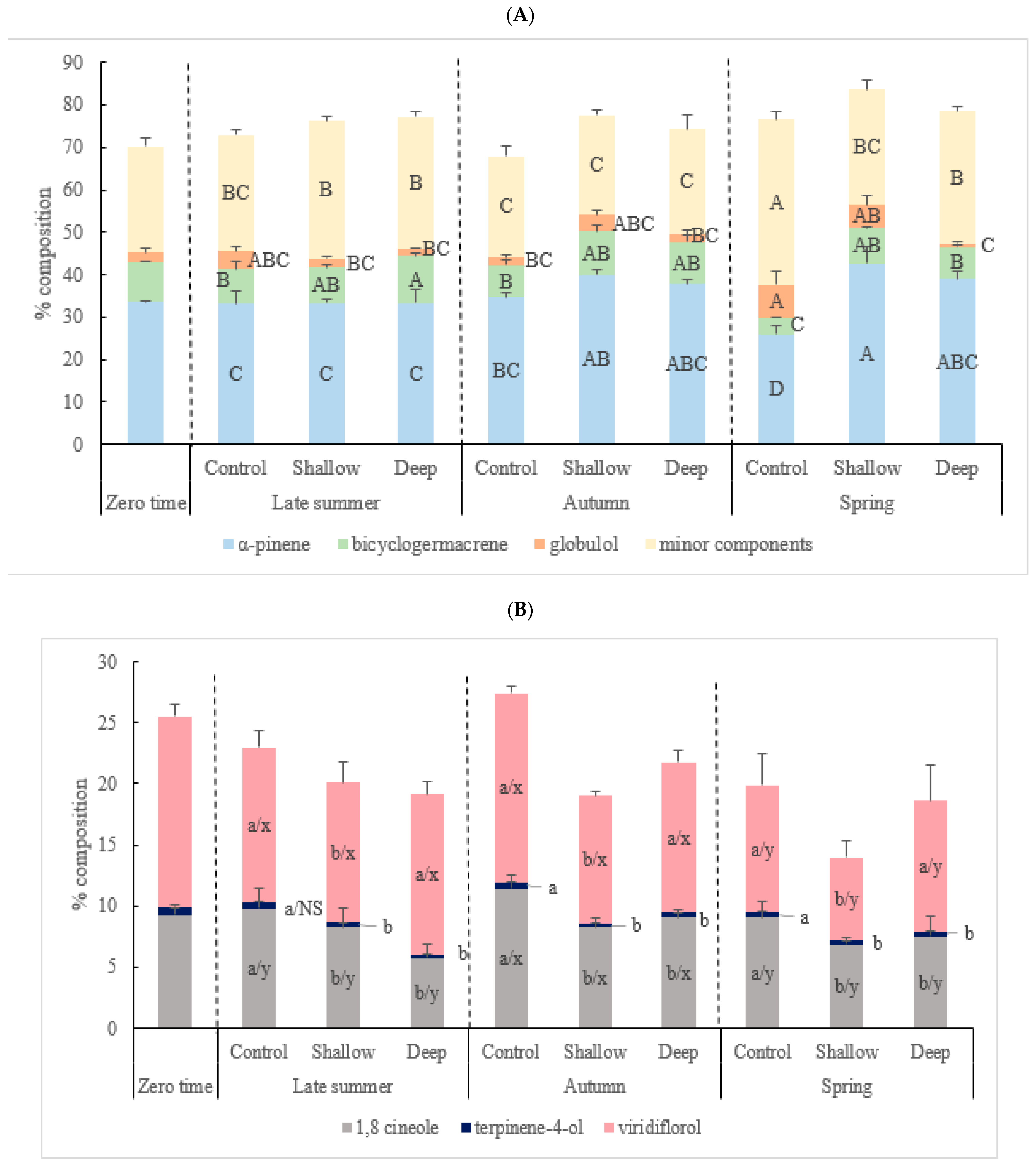
2.3. The Effect of Harvest Intensity and Season on the Chemical Constituents of Kunzea Essential Oil (%)
3. Discussion
3.1. Plant Growth and the Allocation of NSCs of K. ambigua in Response to Harvest Intensity and Season
3.2. The Effect of Harvest Intensity and Season on the Quantity of Kunzea Essential Oil
3.3. The Effect of Harvest Intensity and Season on the Quality of Kunzea Essential Oil
3.4. Optimised Commercial Harvest Production of Kunzea Essential Oil
4. Materials and Methods
4.1. Materials
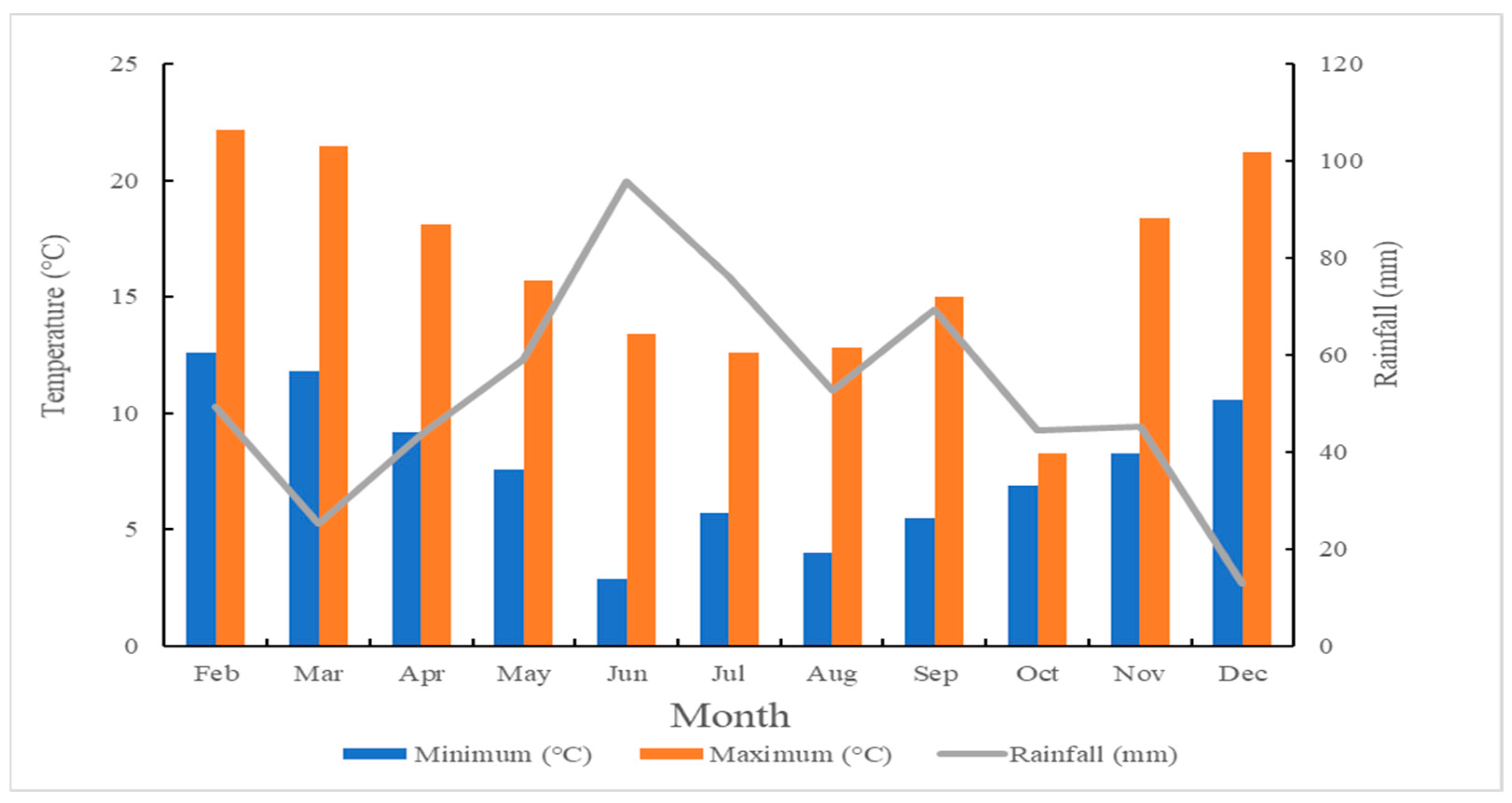
4.2. Experimental Location, Climate and Soil Characteristics
4.3. Plant Materials and Experimental Design
4.4. Isolation of Oils
4.5. Gas Chromatography
4.6. Extraction Methodology for Soluble Sugars and Starch
4.7. Data and Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Elliot, W.R.; Jones, D.L. Encyclopaedia of Australian Plants Suitable for Cultivation; Lothian Publishing Company Pty Co., Ltd.: Melbourne, Australia, 1990; p. 512. [Google Scholar]
- Wrigley, J.W.; Fagg, M. Australian Native Plants: Cultivation, Use in Landscaping and Propagation; Reed New Holland: Sydney, Australia, 2003; p. 720. [Google Scholar]
- Thomas, J. Kunzea Oil: Investigation of Composition, Bioactivity and Therapeitic Potential. Ph.D. Thesis, University of Tasmania, Tasmania, Australia, 2012. [Google Scholar]
- EOT. Essential Oils of Tasmania, Kunzea. Available online: https://www.eotasmania.com.au/kunzea (accessed on 11 May 2019).
- Dragar, V.A. A Preliminary Survey of Selected Species of Endemic Plants to Determine Commercial Cropping Potential for Essential Oils. Ph.D. Thesis, University of Tasmania, Tasmania, Australia, 1986. [Google Scholar]
- Okoh, O.; Sadimenko, A.; Asekun, O.; Afolayan, A. The effects of drying on the chemical components of essential oils of Calendula officinalis L. Afr. J. Biotechnol. 2008, 7, 1500–1502. [Google Scholar]
- Reilly, T. The economics of tea tree. In Proceedings of the Report of the Tea Tree Marketing and Planning Conference, Ballina, Australia, 31 October 1991. [Google Scholar]
- Latt, C.; Nair, P.; Kang, B. Interactions among cutting frequency, reserve carbohydrates, and post-cutting biomass production in Gliricidia sepium and Leucaena leucocephala. Agrofor. Syst. 2000, 50, 27–46. [Google Scholar] [CrossRef]
- Loescher, W.H.; McCamant, T.; Keller, J.D. Carbohydrate reserves, translocation, and storage in woody plant roots. HortScience 1990, 25, 274–281. [Google Scholar] [CrossRef] [Green Version]
- Myers, J.A.; Kitajima, K. Carbohydrate storage enhances seedling shade and stress tolerance in a neotropical forest. J. Ecol. 2007, 95, 383–395. [Google Scholar] [CrossRef]
- Wang, Z.; Bao, W.; Yan, X. Non-structural carbohydrate levels of three co-occurring understory plants and their responses to forest thinning by gap creation in a dense pine plantation. J. For. Res. 2015, 26, 391–396. [Google Scholar] [CrossRef]
- Chiou, T.-J.; Bush, D.R. Sucrose is a signal molecule in assimilate partitioning. Proc. Natl. Acad. Sci. USA 1998, 95, 4784–4788. [Google Scholar] [CrossRef] [Green Version]
- Koch, K. Sucrose metabolism: Regulatory mechanisms and pivotal roles in sugar sensing and plant development. Curr. Opin. Plant Biol. 2004, 7, 235–246. [Google Scholar] [CrossRef]
- Chesney, P.; Vasquez, N. Dynamics of non-structural carbohydrate reserves in pruned Erythrina poeppigiana and Gliricidia sepium trees. Agrofor. Syst. 2007, 69, 89–105. [Google Scholar] [CrossRef]
- Priestly, C. Carbohydrate resources within the perennial plant. Soil Sci. 1962, 94, 198. [Google Scholar] [CrossRef]
- Selvendran, R. Changes in the composition of the xylem exudate of tea plants (Camellia sinensis L.) during recovery from pruning. Ann. Bot. 1970, 34, 825–833. [Google Scholar] [CrossRef]
- Kandiah, S.; Wettasinghe, D.; Wadasinghe, G. Root influence on shoot development in tea (Camellia sinensis (L.) O. Kuntze) following shoot pruning. J. Hortic. Sci. 1984, 59, 581–587. [Google Scholar] [CrossRef]
- Manika, N.; Chanotiya, C.; Negi, M.; Bagchi, G. Copious shoots as a potential source for the production of essential oil in Eucalyptus globulus. Ind. Crops Prod. 2013, 46, 80–84. [Google Scholar] [CrossRef]
- Singh, B.; Sharma, R.A. Plant terpenes: Defense responses, phylogenetic analysis, regulation and clinical applications. 3 Biotech 2015, 5, 129–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goyal, S.; Lambert, C.; Cluzet, S.; Merillon, J.; Ramawat, K.G. Secondary Metabolites and Plant Defence. In Plant Defence: Biological Control; Springer: Berlin, Germany, 2012; pp. 109–138. [Google Scholar]
- Toscano, S.; Trivellini, A.; Cocetta, G.; Bulgari, R.; Francini, A.; Romano, D.; Ferrante, A. Effect of pre-harvest abiotic stresses on the accumulation of bioactive compounds in horticultural produce. Front. Plant Sci. 2019, 10, 1212–1230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, R.; Menary, R. Effects of Photoperiod on the Yield and Composition of Peppermint Oil1. J. Amer. Soc. Hort. Sci. 1979, 104, 699–702. [Google Scholar] [CrossRef]
- Celiktas, O.Y.; Kocabas, E.H.; Bedir, E.; Sukan, F.V.; Ozek, T.; Baser, K. Antimicrobial activities of methanol extracts and essential oils of Rosmarinus officinalis, depending on location and seasonal variations. Food Chem. 2007, 100, 553–559. [Google Scholar] [CrossRef]
- Chauhan, A.; Verma, R.S.; Padalia, R.C. Influence of Biotic and Abiotic Factors on Yield and Quality of Medicinal and Aromatic Plants. Climate Change Effect on Crop Productivity; CRC Press Taylor&Francis Group: Boca Raton, FL, USA, 2014; p. 251. [Google Scholar]
- Hussain, A.I.; Anwar, F.; Sherazi, S.T.H.; Przybylski, R. Chemical composition, antioxidant and antimicrobial activities of basil (Ocimum basilicum) essential oils depends on seasonal variations. Food Chem. 2008, 108, 986–995. [Google Scholar] [CrossRef]
- Hussain, A.I.; Anwar, F.; Nigam, P.S.; Ashraf, M.; Gilani, A.H. Seasonal variation in content, chemical composition and antimicrobial and cytotoxic activities of essential oils from four Mentha species. J. Sci. Food Agric. 2010, 90, 1827–1836. [Google Scholar] [CrossRef]
- Williams, L.; Home, V. Plantation production of oil of melaleuca (tea tree oil)-a revitalized Australian essential oil industry. Search 1988, 19, 294–297. [Google Scholar]
- Oesterheld, M. Effect of defoliation intensity on aboveground and belowground relative growth rates. Oecologia 1992, 92, 313–316. [Google Scholar] [CrossRef]
- Zhao, W.; Chen, S.-P.; Lin, G.-H. Compensatory growth responses to clipping defoliation in Leymus chinensis (Poaceae) under nutrient addition and water deficiency conditions. Plant Ecol. 2008, 196, 85–99. [Google Scholar] [CrossRef]
- Quentin, A.; Beadle, C.; O’grady, A.; Pinkard, E. Effects of partial defoliation on closed canopy Eucalyptus globulus Labilladière: Growth, biomass allocation and carbohydrates. For. Ecol. Manag. 2011, 261, 695–702. [Google Scholar] [CrossRef]
- Honkanen, T.; Haukioja, E. Why does a branch suffer more after branch-wide than after tree-wide defoliation? Oikos 1994, 71, 441–450. [Google Scholar] [CrossRef]
- Trumble, J.; Kolodny-Hirsch, D.; Ting, I. Plant compensation for arthropod herbivory. Ann. Rev. Ent. 1993, 38, 93–119. [Google Scholar] [CrossRef]
- Sterck, F.; Poorter, L.; Schieving, F. Leaf traits determine the growth-survival trade-off across rain forest tree species. Am. Nat. 2006, 167, 758–765. [Google Scholar] [CrossRef]
- Iwasa, Y.; Kubo, T. Optimal size of storage for recovery after unpredictable disturbances. Evol. Ecol. 1997, 11, 41–65. [Google Scholar] [CrossRef]
- Tschaplinski, T.J.; Blake, T.J. Photosynthetic reinvigoration of leaves following shoot decapitation and accelerated growth of coppice shoots. Physiol. Plant. 1989, 75, 157–165. [Google Scholar] [CrossRef]
- Bore, J.; Isutsa, D.; Itulya, F.; Ng’etich, W. Effects of pruning time and resting period on total non-structural carbohydrates, regrowth and yield of tea (Camellia sinensis L.). J. Hortic. Sci. Biotechnol. 2003, 78, 272–277. [Google Scholar] [CrossRef]
- Deslauriers, A.; Caron, L.; Rossi, S. Carbon allocation during defoliation: Testing a defense-growth trade-off in balsam fir. Front. Plant Sci. 2015, 6, 338–352. [Google Scholar] [CrossRef] [Green Version]
- Eyles, A.; Barry, K.M.; Quentin, A.; Pinkard, E.A. Impact of defoliation in temperate eucalypt plantations: Physiological perspectives and management implications. For. Ecol. Manag. 2013, 304, 49–64. [Google Scholar] [CrossRef]
- Rosa, M.; Prado, C.; Podazza, G.; Interdonato, R.; González, J.A.; Hilal, M.; Prado, F.E. Soluble sugars: Metabolism, sensing and abiotic stress: A complex network in the life of plants. Plant. Signal. Behav. 2009, 4, 388–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durand, M.; Mainson, D.; Porcheron, B.; Maurousset, L.; Lemoine, R.; Pourtau, N. Carbon source–sink relationship in Arabidopsis thaliana: The role of sucrose transporters. Planta 2018, 247, 587–611. [Google Scholar] [CrossRef] [PubMed]
- Coley, P.D.; Bryant, J.P.; Chapin, F.S., III. Resource availability and plant antiherbivore defense. Science 1985, 230, 895–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.; Tian, L.; Aziz, N.; Broun, P.; Dai, X.; He, J.; King, A.; Zhao, P.X.; Dixon, R.A. Terpene biosynthesis in glandular trichomes of hop. Plant Physiol. 2008, 148, 1254–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, N.; Luthra, R.; Sangwan, R.S. Effect of leaf position and age on the essential oil quantity and quality in lemongrass (Cymbopogon flexuosus) 1. Planta Med. 1989, 55, 254–256. [Google Scholar] [CrossRef] [PubMed]
- Chimonidou-Pavlidou, D. Correlation between internal and external stages of rose development. Acta Hortic. 1998, 515, 233–244. [Google Scholar] [CrossRef]
- Zekavati, H.R.; Zadeh, A.A. Effect of time and type of pruning on the growth characteristics of rose. Int. J. Agric. Crop Sci. 2013, 6, 698–703. [Google Scholar]
- Thakur, O.; Kumar, V.; Singh, J. A review on advances in pruning to vegetable crops. Int. J. Microbiol. App. Sci. 2018, 7, 3556–3565. [Google Scholar] [CrossRef] [Green Version]
- Eyles, A.; Pinkard, E.A.; Mohammed, C. Shifts in biomass and resource allocation patterns following defoliation in Eucalyptus globulus growing with varying water and nutrient supplies. Tree Physiol. 2009, 29, 753–764. [Google Scholar] [CrossRef] [Green Version]
- Chericoni, S.; Flamini, G.; Campeol, E.; Cioni, P.L.; Morelli, I. GC–MS analyses of the essential oil from the aerial parts of Artemisia verlotiorum: Variability during the year. Biochem. Syst. Ecol. 2004, 32, 423–429. [Google Scholar] [CrossRef]
- Simmons, D.; Parsons, R. Seasonal variation in the volatile leaf oils of twoEucalyptus species. Biochem. Syst. Ecol. 1987, 15, 209–215. [Google Scholar] [CrossRef]
- Boland, D.J.; Brophy, J.J.; Flynn, T.M.; Lassak, E.V. Volatile leaf oils of Eucalyptus delegatensis seedlings. Phytochemistry 1982, 21, 2467–2469. [Google Scholar] [CrossRef]
- Doran, J.; Bell, R. Influence of non-genetic factors on yield of monoterpenes in leaf oils of Eucalyptus camaldulensis. New For. 1994, 8, 363–379. [Google Scholar] [CrossRef]
- Alma, M.H.; Nitz, S.; Kollmannsberger, H.; Digrak, M.; Efe, F.T.; Yilmaz, N. Chemical composition and antimicrobial activity of the essential oils from the gum of Turkish pistachio (Pistacia vera L.). J. Agric. Food Chem. 2004, 52, 3911–3914. [Google Scholar] [CrossRef] [PubMed]
- Lopes, P.M.; Costa, D.C.M.; Alviano, C.S.; Alviano, D.S. Biological Activities of a-Pinene and [beta]-Pinene Enantiomers. Molecules 2012, 17, 6305–6316. [Google Scholar] [CrossRef] [Green Version]
- Durán-Peña, M.J.; Ares, J.M.B.; Hanson, J.R.; Collado, I.G.; Hernández-Galán, R. Biological activity of natural sesquiterpenoids containing a gem-dimethylcyclopropane unit. Nat. Prod. Rep. 2015, 32, 1236–1248. [Google Scholar] [CrossRef]
- Dhami, A.; Singh, A.; Palariya, D.; Kumar, R.; Prakash, O.; Rawat, D.; Pant, A. α-Pinene rich bark essential oils of Zanthoxylum armatum DC. from three different altitudes of Uttarakhand, India and their antioxidant, in vitro anti-inflammatory and antibacterial activity. J. Essent. Oil-Bear. Plants 2019, 22, 660–674. [Google Scholar] [CrossRef]
- Lee, B.-H.; Annis, P.C.; Choi, W.-S. Fumigant toxicity of essential oils from the Myrtaceae family and 1, 8-cineole against 3 major stored-grain insects. J. Stored Prod. Res. 2004, 40, 553–564. [Google Scholar] [CrossRef]
- Rodrigues, F.F.; Costa, J.G.; Coutinho, H.D. Enhancement of the antibiotic activity of gentamicin by volatile compounds of Zanthoxylum articulatum. Indian J. Med. Res. 2010, 131, 833–836. [Google Scholar]
- Williams, L.R.; Lusunzi, I. Essential oil from Melaleuca dissitiflora: A potential source of high quality tea tree oil. Ind. Crops Prod. 1994, 2, 211–217. [Google Scholar] [CrossRef]
- Senatore, F. Influence of harvesting time on yield and composition of the essential oil of a thyme (Thymus pulegioides L.) growing wild in Campania (Southern Italy). J. Agric. Food Chem. 1996, 44, 1327–1332. [Google Scholar] [CrossRef]
- Figueiredo, A.C.; Barroso, J.G.; Pedro, L.G.; Scheffer, J.J. Factors affecting secondary metabolite production in plants: Volatile components and essential oils. Flavour Fragr. J. 2008, 23, 213–226. [Google Scholar] [CrossRef]
- Barra, A. Factors affecting chemical variability of essential oils: A review of recent developments. Nat. Prod. Commun. 2009, 4, 1147–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeffrey, S.J.; Carter, J.O.; Moodie, K.B.; Beswick, A.R. Using spatial interpolation to construct a comprehensive archive of Australian climate data. Environ. Model. Softw. 2001, 16, 309–330. [Google Scholar] [CrossRef]
- Davies, N. Gas chromatographic retention indices of monoterpenes and sesquiterpenes on methyl silicon and Carbowax 20M phases. J. Chromatogr. A 1990, 503, 1–24. [Google Scholar] [CrossRef]
- Thomas, J.; Narkowicz, C.; Jacobson, G.; Davies, N. An examination of the essential oils of Tasmanian Kunzea ambigua, other Kunzea spp. and commercial Kunzea oil. J. Essent. Oil Res. 2010, 22, 381–385. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Season (S) | Initial Harvest Intensity (HI) | GR of Biomass (g DW/m2/Day) |
|---|---|---|
| Zero time | Control | |
| Shallow | ||
| Deep | ||
| Late summer | Control | 6.98 ± 0.54 A |
| Shallow | 3.29 ± 0.18 C | |
| Deep | 2.21 ± 0.32 DE | |
| Autumn | Control | 5.74 ± 0.40 B |
| Shallow | 2.80 ± 0.11 CD | |
| Deep | 1.76 ± 0.22 EF | |
| Spring | Control | 2.76 ± 0.16 CD |
| Shallow | 1.87 ± 0.12 EF | |
| Deep | 1.18 ± 0.18 F | |
| HI | *** | |
| S | *** | |
| HI × S | *** |
| Starch (mg/g DW) | Fructose (mg/g DW) | Glucose (mg/g DW) | Sucrose (mg/g DW) | Total Soluble Sugar (mg/g DW) | NSCs (mg/g DW) | |
|---|---|---|---|---|---|---|
| Leaves | ||||||
| Control | 2.51 ± 0.08 a | 9.48 ± 1.12 b | 7.68 ± 0.99 c | 23.53 ± 0.46 b | 40.69 ± 2.56 c | 43.21 ± 2.55 c |
| Shallow | 1.90 ± 0.14 b | 10.96 ± 0.58 b | 11.53 ± 0.82 b | 41.54 ± 2.22 a | 64.02 ± 1.02 b | 65.92 ± 1.11 b |
| Deep | 1.82 ± 0.04 b | 13.98 ± 1.04 a | 14.31 ± 0.76 a | 44.96 ± 2.17 a | 73.25 ± 3.71 a | 75.07 ± 3.74 a |
| Branches | ||||||
| Control | 4.24 ± 0.34 NS | 6.35 ± 0.50 a | 7.34 ± 0.65 a | 19.38 ± 2.09 a | 33.07 ± 3.23 a | 37.31 ± 3.56 a |
| Shallow | 5.29 ± 0.65 | 3.61 ± 0.60 c | 3.48 ± 0.59 c | 13.35 ± 1.87 b | 20.44 ± 3.05 b | 25.73 ± 3.26 b |
| Deep | 5.08 ± 0.25 | 5.06 ± 0.33 b | 5.76 ± 0.59 b | 19.14 ± 1.60 a | 29.97 ± 1.50 a | 35.05 ± 1.33 a |
| Roots | ||||||
| Control | 9.31 ± 0.89 a | 3.62 ± 0.46 NS | 3.83 ± 0.45 NS | 16.79 ± 2.58 a | 24.23 ± 3.45 a | 33.54 ± 3.48 a |
| Shallow | 8.33 ± 1.21 a | 3.63 ± 0.56 | 3.50 ± 0.55 | 13.43 ± 1.73 b | 20.56 ± 2.83 ab | 28.89 ± 2.05 ab |
| Deep | 7.51 ± 0.59 b | 3.13 ± 0.34 | 3.01 ± 0.30 | 12.27 ± 0.35 b | 18.41 ± 0.77 b | 25.92 ± 0.96 b |
| Season (S) | Initial Harvest Intensity (HI) | Essential Oil Content (% DW) | Essential Oil Yield (g DW Per m2) | Cumulative Essential Oil Yield (g DW Per m2) |
|---|---|---|---|---|
| Zero time | Control | 1.96 ± 0.24 a | 7.01 ± 1.01 C | 0 H |
| Shallow | 2.04 ± 0.12 a | 3.11 ± 0.15 E | 3.11 ± 0.15 G | |
| Deep | 1.96 ± 0.24 b | 0 F | 7.01 ± 1.01 F | |
| Late summer | Control | 1.84 ± 0.06 a | 7.19 ± 0.96 C | 7.19 ± 0.96 EF |
| Shallow | 1.98 ± 0.20 a | 3.65 ± 0.89 DE | 6.76 ± 1.04 F | |
| Deep | 1.66 ± 0.13 b | 2.05 ± 0.53 EF | 9.06 ± 1.54 DEF | |
| Autumn | Control | 1.90 ± 0.15 a | 11.13 ± 1.31 B | 11.13 ± 1.31 CD |
| Shallow | 2.00 ± 0.13 a | 5.72 ± 0.31 CD | 8.83 ± 0.46 DEF | |
| Deep | 1.65 ± 0.13 b | 2.97 ± 0.18 E | 9.98 ± 1.09 DE | |
| Spring | Control | 1.80 ± 0.19 a | 17.05 ± 1.21 A | 17.05 ± 1.21 a/A |
| Shallow | 1.84 ± 0.11 a | 11.79 ± 0.23 B | 14.90 ± 0.48 b/AB | |
| Deep | 1.54 ± 0.32 b | 6.24 ± 0.18 C | 13.25 ± 0.65 c/BC | |
| HI | * | *** | * | |
| S | NS | *** | *** | |
| HI × S | NS | ** | *** |
| The Major Chemical Components in Kunzea Oil | Minor Components | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| α-Pinene | 1,8-Cineole | Terpinen-4-ol | α-Terpineol | Bicyclogermacrene | Globulol | Viridiflorol | Ledol | ||
| HI | *** | ** | ** | ** | *** | ** | * | * | NS |
| S | * | * | NS | * | *** | * | ** | * | *** |
| HI × S | *** | NS | NS | NS | * | * | NS | NS | *** |
| Texture | Nitrate Nitrogen (mg/kg) | Phosphorus (mg/kg) | Potassium (mg/kg) | Sulfur (mg/kg) | pH Level (Cacl2) | DTPA Copper (mg/kg) | DTPA Iron (mg/kg) | DTPA Manganese (mg/kg) |
|---|---|---|---|---|---|---|---|---|
| 1.88 ± 0.25 | <1 | 8.25 ± 1.50 | 67.25 ± 36.39 | 2.37 ± 0.47 | 4.25 ± 0.26 | 0.52 ± 0.15 | 156.78 ± 31.57 | 1.36 ± 0.98 |
| Components | RT 2 | KI 3 Calc. | KI Lit 4 Ref | |
|---|---|---|---|---|
| 1 | α-pinene | 2.906 | 917 | 938 |
| 2 | camphene | 3.401 | 969 | 952 |
| 3 | sabinene | 3.44 | 973 | 977 |
| 4 | β-pinene | 3.618 | 990 | 982 |
| 5 | limonene | 4.018 | 1015 | 1032 |
| 6 | 1,8-cineole | 4.253 | 1026 | 1036 |
| 7 | (Z)-B-ocimene | 4.517 | 1038 | 1036 |
| 8 | (E)-B-ocimene | 4.706 | 1046 | 1046 |
| 9 | isoamyl butyrate | 4.889 | 1054 | 1054 |
| 10 | cis-linalool oxide | 5.428 | 1075 | 1071 |
| 11 | terpinolene | 5.53 | 1078 | 1085 |
| 12 | linalool | 5.938 | 1093 | 1097 |
| 13 | isoamyl isovalerate | 6.173 | 1102 | 1102 |
| 14 | trans-pinocarveol | 6.646 | 1142 | 1143 |
| 15 | pinocarvone | 6.719 | 1148 | 1165 |
| 16 | terpinen-4-ol | 6.925 | 1165 | 1170 |
| 17 | α-terpineol | 7.194 | 1186 | 1197 |
| 18 | citronellol | 7.958 | 1228 | 1227 |
| 19 | geraniol | 8.493 | 1253 | 1252 |
| 20 | ϒ-elemene | 9.695 | 1303 | 1333 |
| 21 | α-cubebene | 10.531 | 1351 | 1352 |
| 22 | α-copaene | 10.972 | 1374 | 1380 |
| 23 | β-elemene | 11.299 | 1391 | 1392 |
| 24 | α-gurjunene | 11.641 | 1409 | 1412 |
| 25 | β-caryophyllene | 11.828 | 1419 | 1425 |
| 26 | aromadendrene | 12.228 | 1441 | 1444 |
| 27 | a-humulene | 12.415 | 1450 | 1461 |
| 28 | allo-aromadendrene | 12.645 | 1462 | 1466 |
| 29 | germacrene D | 13.189 | 1489 | 1468 |
| 30 | bicyclogermacrene | 13.36 | 1497 | 1501 |
| 31 | calamenene | 13.88 | 1525 | 1527 |
| 32 | palustrol | 14.699 | 156 | 1577 |
| 33 | spathulenol | 14.896 | 1578 | 1584 |
| 34 | globulol | 15.013 | 1584 | 1594 |
| 35 | viridiflorol | 15.189 | 1593 | 1603 |
| 36 | ledol | 15.375 | 1601 | 1613 |
| 37 | isospathulenol | 16.07 | 1622 | 1639 |
| 38 | α-muurolol | 16.308 | 1628 | 1654 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, C.; Garland, S.M.; Close, D.C. The Effect of the Height of Coppicing and Harvest Season on the Yield and Quality of the Essential Oil of Kunzea ambigua. Plants 2023, 12, 20. https://doi.org/10.3390/plants12010020
Park C, Garland SM, Close DC. The Effect of the Height of Coppicing and Harvest Season on the Yield and Quality of the Essential Oil of Kunzea ambigua. Plants. 2023; 12(1):20. https://doi.org/10.3390/plants12010020
Chicago/Turabian StylePark, Chanjoo, Sandra M. Garland, and Dugald C. Close. 2023. "The Effect of the Height of Coppicing and Harvest Season on the Yield and Quality of the Essential Oil of Kunzea ambigua" Plants 12, no. 1: 20. https://doi.org/10.3390/plants12010020
APA StylePark, C., Garland, S. M., & Close, D. C. (2023). The Effect of the Height of Coppicing and Harvest Season on the Yield and Quality of the Essential Oil of Kunzea ambigua. Plants, 12(1), 20. https://doi.org/10.3390/plants12010020