
Biocuration of a Transcription Factors Network Involved in Submergence Tolerance during Seed Germination and Coleoptile Elongation in Rice (Oryza sativa)

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Co-Expression Analysis of Transcription Factor Genes during Rice Seed Germination and Coleoptile Elongation under Submergence
2.2. Key Transcription Factors Integrate Multiple Signaling Pathways to Regulate Rice Seed Germination and Coleoptile Elongation under Submergence

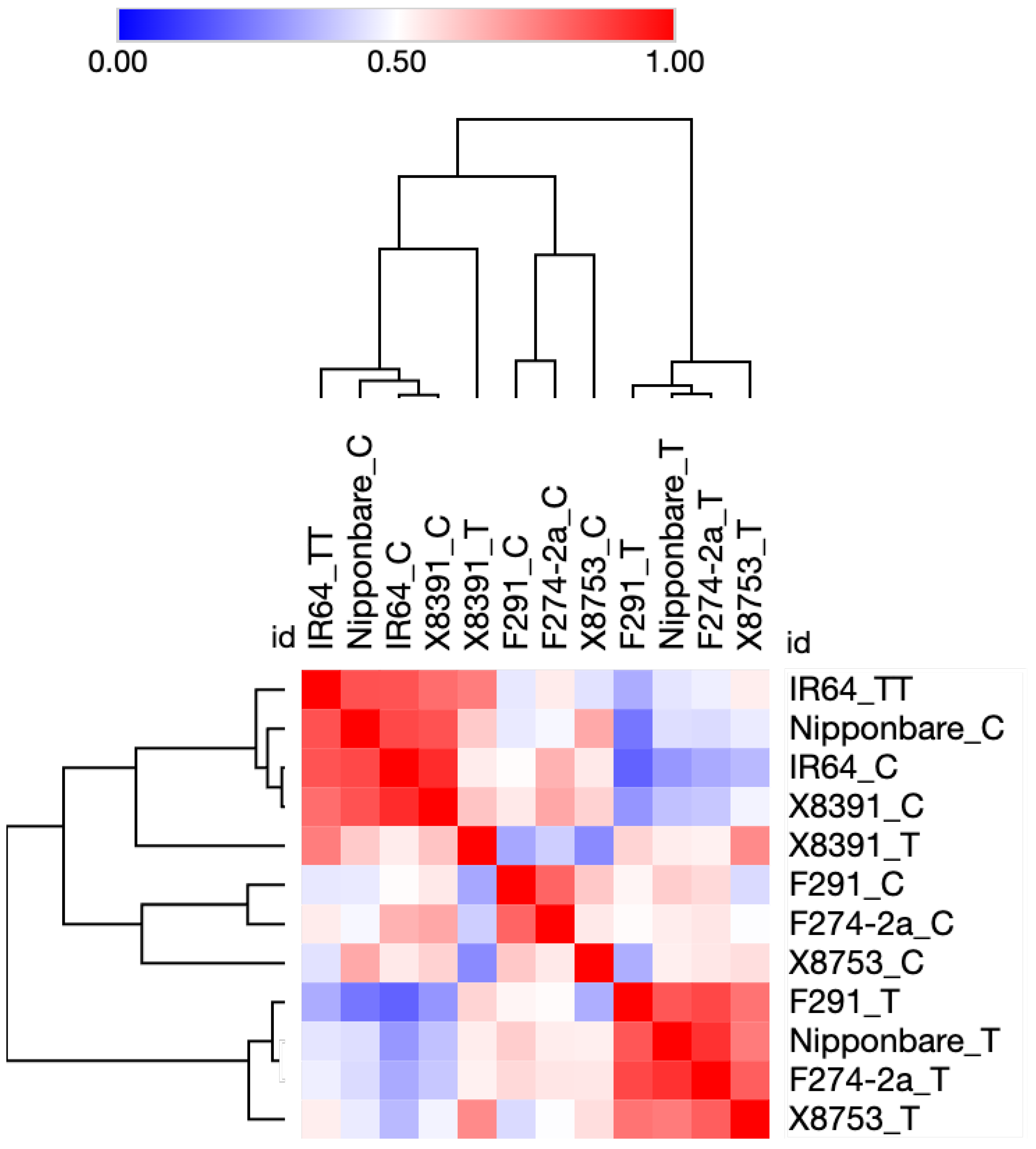{kind=link}
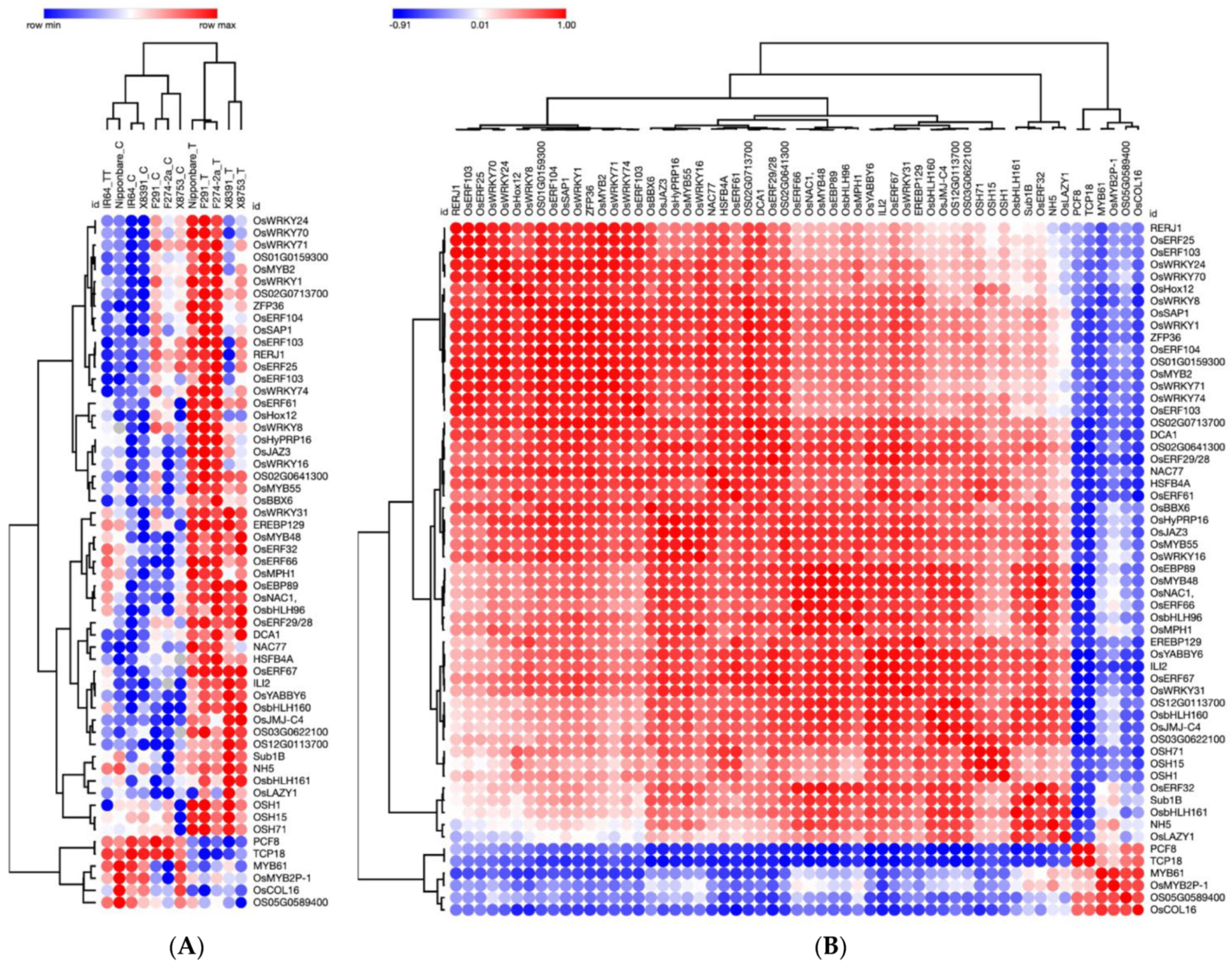{kind=link}
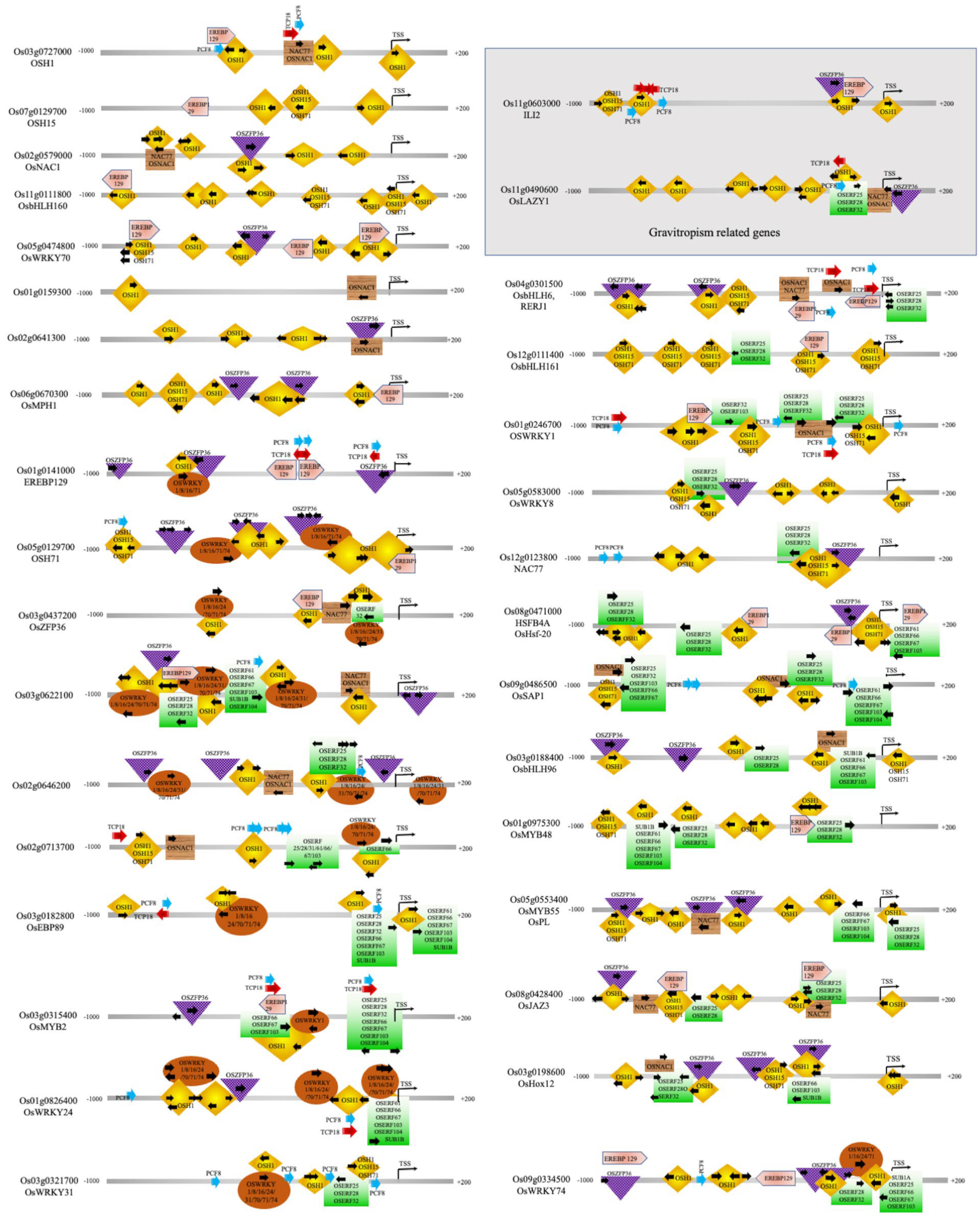{kind=link}
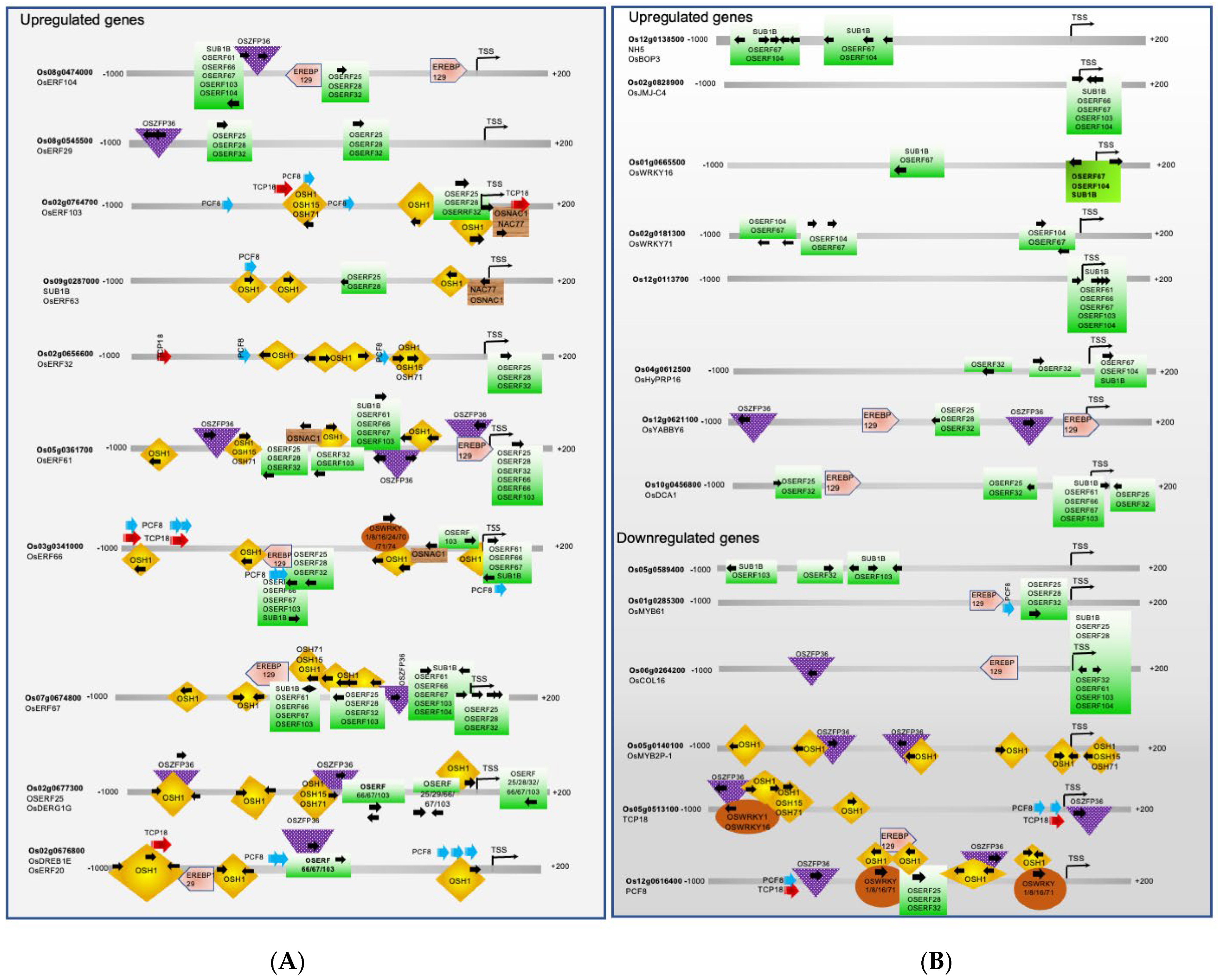{kind=link}
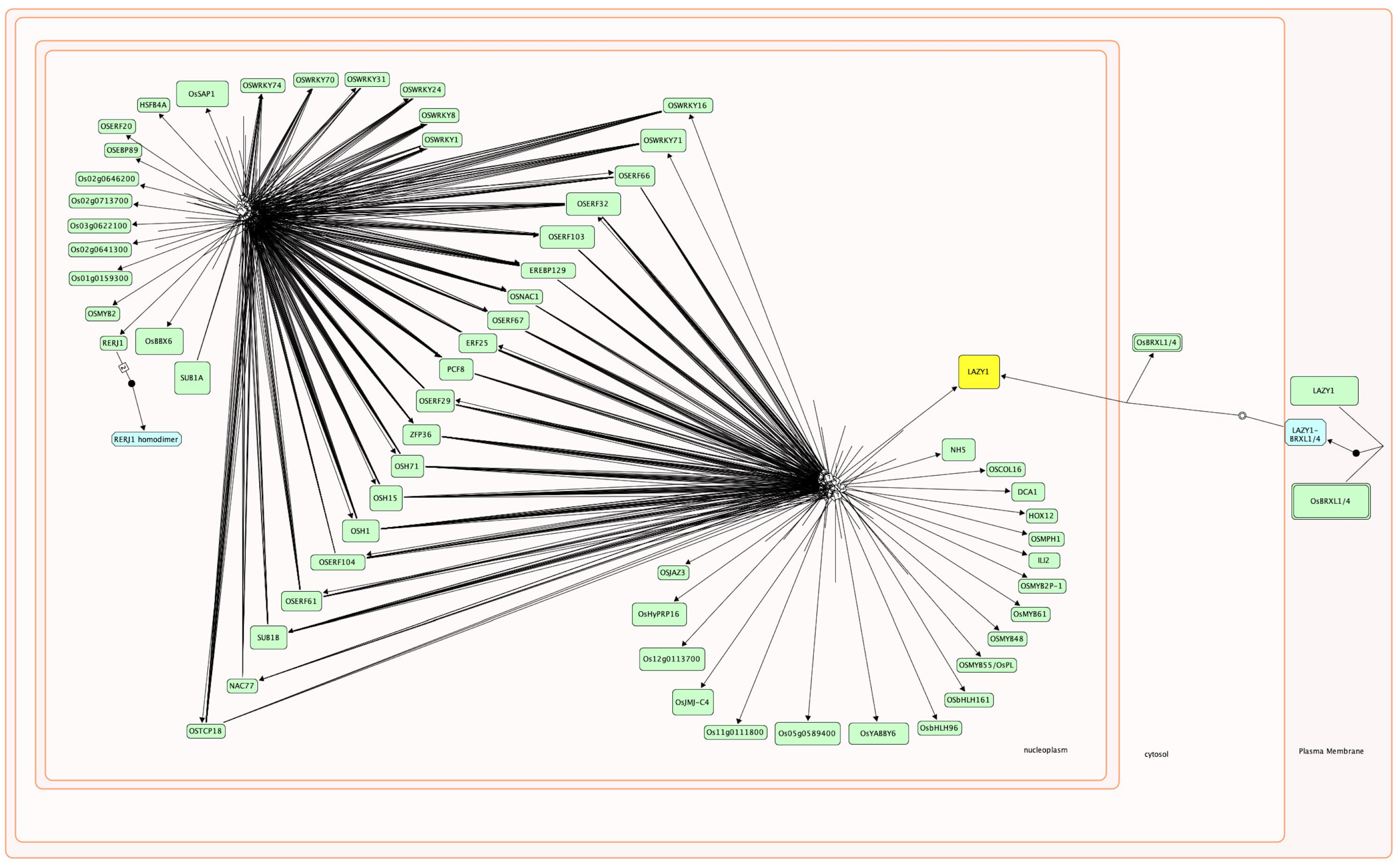{kind=link}
| RAP Gene ID and Gene Symbols | UniProt Protein ID | Gene Summary |
|---|---|---|
| TFs involved in shoot apical meristem, and early embryonic development | ||
| Os03g0727000 * OSH1, Oskn1, | P46609 | Oryza sativa Homeobox protein 1 (OSH1) plays a key role in the development and maintenance of the shoot apical meristem (SAM) [44,45,46] as well as in morphogenetic processes throughout plant development [47,48]. It is involved in the regionalization of cell identity and expresses before the development of SAM in the mature embryo. During postembryonic organ development, SAM differentiates into all aerial organs, such as leaves, stems, and flowers. The expression of the OSH1 gene is autoregulated to maintain SAM [49]. It is upregulated in response to submergence during seed germination and coleoptile elongation (this study). |
| Os07g0129700 * OSH15, Oskn3, OsKNOX15, HOS3, | O80416 | Oryza sativa Homeobox protein 15 (OSH15) is involved in shoot formation, internode development, repression of lignin biosynthesis, and control of seed shattering [50]. Loss-of-function mutants of OSH15 have a d6-type dwarf phenotype [51]. It was upregulated in response to submergence during seed germination and coleoptile elongation (this study). |
| Os05g0129700 OSH71, Oskn2, HOS9 | Q7GDL5 | Oryza sativa Homeobox protein 71 (OSH71) is involved in shoot formation during embryogenesis [47,52]. Its ectopic expression induced defects in panicle branching, internode elongation, and leaf patterning [53]. It was upregulated in response to submergence during seed germination and coleoptile elongation [12]. |
| TFs involved in plant growth and development, shoot architecture, and gravitropism | ||
| Os12g0138500 *$ NH5, OsBOP3 | Q2QXZ2 | NPR1 Homolog 5 (NH5), also known as Blade-On-Petiole3 (OsBOP3), controls the shape of the first leaf and ligule formation in rice [54]. OsBOP1, OsBOP2, and OsBOP3 functions are redundant in rice: these TFs promote leaf sheath differentiation while repressing blade differentiation. OsBOP1 and OsBOP2/3 expression was highest in the first leaf primordia, in which blade differentiation is strongly inhibited, and show decrease in the second leaf primordium [54]. |
| Os10g0456800 $ DCA1 | Q337P2 | DST Co-Activator 1 (DCA1) is upregulated in response to submergence during seed germination and coleoptile growth [12]. Previous studies have shown that DCA1 and Drought and Salt Tolerance (DST) form a hetero-tetrameric transcriptional complex that promotes stomatal opening in the guard cells and promotes transpiration, thus, negatively regulating drought and salt tolerance [55]. |
| Os12g0621100 *$ OsYABBY6 | Q2QM17 | OsYABBY6 is a member of the plant-specific transcription factor family. It regulates the initiation and development of the leaf blade (the development of vascular bundles, mestome sheath, and sclerenchyma) by suppressing the expression of meristem-specific genes in leaf primordia [56,57]. It is upregulated in response to submergence during seed germination and coleoptile elongation (this study). |
| Os06g0670300 *$ OsMPH1, OsMYB45 | A0A0P0X0C0 | The Rice MYB-like gene of Plant Height 1(OsMPH1) acts as a positive regulator of plant height by elongating internodal cell length and cell wall synthesis. It shows extremely high expression in the leaves (sheaths and especially in the pulvinus), and low expression in the internodes and spikes [58]. It also promotes large panicle size and grain yield [58]; and plays a key role in the tolerance to Cd stress in rice [59]. It is upregulated in response to submergence during seed germination and coleoptile elongation (this study). |
| Os03g0198600 $ OsHox12, HOX12 | Q10QF2 | Homeobox gene 12 (HOX12) is a negative regulator of internode growth. HOX12 positively regulates transcription of Elongated Uppermost Internode1 (EUI1) by directly binding to the EUI1 promoter [60]. EUI1 is a GA-deactivating enzyme. Inactivation of HOX12 or EUI1 results in higher GA4 levels in the uppermost internode, promoting cell division and elongation. The HOX12 RNAi plants and eui1 knockdown mutants show similar phenotype: enhancement in panicle exsertion due to elongation of the uppermost internode. |
| Os06g0264200 *$ OsCOL16, OsBBX17 | Q0DD26 | Constans-like 16 (OsCOL16) shows higher expression in vegetative tissues than in reproductive tissues. In the vegetative stage, it pro-motes plant height and delays flowering. During the reproductive stage, it promotes grain yield [61]. It is downregulated in response to submergence during seed germination and coleoptile elongation in sub-mergence-tolerant rice genotypes (this study). |
| Os02g0713700 *$ | Q6ZFU2 | Os02g0713700 is a DUF296 domain-containing TF [61]. It shows differential expression in response to imbalanced Carbon: Nitrogen availabilities [62]. It is upregulated in response to submergence during seed germination and coleoptile growth (this study). |
| Os01g0285300 *$ MYB61, OsMYB61 | Q9AQV2 | OsMYB61 is involved in the regulation of cellulose synthesis, nitrogen assimilation, carbon fixation, and growth. It works in the same pathway as Growth-Regulating Factor 4 (GRF4) [63]. It connects Carbon and Nitrogen metabolism in rice [63]. During seed germination and coleoptile growth, OSMYB61 is downregulated in response to submergence in tolerant rice genotypes (this study). |
| Os05g0140100 $ OsMYB2P-1 | Q688D6 | MYB2 phosphate-responsive gene 1 (OsMYB2P-1) regulates phosphate-starvation response and root architecture in rice [64]. It is downregulated in response to submergence during seed germination and coleoptile growth in the tolerant rice genotypes (this study). |
| Os11g0490600 *$ OsLAZY1 | Q2R435 | OsLAZY1 regulates shoot gravitropism and tiller angle through negative regulation of basipetal polar auxin transport (PAT) and positive regulation of lateral auxin transport (LAT) that results in enhancing vertical shoot growth [65,66,67,68,69,70]. OsLAZY1 is mainly expressed in gravity-sensitive shoot tissues such as coleoptiles, leaf sheath pulvini, and lamina joints [67] and less expressed in roots. It is expressed specifically in the cells at the inner side of the vascular bundles of young leaf sheaths and peripheral cylinders of vascular bundles in the unelongated stems [66]. It is upregulated in response to submergence during seed germination and coleoptile growth [12]. |
| Os11g0603000 *$ ILI2 | Q2R1J3 | Increased Leaf Inclination 2 (ILI2), a bHLH transcription factor, is mainly expressed in the lamina joint during leaf development and negatively regulates leaf angle. The lc2 mutants have enlarged leaf angles. It is induced by ABA, GA, auxin, and BR [71]. Furthermore, it is upregulated in response to submergence during seed germination and coleoptile growth (this study). |
| TFs involved in response to submergence and other abiotic and biotic stress conditions | ||
| Os09g0287000 * Sub1B, EREBP166, OsERF#063, OsERF63 | Q6EN65 | Submergence-1B (Sub1B) is a member of the ERF family. It is induced by drought, salinity, and submergence in rice seedlings [72]. It is upregulated in response to submergence during seed germination and coleoptile growth (this study). |
| Os03g0341000 * OsERF66, EREBP30 | Q10LN8 | OsERF66 is a direct transcriptional target of Sub1A in some submergence-tolerant indica rice genotypes. Sub1A, OsERF66, and OsERF67 form a regulatory cascade, and both OsERF66 and OsERF67 are substrates of the N-end rule pathway and promote submergence tolerance of rice seedlings [31,73]. OsERF66 is upregulated in response to submergence during seed germination and coleoptile growth (this study). |
| Os07g0674800 * OsERF67 | Q69J87 | OsERF67 is a direct target of Sub1A. Together with OsERF66, it functions downstream of Sub1A to form a regulatory cascade in response to submergence stress. It is a substrate of the N-end rule pathway and enhances transcriptional response to increasing submergence survival in rice [31,73]. In addition, it is upregulated in response to submergence during seed germination and coleoptile growth (this study). |
| Os05g0361700 * OsERF61, EREBP93 | Q6L4M2 | OsERF61 regulates rice growth and metabolism in response to abiotic stresses [74]. It is involved in the OsDRAP1-mediated regulation of transcription in response to salt stress [72]. It is upregulated in response to submergence during seed germination and coleoptile growth (this study). |
| Os02g0656600 OsERF32, OsERF032, EREBP21 | Q6H6G7 | OsERF32 is induced by drought, high salt, high temperature, and in response to submergence during seed germination and coleoptile growth [12,75,76,77]. |
| Os02g0677300 OsDREB1G, OsERF25, EREBP138 | Q6EP77 | Dehydration-Responsive Element-Binding 1G (OsDREB1G) is involved in both abiotic and biotic stress tolerance in rice [12,76,78,79,80,81]. It expresses in leaf sheath, blade, internode, and roots [78]. It binds to the promoters containing DRE elements to activate stress-responsive genes [76]. In addition, its expression is upregulated in response to submergence during seed germination and coleoptile growth [12]. |
| Os08g0545500 * OsERF28, OsERF29, OsDERF4, EREBP161 | Q0J3Y6 | OsERF29 is in a co-expression network of TFs which enhance drought, cold, and salt tolerance in rice [76,78,81]. Its expression reaches the highest level at 18 h after seed imbibition, and then decreases gradually [82]. Its expression is upregulated in response to submergence during seed germination and coleoptile growth (this study). |
| Os02g0764700 * OsERF103, OsDERF5, EREBP130 | Q6Z7P9 | OsERF103 negatively regulates ethylene biosynthesis and drought stress tolerance in rice. Its expression is also upregulated at the reproductive stage in rice [83]; and in response to submergence during seed germination and coleoptile growth (this study). |
| Os08g0474000 * EREBP152, OsERF104, OsDERF3, | Q0J525 | OsERF104 is known to be regulated by OsNAC45 in response to salt [84]. It is upregulated in response to submergence during seed germination and coleoptile growth (this study). |
| Os02g0676800 *$ OsERF20 OsDREB1E | Q6EP81 | OsERF20 positively regulates chilling tolerance in rice seedlings by controlling ROS scavenging and reducing cell death [81]. It is also upregulated in response to submergence during seed germination and coleoptile growth (this study). |
| Os01g0141000 * EREBP129, OsRAV2, OsRAV9, OsTEM1 | Q9AWS0 | Ethylene-Responsive Element Binding Protein 129 (EREBP129) is regulated in response to abiotic and biotic stresses [85,86,87]. It acts as a repressor of photoperiodic flowering in rice [85]. It is upregulated in response to submergence during rice seed germination and elongation of the coleoptile; and it is likely to regulate the expression of OSH1 and many other TF genes (this study). |
| Os03g0437200 ZFP36, OsBSRD1, OsDLN91 | Q75KE5 | Zinc Finger Protein 36 (ZFP36) regulates ABA-mediated abiotic stress response through reactive oxygen species signaling and promotes oxidative stress tolerance [88]. It is upregulated in response to submergence during seed germination and coleoptile growth [12]. |
| Os02g0579000 * OsNAC1, ONAC27, OMTN1, OsDLN60 | Q0E046 | Rice NAC domain-containing protein 1 (OsNAC1) shows higher expression in the stamen, leave blade, embryo, root, and panicle. Its expression is downregulated in response to drought; and upregulated in response to cold stress and ABA treatment [89] and in response to submergence during seed germination and coleoptile growth (this study). |
| Os12g0123800 * NAC77, NAC077, ONAC300, OsNAC132, OsDLN250 | Q5CD17 | Rice NAC domain-containing transcription factor 77 (NAC77) expresses at early developmental stages in the shoot, root, flower, and the mature phloem of vascular tissues [90]. It is upregulated in response to both abiotic (salt, drought, and cold) and biotic stresses in rice [91,92]. In addition, it is upregulated in response to submergence during seed germination (this study). |
| Os03g0182800 *$ OsEBP89 | Q7XBH8 | Ethylene-responsive element binding protein 89 (OsEBP89) is a negative regulator of ABA-dependent stress responses [93]. Its expression is strongly inhibited by drought stress and stimulated by submergence stress in the root and meristem [93]. We find that it is upregulated during seed germination and coleoptile elongation under submergence rice (this study). OsEBP89 knockout mutant showed improved seed germination rate under submerged conditions and, thus, enhances direct seeding on wetlands and confers drought tolerance in rice [93]. OsEBP89 is phosphorylated by sucrose non-fermenting-1-related protein kinase-1 (OsSnRK1α) [93]. |
| Os01g0246700 * OsWRKY1 | Q0JP37 | OsWRKY1 is upregulated in response to submergence during seed germination and coleoptile growth (this study). |
| Os05g0583000 OsWRKY8 | Q75HY3 | OsWRKY8 is induced by PEG, NaCl, ABA, and naphthalene acetic acid in rice. It is upregulated in response to submergence during seed germination and coleoptile growth [12]. |
| Os01g0665500 * OsWRKY16 | Q0JKL7 | OsWRKY16 is upregulated in response to submergence during seed germination and coleoptile growth (this study). |
| Os01g0826400 * OsWRKY24 | Q6IEQ7 | OsWRKY24 acts as both a transcriptional repressor and an activator. It is expressed chiefly in aleurone cells and embryos within the seeds [94] and shows low expression in leaves, roots, and panicles [95]. Its expression is induced by ABA and JA but repressed by GA [94,96]. It is upregulated in response to submergence during seed germination and coleoptile growth (this study). |
| Os03g0321700 * OsWRKY31, OsWRKY55 | Q10M65 | OsWRKY31 is a component of the auxin signaling pathways and is also involved in the defense response in rice [97]. It is upregulated in response to submergence during seed germination and coleoptile growth (this study). |
| Os05g0474800: OsWRKY70 | Q65WW1 | OsWRKY70, together with OsWRKY24, OsWRKY53 acts as a negative regulator of both GA and ABA signaling [96]. OsWRKY70 positively regulates early defense signaling by inducing the synthesis of the volatile indole [98]. It is also upregulated in response to submergence during seed germination and coleoptile growth [12]. |
| Os02g0181300 * OsWRKY71 | Q6QHD1 | OsWRKY71 expression is induced by SA, JA, 1-aminocyclo-propane-1-carboxylic acid, wounding, and pathogen infection. It is likely to function as a transcriptional regulator of OsNPR1 and OsPR1b in rice defense signaling pathways [99]. |
| Os09g0334500 * OsWRKY74 | Q6ERI5 | OsWRKY74 expresses in roots and leaves. It is involved in the tolerance to phosphate starvation and shows differential expression in response to iron and nitrogen deficiencies, and cold stress [100]. It is upregulated in response to submergence during rice seed germination and coleoptile growth (this study). |
| Os03g0315400 *$ OsMYB2 | Q10MB4 | OsMYB2 is a positive regulator of salt, cold, and dehydration tolerance in rice [101]. Overexpression of OsMYB2 leads to the accumulation of soluble sugars and proline, and a decline in the levels of H2O2 and malondialdehyde. OsMYB2 overexpressing plants also showed upregulation of stress-related genes, including OsLEA3, OsRab16A, and OsDREB2A [101]. In addition, it is upregulated in response to submergence during seed germination and coleoptile growth in rice (this study). |
| Os05g0553400 $ OsMYB55 OsPL9 | Q6I634 | OsMYB55 confers heat tolerance in rice by increasing the biosynthesis of glutamic acid, proline, arginine, and GABA [102]. Under heat stress, rice coleoptiles that overexpressed OsMYB55 became longer [102] and showed higher plant growth and grain yields than the wild type. It also enhances anthocyanin biosynthesis in rice [103]. In addition, it is upregulated in response to submergence during seed germination and coleoptile growth [12]. |
| Os01g0975300 *$ OsMYB48, OsMYB48-1, OsMYB48-2 | Q0JFK3 | OsMYB48 is strongly induced by polyethylene glycol, ABA, H2O2 and dehydration, and slightly induced by high salinity and cold. It is a positive regulator of drought and salinity tolerance [104]. It is upregulated in response to submergence during seed germination and coleoptile growth in rice [12]. |
| Os02g0641300 *$ OsMYB21 | Q0DZ73 | Os02g0641300/OsMYB21 is upregulated in response to chilling stress [105]. Also, it is induced in response to submergence during seed germination and coleoptile growth in rice (this study). |
| Os05g0589400 *$ | Q6I5E6 | Os05g0589400 encodes a R2R3MYB transcription factor. It is downregulated in response to submergence during seed germination and coleoptile growth in submergence-tolerant rice genotypes (this study). |
| Os08g0471000 $ HSFB4A, OsHsf-20 | Q6Z9R8 | Heat stress transcription factor B-4a (HSFB4A) could act as a seed-specific transcriptional repressor co-activator in rice [106,107,108]. It is likely to form homotrimer. It is upregulated in response to submergence during seed germination and coleoptile growth [12]. |
| Os03g0188400 $ OsbHLH96 | Q8H7N8 | Rice basic helix-loop-helix 96 (OsbHLH96) forms the OsbHLH96-OsHLH61 complex that regulates PR genes in response to brown planthopper attack and is likely to be involved in mediating crosstalk between SA and JA signaling. OsbHLH96 could also interact with OsJAZ3 [109]. In addition, OsbHLH96 is upregulated in response to submergence during seed germination and coleoptile growth [12]. |
| Os11g0111800 *$ OsbHLH160 | A3C7W7 | OsbHLH160 expresses in rice roots [110]. It is upregulated in response to submergence during seed germination and coleoptile growth (this study). |
| Os12g0111400 *$ OsbHLH161 | Q2QYP2 | OsbHLH161 [111] is upregulated in response to submergence during seed germination and coleoptile growth (this study). |
| Os04g0301500 *$ OsbHLH6, RERJ1 | Q0JEB7 | OsbHLH6 is induced by JA, wounding, drought, and Pi deficiency [112,113]. It acts as a positive regulator for Pi signaling and homeostasis. It is also upregulated in response to submergence during seed germination and coleoptile growth (this study). |
| Os09g0486500 $ OsSAP1, ISAP1 | A3C039 | Rice Stress Associated 1 (OsSAP1) expression is induced by cold [114] and submergence stress during seed germination and coleoptile growth [12]. It regulates biotic and abiotic stress responses [115]. |
| Os04g0612500 *$ OsHyPRP16 | B9FCG2 | Hybrid Proline-Rich Protein 16 (OsHyPRP16) is upregulated in response to M. oryzae infection [116]. It is also upregulated in response to submergence stress during seed germination and coleoptile growth (this study). |
| Os01g0159300 *$ | Q0JQI1 | Os01g0159300 is upregulated in response to submergence during seed germination and coleoptile growth (this study). |
| Os02g0646200 $ OsBBX6 | Q6H630 | The Rice B-box domain containing 6 (OsBBX6) is a member of the Zinc Finger TF family [117]. It is upregulated in response to submergence during seed germination and coleoptile elongation [12]. |
| Os12g0113700 $ | A0A0P0Y689 | Os12g0113700 is a Zinc finger C3HC4 type TF. It is also upregulated in response to submergence during seed germination and coleoptile growth [12]. |
| Os02g0828900 *$ OsJMJ-C4 | Q6K7P0 | OsJMJ-C4 (Jumonji C domain-containing protein 4) is likely to function as histone demethylases or a TF involved in the epigenetic regulation of plant development [118,119]. It is upregulated in response to submergence during seed germination and coleoptile growth (this study). |
| Os08g0428400 $ OsJAZ3, TIFY6a | Q6ZJU3 | OsJAZ3 repressor is induced by ABA, drought, and salt stresses. It is targeted for degradation by the SCF (COI1) E3 ubiquitin ligase–proteasome pathway during JA signaling [120,121]. It is upregulated in response to submergence during seed germination and coleoptile growth [12]. |
| Os03g0622100 *$ | Q10GM4 | Os03g0622100, a B3 family TF, is upregulated in response to submergence during rice seed germination and coleoptile growth (this study). |
| Os12g0616400 * PCF8 | Q2QM59 | Proliferating Cell Factor 8 (PCF8) acts as a negative regulator of cold tolerance in rice, and its expression is downregulated by miR319 [122]. Furthermore, it is downregulated in response to submergence during seed germination and coleoptile growth in the tolerant genotypes (this study). |
| Os05g0513100 * OsTCP18, TCP18 | Q5TKH1 | Teosinte branched 1, Cycloidea and Proliferating 18 (TCP 18) could act as transcriptional activators or repressors [123]. However, it is downregulated in submergence-tolerant genotypes of rice in response to submergence during seed germination and coleoptile growth (this study). |
2.2.1. Transcription Factors involved in Shoot Apical Meristem Maintenance, and Coleoptile Elongation
2.2.2. Transcription Factors Involved in the Shoot, Leaf and Flower Development, and Gravitropism
2.2.3. TFs Involved in Stress Response during Seed Germination under Submergence in Rice
2.3. Plant Reactome Knowledgebase Supports the Integration of Heterogeneous OMICs Datasets for Plant Pathways In Silico Modeling
3. Discussion
4. Materials and Methods
4.1. Identification of 57 Transcription Regulators with Strong Co-Expression Pattern during Seed Germination under Submergence
4.2. Promoter Analysis
4.3. Biocuration of a Transcription Factor Network
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Naithani, S.; Dikeman, D.; Garg, P.; Al-Bader, N.; Jaiswal, P. Beyond Gene Ontology (Go): Using Biocuration Approach to Improve the Gene Nomenclature and Functional Annotation of Rice S-Domain Kinase Subfamily. PeerJ 2021, 9, e11052. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, B.; Kitazumi, A.; Cheung, C.M.; Lakshmanan, M.; Reyes, B.G.d.L.; Jang, I.-C.; Lee, D.-Y. Identification of Candidate Network Hubs Involved in Metabolic Adjustments of Rice under Drought Stress by Integrating Transcriptome Data and Genome-Scale Metabolic Network. Plant Sci. 2016, 242, 224–239. [Google Scholar] [CrossRef] [PubMed]
- Seo, Y.-S.; Chern, M.; Bartley, L.; Han, M.; Jung, K.H.; Lee, I.; Walia, H.; Richter, T.; Xu, X.; Cao, P.; et al. Towards Establishment of a Rice Stress Response Interactome. PLoS Genet. 2011, 7, e1002020. [Google Scholar] [CrossRef]
- Lee, I.; Seo, Y.-S.; Coltrane, D.; Hwang, S.; Oh, T.; Marcotte, E.M.; Ronald, P.C. Genetic Dissection of the Biotic Stress Response Using a Genome-Scale Gene Network for Rice. Proc. Natl. Acad. Sci. USA 2011, 108, 18548–18553. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.P.; Serra, T.; Figueiredo, D.D.; Barros, P.; Lourenço, T.; Chander, S.; Oliveira, M.M.; Saibo, N.J. Transcription Regulation of Abiotic Stress Responses in Rice: A Combined Action of Transcription Factors and Epigenetic Mechanisms. Omics 2011, 15, 839–857. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Soto, A.; Echeverría-Beirute, F.; Abdelnour-Esquivel, A.; Valdez-Melara, M.; Boch, J.; Gatica-Arias, A. Rice Breeding in the New Era: Comparison of Useful Agronomic Traits. Curr. Plant Biol. 2021, 27, 100211. [Google Scholar] [CrossRef]
- Bailey-Serres, J.; Voesenek, L.A.C.J. Life in the Balance: A Signaling Network Controlling Survival of Flooding. Curr. Opin. Plant Biol. 2010, 13, 489–494. [Google Scholar] [CrossRef]
- Yu, S.M.; Lee, H.T.; Lo, S.F.; Ho, T.H.D. How Does Rice Cope with Too Little Oxygen During Its Early Life? New Phytol. 2021, 229, 36–41. [Google Scholar] [CrossRef]
- Jang, S.-G.; Kim, B.; Kim, Y.; Kwon, S.-W. Genome-Wide Association Study of Submergence Tolerance in Rice (Oryza sativa L.). Plant Breed. Biotechnol. 2023, 11, 25–33. [Google Scholar] [CrossRef]
- Ma, M.; Cen, W.; Li, R.; Wang, S.; Luo, J. The Molecular Regulatory Pathways and Metabolic Adaptation in the Seed Germination and Early Seedling Growth of Rice in Response to Low O2 Stress. Plants 2020, 9, 1363. [Google Scholar] [CrossRef]
- Septiningsih, E.M.; Pamplona, A.M.; Sanchez, D.L.; Neeraja, C.N.; Vergara, G.V.; Heuer, S.; Ismail, A.M.; Mackill, D.J. Development of Submergence-Tolerant Rice Cultivars: The Sub1 Locus and Beyond. Ann. Bot. 2009, 103, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.-K.; Tung, C.-W. RNA-Seq Analysis of Diverse Rice Genotypes to Identify the Genes Controlling Coleoptile Growth during Submerged Germination. Front. Plant Sci. 2017, 8, 762. [Google Scholar] [CrossRef] [PubMed]
- Alam, R.; Hummel, M.; Yeung, E.; Locke, A.M.; Ignacio, J.C.I.; Baltazar, M.D.; Jia, Z.; Ismail, A.M.; Septiningsih, E.M.; Bailey-Serres, J. Flood Resilience Loci Submergence 1 and Anaerobic Germination 1 Interact in Seedlings Established Underwater. Plant Direct 2020, 4, e00240. [Google Scholar] [CrossRef]
- Ismail, A.M.; Ella, E.S.; Vergara, G.V.; Mackill, D.J. Mechanisms Associated with Tolerance to Flooding during Germination and Early Seedling Growth in Rice (Oryza sativa). Ann. Bot. 2009, 103, 197–209. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.; Liu, G.; Monje, O.; Stutte, G.; Porterfield, D. Induction of Hypoxic Root Metabolism Results from Physical Limitations in O2 Bioavailability in Microgravity. Adv. Space Res. 2004, 34, 1579–1584. [Google Scholar] [CrossRef] [PubMed]
- Stout, S.C.; Porterfield, D.M.; Briarty, L.G.; Kuang, A.; Musgrave, M.E. Evidence of Root Zone Hypoxia in Brassica rapa L. Grown in Microgravity. Int. J. Plant Sci. 2001, 162, 249–255. [Google Scholar] [CrossRef]
- Manzano, A.; Villacampa, A.; Sáez-Vásquez, J.; Kiss, J.Z.; Medina, F.J.; Herranz, R. The Importance of Earth Reference Controls in Spaceflight Omics Research: Characterization of Nucleolin Mutants from the Seedling Growth Experiments. iScience 2020, 23, 101686. [Google Scholar] [CrossRef]
- Wang, X.; Komatsu, S. The Role of Phytohormones in Plant Response to Flooding. Int. J. Mol. Sci. 2022, 23, 6383. [Google Scholar] [CrossRef]
- Jayawardhane, J.; Wijesinghe, M.K.P.S.; Bykova, N.V.; Igamberdiev, A.U. Metabolic Changes in Seed Embryos of Hypoxia-Tolerant Rice and Hypoxia-Sensitive Barley at the Onset of Germination. Plants 2021, 10, 2456. [Google Scholar] [CrossRef]
- Nghi, K.N.; Tagliani, A.; Mariotti, L.; Weits, D.A.; Perata, P.; Pucciariello, C. Auxin Is Required for the Long Coleoptile Trait in Japonica Rice under Submergence. New Phytol. 2021, 229, 85–93. [Google Scholar] [CrossRef]
- Waller, F.; Furuya, M.; Nick, P. OsARF1, An Auxin Response Factor from Rice, Is Auxin-Regulated And Classifies As A Primary Auxin Responsive Gene. Plant Mol. Biol. 2002, 50, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Huang, L.; Gao, J.; Wang, Y. The Molecular Mechanism of Plant Gravitropism. Hereditas 2016, 38, 589–602. [Google Scholar] [PubMed]
- Sims, K.; Abedi-Samakush, F.; Szulc, N.; Honti, M.G.M.; Mattsson, J. OsARF11 Promotes Growth, Meristem, Seed, and Vein Formation during Rice Plant Development. Int. J. Mol. Sci. 2021, 22, 4089. [Google Scholar] [CrossRef]
- Wu, Y.-S.; Yang, C.-Y. Comprehensive Transcriptomic Analysis of Auxin Responses in Submerged Rice Coleoptile Growth. Int. J. Mol. Sci. 2020, 21, 1292. [Google Scholar] [CrossRef] [PubMed]
- Nagai, K.; Mori, Y.; Ishikawa, S.; Furuta, T.; Gamuyao, R.; Niimi, Y.; Hobo, T.; Fukuda, M.; Kojima, M.; Takebayashi, Y.; et al. Antagonistic Regulation of the Gibberellic Acid Response during Stem Growth in Rice. Nature 2020, 584, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Naithani, S.; Nonogaki, H.; Jaiswal, P. Exploring Crossroads between Seed Development and Stress-Response. In Mechanism of Plant Hormone Signaling under Stress; Pandey, G.K., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2017; pp. 415–454. [Google Scholar]
- Xiong, Q.; Ma, B.; Lu, X.; Huang, Y.-H.; He, S.-J.; Yang, C.; Yin, C.-C.; Zhao, H.; Zhou, Y.; Zhang, W.-K.; et al. Ethylene-Inhibited Jasmonic Acid Biosynthesis Promotes Mesocotyl/Coleoptile Elongation of Etiolated Rice Seedlings. Plant Cell 2017, 29, 1053–1072. [Google Scholar] [CrossRef]
- Yin, C.; Huang, Y.; Zhang, X.; Zhou, Y.; Chen, S.; Zhang, J. Ethylene-Mediated Regulation of Coleoptile Elongation in Rice Seedlings. Plant Cell Envion. 2023, 46, 1060–1074. [Google Scholar] [CrossRef]
- Xu, K.; Xu, X.; Fukao, T.; Canlas, P.; Maghirang-Rodriguez, R.; Heuer, S.; Ismail, A.M.; Bailey-Serres, J.; Ronald, P.C.; Mackill, D.J. Sub1a Is an Ethylene-Response-Factor-Like Gene That Confers Submergence Tolerance to Rice. Nature 2006, 442, 705–708. [Google Scholar] [CrossRef]
- Hattori, Y.; Nagai, K.; Furukawa, S.; Song, X.-J.; Kawano, R.; Sakakibara, H.; Wu, J.; Matsumoto, T.; Yoshimura, A.; Kitano, H.; et al. The Ethylene Response Factors Snorkel1 and Snorkel2 Allow Rice to Adapt to Deep Water. Nature 2009, 460, 1026–1030. [Google Scholar] [CrossRef]
- Lin, C.-C.; Chao, Y.-T.; Chen, W.-C.; Ho, H.-Y.; Chou, M.-Y.; Li, Y.-R.; Wu, Y.-L.; Yang, H.-A.; Hsieh, H.; Lin, C.-S.; et al. Regulatory Cascade Involving Transcriptional and N-End Rule Pathways in Rice under Submergence. Proc. Natl. Acad. Sci. USA 2019, 116, 3300–3309. [Google Scholar] [CrossRef]
- Hussain, S.; Yin, H.; Peng, S.; Khan, F.A.; Khan, F.; Sameeullah, M.; Hussain, H.A.; Huang, J.; Cui, K.; Nie, L. Comparative Transcriptional Profiling of Primed and Non-primed Rice Seedlings under Submergence Stress. Front. Plant Sci. 2016, 7, 1125. [Google Scholar] [CrossRef] [PubMed]
- Naithani, S.; Komath, S.S.; Nonomura, A.; Govindjee, G. Plant Lectins and Their Many Roles: Carbohydrate-Binding and Beyond. J. Plant Physiol. 2021, 266, 153531. [Google Scholar] [CrossRef]
- Naithani, S.; Gupta, P.; Preece, J.; D’Eustachio, P.; Elser, J.L.; Garg, P.; Dikeman, A.D.; Kiff, J.; Cook, J.; Olson, A.; et al. Plant Reactome: A Knowledgebase and Resource for Comparative Pathway Analysis. Nucleic Acids Res. 2020, 48, D1093–D1103. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Naithani, S.; Tello-Ruiz, M.K.; Chougule, K.; D’eustachio, P.; Fabregat, A.; Jiao, Y.; Keays, M.; Lee, Y.K.; Kumari, S.; et al. Gramene Database: Navigating Plant Comparative Genomics Resources. Curr. Plant Biol. 2016, 7, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Naithani, S.; Preece, J.; Kim, S.; Cheng, T.; D’eustachio, P.; Elser, J.; Bolton, E.E.; Jaiswal, P. Plant Reactome and PubChem: The Plant Pathway and (Bio)Chemical Entity Knowledgebases; Springer: New York, NY, USA, 2022; Volume 2443, pp. 511–525. [Google Scholar] [CrossRef]
- Naithani, S.; Preece, J.; D’Eustachio, P.; Gupta, P.; Amarasinghe, V.; Dharmawardhana, P.D.; Wu, G.; Fabregat, A.; Elser, J.L.; Weiser, J.; et al. Plant Reactome: A Resource for Plant Pathways and Comparative Analysis. Nucleic Acids Res. 2017, 45, D1029–D1039. [Google Scholar] [CrossRef]
- Sakai, H.; Lee, S.S.; Tanaka, T.; Numa, H.; Kim, J.; Kawahara, Y.; Wakimoto, H.; Yang, C.-C.; Iwamoto, M.; Abe, T.; et al. Rice Annotation Project Database (RAP-DB): An Integrative and Interactive Database for Rice Genomics. Plant Cell Physiol. 2013, 54, e6. [Google Scholar] [CrossRef]
- Tello-Ruiz, M.K.; Naithani, S.; Gupta, P.; Olson, A.; Wei, S.; Preece, J.; Jiao, Y.; Wang, B.; Chougule, K.; Garg, P.; et al. Gramene 2021: Harnessing the Power of Comparative Genomics and Pathways for Plant Research. Nucleic Acids Res. 2021, 49, D1452–D1463. [Google Scholar] [CrossRef]
- Mohanty, B. Promoter Architecture and Transcriptional Regulation of Genes Upregulated in Germination and Coleoptile Elongation of Diverse Rice Genotypes Tolerant to Submergence. Front. Genet. 2021, 12, 639654. [Google Scholar] [CrossRef]
- Gupta, P.; Geniza, M.; Naithani, S.; Phillips, J.L.; Haq, E.; Jaiswal, P. Chia (Salvia hispanica) Gene Expression Atlas Elucidates Dynamic Spatio-Temporal Changes Associated With Plant Growth and Development. Front. Plant Sci. 2021, 12, 667678. [Google Scholar] [CrossRef]
- Song, Q.; Lee, J.; Akter, S.; Rogers, M.; Grene, R.; Li, S. Prediction of Condition-Specific Regulatory Genes Using Machine Learning. Nucleic Acids Res. 2020, 48, 19. [Google Scholar] [CrossRef]
- Song, Q.; Grene, R.; Heath, L.S.; Li, S. Identification of Regulatory Modules in Genome Scale Transcription Regulatory Networks. BMC Syst. Biol. 2017, 11, 140. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Hong, S.K.; Tagiri, A.; Kitano, H.; Yamamoto, N.; Nagato, Y.; Matsuoka, M. A Rice Homeobox Gene, OSH1, Is Expressed before Organ Differentiation in a Specific Region during Early Embryogenesis. Proc. Natl. Acad. Sci. USA 1996, 93, 8117–8122. [Google Scholar] [CrossRef]
- Vollbrecht, E.; Reiser, L.; Hake, S. Shoot Meristem Size Is Dependent on Inbred Background and Presence of the Maize Homeobox Gene, knotted1. Development 2000, 127, 3161–3172. [Google Scholar] [CrossRef] [PubMed]
- Hay, A.; Tsiantis, M. Knox Genes: Versatile Regulators of Plant Development and Diversity. Development 2010, 137, 3153–3165. [Google Scholar] [CrossRef]
- Sentoku, N.; Sato, Y.; Kurata, N.; Ito, Y.; Kitano, H.; Matsuoka, M. Regional Expression of the Rice Kn1-Type Homeobox Gene Family During Embryo, Shoot, and Flower Development. Plant Cell 1999, 11, 1651–1664. [Google Scholar] [CrossRef] [PubMed]
- Hake, S.; Smith, H.M.; Holtan, H.; Magnani, E.; Mele, G.; Ramirez, J. The Role of Knox Genes in Plant Development. Annu. Rev. Cell Dev. Biol. 2004, 20, 125–151. [Google Scholar] [CrossRef]
- Tsuda, K.; Ito, Y.; Sato, Y.; Kurata, N. Positive Autoregulation of a Knox Gene Is Essential for Shoot Apical Meristem Maintenance in Rice. Plant Cell 2011, 23, 4368–4381. [Google Scholar] [CrossRef]
- Yoon, J.; Cho, L.-H.; Antt, H.W.; Koh, H.-J.; An, G. Knox Protein Osh15 Induces Grain Shattering by Repressing Lignin Biosynthesis Genes. Plant Physiol. 2017, 174, 312–325. [Google Scholar] [CrossRef]
- Sato, Y.; Sentoku, N.; Miura, Y.; Hirochika, H.; Kitano, H.; Matsuoka, M. Loss-of-Function Mutations in the Rice Homeobox Gene Osh15 Affect the Architecture of Internodes Resulting in Dwarf Plants. EMBO J. 1999, 18, 992–1002. [Google Scholar] [CrossRef]
- Postma-Haarsma, A.D.; Verwoert, I.I.; Stronk, O.P.; Koster, J.; Lamers, G.E.; Hoge, J.H.C.; Meijer, A.H. Characterization of the Knox Class Homeobox Genes Oskn2 and Oskn3 Identified in a Collection of cDNA Libraries Covering the Early Stages of Rice Embryogenesis. Plant Mol. Biol. 1999, 39, 257–271. [Google Scholar] [CrossRef]
- Postma-Haarsma, A.D.; Rueb, S.; Scarpella, E.; Besten, W.D.; Hoge, J.H.C.; Meijer, A.H. Developmental Regulation and Downstream Effects of the Knox Class Homeobox Genes Oskn2 and Oskn3 from Rice. Plant Mol. Biol. 2002, 48, 423–441. [Google Scholar] [CrossRef]
- Toriba, T.; Tokunaga, H.; Shiga, T.; Nie, F.; Naramoto, S.; Honda, E.; Tanaka, K.; Taji, T.; Itoh, J.-I.; Kyozuka, J. Blade-on-Petiole Genes Temporally and Developmentally Regulate the Sheath to Blade Ratio of Rice Leaves. Nat. Commun. 2019, 10, 619. [Google Scholar] [CrossRef]
- Cui, L.-G.; Shan, J.-X.; Shi, M.; Gao, J.-P.; Lin, H.-X. DCA1 Acts as a Transcriptional Co-activator of DST and Contributes to Drought and Salt Tolerance in Rice. PLoS Genet. 2015, 11, e1005617. [Google Scholar] [CrossRef]
- Romanova, M.A.; Maksimova, A.I.; Pawlowski, K.; Voitsekhovskaja, O.V. Yabby Genes in the Development and Evolution of Land Plants. Int. J. Mol. Sci. 2021, 22, 4139. [Google Scholar] [CrossRef] [PubMed]
- Veisi, S.; Sabouri, A.; Abedi, A. Meta-Analysis of QTLs and Candidate Genes Associated with Seed Germination in Rice (Oryza sativa L.). Physiol. Mol. Biol. Plants 2022, 28, 1587–1605. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yu, C.; Lin, J.; Liu, J.; Liu, B.; Wang, J.; Huang, A.; Li, H.; Zhao, T. Osmph1 Regulates Plant Height and Improves Grain Yield in Rice. PLoS ONE 2017, 12, e0180825. [Google Scholar] [CrossRef]
- Hu, S.; Yu, Y.; Chen, Q.; Mu, G.; Shen, Z.; Zheng, L. Osmyb45 Plays an Important Role in Rice Resistance to Cadmium Stress. Plant Sci. 2017, 264, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Fang, J.; Xu, F.; Wang, W.; Chu, C. Rice Hox12 Regulates Panicle Exsertion by Directly Modulating the Expression of Elongated Uppermost Internode1. Plant Cell 2016, 28, 680–695. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Favero, D.S.; Qiu, J.; Roalson, E.H.; Neff, M.M. Insights into the Evolution and Diversification of the AT-Hook Motif Nuclear Localized Gene Family in Land Plants. BMC Plant Biol. 2014, 14, 266. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.; Sang, Y.; Sun, W.; Fu, Y.; Yang, Z. Transcriptomic Analysis of Responses to Imbalanced Carbon: Nitrogen Availabilities in Rice Seedlings. PLoS ONE 2016, 11, e0165732. [Google Scholar] [CrossRef]
- Gao, Y.; Xu, Z.; Zhang, L.; Li, S.; Wang, S.; Yang, H.; Liu, X.; Zeng, D.; Liu, Q.; Qian, Q.; et al. Myb61 is Regulated by Grf4 and Promotes Nitrogen Utilization and Biomass Production in Rice. Nat. Commun. 2020, 11, 5219. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Wang, Y.; Yang, A.; Zhang, W.-H. OsMYB2P-1, an R2R3 MYB Transcription Factor, Is Involved in the Regulation of Phosphate-Starvation Responses and Root Architecture in Rice. Plant Physiol. 2012, 159, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Lin, Z.; Li, H.; Li, X.; Li, J.; Wang, Y.; Zhang, X.; Zhu, Z.; Zhai, W.; Wang, X.; et al. Tac1, a Major Quantitative Trait Locus Controlling Tiller Angle in Rice. Plant J. 2007, 52, 891–898. [Google Scholar] [CrossRef]
- Li, P.; Wang, Y.; Qian, Q.; Fu, Z.; Wang, M.; Zeng, D.; Li, B.; Wang, X.; Li, J. Lazy1 Controls Rice Shoot Gravitropism through Regulating Polar Auxin Transport. Cell Res. 2007, 17, 402–410. [Google Scholar] [CrossRef]
- Yoshihara, T.; Iino, M. Identification of the Gravitropism-Related Rice Gene Lazy1 and Elucidation of Lazy1-Dependent and -Independent Gravity Signaling Pathways. Plant Cell Physiol. 2007, 48, 678–688. [Google Scholar] [CrossRef]
- Sang, D.; Chen, D.; Liu, G.; Liang, Y.; Huang, L.; Meng, X.; Chu, J.; Sun, X.; Dong, G.; Yuan, Y.; et al. Strigolactones Regulate Rice Tiller Angle by Attenuating Shoot Gravitropism through Inhibiting Auxin Biosynthesis. Proc. Natl. Acad. Sci. USA 2014, 111, 11199–11204. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Hu, Y.; Tong, A.; Yan, B.; Lv, Y.; Wang, S.; Ma, W.; Cui, Z.; Wang, X. Lazy1 Controls Tiller Angle and Shoot Gravitropism by Regulating the Expression of Auxin Transporters and Signaling Factors in Rice. Plant Cell Physiol. 2021, 61, 2111–2125. [Google Scholar] [CrossRef]
- Yoshihara, T.; Spalding, E.P. Lazy Genes Mediate the Effects of Gravity on Auxin Gradients and Plant Architecture. Plant Physiol. 2017, 175, 959–969. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.-Q.; Hu, J.; Guo, L.-B.; Qian, Q.; Xue, H.-W. Rice Leaf Inclination2, a Vin3-Like Protein, Regulates Leaf Angle through Modulating Cell Division of the Collar. Cell Res. 2010, 20, 935–947. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, L.; Du, F.; Wang, J.; Zhao, X.; Li, Z.; Wang, W.; Xu, J.; Fu, B. Comparative Transcriptome and Metabolome Profiling Reveal Molecular Mechanisms Underlying Osdrap1-Mediated Salt Tolerance in Rice. Sci. Rep. 2021, 11, 5166. [Google Scholar] [CrossRef]
- Huang, C.; Wang, D.; Chen, H.; Deng, W.; Chen, D.; Chen, P.; Wang, J. Genome-Wide Identification of Duf26 Domain-Containing Genes in Dongxiang Wild Rice and Analysis of Their Expression Responses under Submergence. Curr. Issues Mol. Biol. 2022, 44, 3351–3363. [Google Scholar] [CrossRef]
- Rashid, M.; Guangyuan, H.; Guangxiao, Y.; Hussain, J.; Xu, Y. Ap2/Erf Transcription Factor in Rice: Genome-Wide Canvas and Syntenic Relationships between Monocots and Eudicots. Evol. Bioinform. Online 2012, 8, 321–355. [Google Scholar] [CrossRef] [PubMed]
- Matsukura, S.; Mizoi, J.; Yoshida, T.; Todaka, D.; Ito, Y.; Maruyama, K.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Comprehensive Analysis of Rice Dreb2-Type Genes That Encode Transcription Factors Involved in the Expression of Abiotic Stress-Responsive Genes. Mol. Genet. Genom. 2010, 283, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-Q.; Meng, X.-P.; Zhang, Y.; Xia, M.; Wang, X.-P. Over-Expression of Osdreb Genes Lead to Enhanced Drought Tolerance in Rice. Biotechnol. Lett. 2008, 30, 2191–2198. [Google Scholar] [CrossRef]
- Huang, L.; Wang, Y.; Wang, W.; Zhao, X.; Qin, Q.; Sun, F.; Hu, F.; Zhao, Y.; Li, Z.; Fu, B.; et al. Characterization of Transcription Factor Gene Osdrap1 Conferring Drought Tolerance in Rice. Front. Plant Sci. 2018, 9, 94. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.J.; Min, M.K.; Kim, J.A.; Kim, D.Y.; Yoon, I.S.; Kwon, T.R.; Byun, M.O.; Kim, B.G. Ectopic Expression of Osdreb1g, a Member of the Osdreb1 Subfamily, Confers Cold Stress Tolerance in Rice. Front. Plant Sci. 2019, 10, 297. [Google Scholar] [CrossRef]
- Sánchez-Sanuy, F.; Peris-Peris, C.; Tomiyama, S.; Okada, K.; Hsing, Y.I.; San Segundo, B.; Campo, S. Osa-Mir7695 Enhances Transcriptional Priming in Defense Responses against the Rice Blast Fungus. BMC Plant Biol. 2019, 19, 563. [Google Scholar] [CrossRef]
- Xu, K.; Chen, S.; Li, T.; Yu, S.; Zhao, H.; Liu, H.; Luo, L. Overexpression of Oshmgb707, a High Mobility Group Protein, Enhances Rice Drought Tolerance by Promoting Stress-Related Gene Expression. Front. Plant Sci. 2021, 12, 711271. [Google Scholar] [CrossRef]
- Wang, H.; Lu, S.; Guan, X.; Jiang, Y.; Wang, B.; Hua, J.; Zou, B. Dehydration-Responsive Element Binding Protein 1c, 1e, and 1g Promote Stress Tolerance to Chilling, Heat, Drought, and Salt in Rice. Front. Plant Sci. 2022, 13, 851731. [Google Scholar] [CrossRef]
- Yongqi, H.; Jia, Z.; Defeng, F.; Zhibo, H.; Jiaming, L.; Yufei, Z.; Jinping, C.; Jifeng, Y.; Zhoufei, W. RNA-Seq study reveals AP2-Domain-Containing signalling regulators involved in initial imbibition of seed germination in rice. Rice Sci. 2020, 27, 302–314. [Google Scholar] [CrossRef]
- Mishra, M.; Kanwar, P.; Singh, A.; Pandey, A.; Kapoor, S.; Pandey, G.K. Plant Omics: Genome-Wide Analysis of ABA Repressor1 (ABR1) Related Genes in Rice During Abiotic Stress and Development. OMICS A J. Integr. Biol. 2013, 17, 439–450. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Long, Y.; Huang, J.; Xia, J. OsNAC45 is Involved in ABA Response and Salt Tolerance in Rice. Rice 2020, 13, 79. [Google Scholar] [CrossRef] [PubMed]
- Osnato, M.; Matias-Hernandez, L.; Aguilar-Jaramillo, A.E.; Kater, M.M.; Pelaz, S. Genes of the RAV Family Control Heading Date and Carpel Development in Rice. Plant Physiol. 2020, 183, 1663–1680. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.-B.; Li, J.; Qin, R.-Y.; Xu, R.-F.; Li, H.; Yang, Y.-C.; Ma, H.; Li, L.; Wei, P.-C.; Yang, J.-B. Identification of a Regulatory Element Responsible for Salt Induction of Rice Osrav2 through Ex Situ and In Situ Promoter Analysis. Plant Mol. Biol. 2016, 90, 49–62. [Google Scholar] [CrossRef]
- Moon, S.-J.; Park, H.J.; Kim, T.-H.; Kang, J.-W.; Lee, J.-Y.; Cho, J.-H.; Lee, J.-H.; Park, D.-S.; Byun, M.-O.; Kim, B.-G.; et al. Ostga2 Confers Disease Resistance to Rice against Leaf Blight by Regulating Expression Levels of Disease Related Genes Via Interaction with Nh1. PLoS ONE 2018, 13, e0206910. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, Y.; Wen, F.; Yao, D.; Wang, L.; Guo, J.; Ni, L.; Zhang, A.; Tan, M.; Jiang, M. A Novel Rice C2h2-Type Zinc Finger Protein, Zfp36, Is a Key Player Involved in Abscisic Acid-Induced Antioxidant Defence and Oxidative Stress Tolerance in Rice. J. Exp. Bot. 2014, 65, 5795–5809. [Google Scholar] [CrossRef]
- Fang, Y.; Xie, K.; Xiong, L. Conserved miR164-Targeted NAC Genes Negatively Regulate Drought Resistance in Rice. J. Exp. Bot. 2014, 65, 2119–2135. [Google Scholar] [CrossRef]
- Kusano, H.; Asano, T.; Shimada, H.; Kadowaki, K.-I. Molecular Characterization of Onac300, a Novel Nac Gene Specifically Expressed at Early Stages in Various Developing Tissues of Rice. Mol. Genet. Genom. 2005, 272, 616–626. [Google Scholar] [CrossRef]
- Sun, L.; Huang, L.; Hong, Y.; Zhang, H.; Song, F.; Li, D. Comprehensive Analysis Suggests Overlapping Expression of Rice Onac Transcription Factors in Abiotic and Biotic Stress Responses. Int. J. Mol. Sci. 2015, 16, 4306–4326. [Google Scholar] [CrossRef]
- Fang, Y.; You, J.; Xie, K.; Xie, W.; Xiong, L. Systematic Sequence Analysis and Identification of Tissue-Specific or Stress-Responsive Genes of Nac Transcription Factor Family in Rice. Mol. Genet. Genom. 2008, 280, 547–563. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, J.; Chen, S.; Ma, X.; Wei, H.; Chen, C.; Gao, N.; Zou, Y.; Kong, D.; Li, T.; et al. An APETALA2/ethylene responsive factor, Osebp89 Knockout Enhances Adaptation to Direct-Seeding on Wet Land and Tolerance to Drought Stress in Rice. Mol. Genet. Genom. 2020, 295, 941–956. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Zhang, Z.-L.; Zou, X.; Huang, J.; Ruas, P.; Thompson, D.; Shen, Q.J. Annotations and Functional Analyses of the Rice Wrky Gene Superfamily Reveal Positive and Negative Regulators of Abscisic Acid Signaling in Aleurone Cells. Plant Physiol. 2005, 137, 176–189. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Gu, L.; Ringler, P.; Smith, S.; Rushton, P.J.; Shen, Q.J. Three Wrky Transcription Factors Additively Repress Abscisic Acid and Gibberellin Signaling in Aleurone Cells. Plant Sci. 2015, 236, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.-L.; Shin, M.; Zou, X.; Huang, J.; Ho, T.-H.D.; Shen, Q.J. A Negative Regulator Encoded by a Rice Wrky Gene Represses Both Abscisic Acid and Gibberellins Signaling in Aleurone Cells. Plant Mol. Biol. 2009, 70, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Peng, Y.; Guo, Z. Constitutive Expression of Pathogen-Inducible Oswrky31 Enhances Disease Resistance and Affects Root Growth and Auxin Response in Transgenic Rice Plants. Cell Res. 2008, 18, 508–521. [Google Scholar] [CrossRef] [PubMed]
- Ye, M.; Glauser, G.; Lou, Y.; Erb, M.; Hu, L. Molecular Dissection of Early Defense Signaling Underlying Volatile-Mediated Defense Regulation and Herbivore Resistance in Rice. Plant Cell 2019, 31, 687–698. [Google Scholar] [CrossRef]
- Liu, X.; Bai, X.; Wang, X.; Chu, C. Oswrky71, a Rice Transcription Factor, Is Involved in Rice Defense Response. J. Plant Physiol. 2007, 164, 969–979. [Google Scholar] [CrossRef]
- Dai, X.; Wang, Y.; Zhang, W.-H. Oswrky74, a Wrky Transcription Factor, Modulates Tolerance to Phosphate Starvation in Rice. J. Exp. Bot. 2016, 67, 947–960. [Google Scholar] [CrossRef]
- Yang, A.; Dai, X.; Zhang, W.H. A R2r3-Type Myb Gene, Osmyb2, Is Involved in Salt, Cold, and Dehydration Tolerance in Rice. J. Exp. Bot. 2012, 63, 2541–2556. [Google Scholar] [CrossRef]
- El-Kereamy, A.; Bi, Y.-M.; Ranathunge, K.; Beatty, P.H.; Good, A.G.; Rothstein, S.J. The Rice R2r3-Myb Transcription Factor Osmyb55 Is Involved in the Tolerance to High Temperature and Modulates Amino Acid Metabolism. PLoS ONE 2012, 7, e52030. [Google Scholar] [CrossRef]
- Akhter, D.; Qin, R.; Nath, U.K.; Eshag, J.; Jin, X.; Shi, C. A Rice Gene, Ospl, Encoding a Myb Family Transcription Factor Confers Anthocyanin Synthesis, Heat Stress Response and Hormonal Signaling. Gene 2019, 699, 62–72. [Google Scholar] [CrossRef]
- Xiong, H.; Li, J.; Liu, P.; Duan, J.; Zhao, Y.; Guo, X.; Li, Y.; Zhang, H.; Ali, J.; Li, Z. Overexpression of Osmyb48-1, a Novel Myb-Related Transcription Factor, Enhances Drought and Salinity Tolerance in Rice. PLoS ONE 2014, 9, e92913. [Google Scholar] [CrossRef]
- Yun, K.-Y.; Park, M.R.; Mohanty, B.; Herath, V.; Xu, F.; Mauleon, R.; Wijaya, E.; Bajic, V.B.; Bruskiewich, R.; de Los Reyes, B.G. Transcriptional Regulatory Network Triggered by Oxidative Signals Configures the Early Response Mechanisms of Japonica Rice to Chilling Stress. BMC Plant Biol. 2010, 10, 16. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhang, Q.; Shou, H.-X. Identification and Expression Analysis of OsHsfs in rice. J. Zhejiang Univ. B 2009, 10, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Yi, F.; Gu, W.; Chen, J.; Song, N.; Gao, X.; Zhang, X.; Zhou, Y.; Ma, X.; Song, W.; Zhao, H.; et al. High Temporal-Resolution Transcriptome Landscape of Early Maize Seed Development. Plant Cell 2019, 31, 974–992. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, C.; Wang, C.; Yun, L.; Song, L.; Idrees, M.; Liu, H.; Zhang, Q.; Yang, J.; Zheng, X.; et al. Oshsfb4b Confers Enhanced Drought Tolerance in Transgenic Arabidopsis and Rice. Int. J. Mol. Sci. 2022, 23, 10830. [Google Scholar] [CrossRef]
- Wang, M.; Yang, D.; Ma, F.; Zhu, M.; Shi, Z.; Miao, X. Oshlh61-Osbhlh96 Influences Rice Defense to Brown Planthopper through Regulating the Pathogen-Related Genes. Rice 2019, 12, 9. [Google Scholar] [CrossRef] [PubMed]
- Wei, K.; Chen, H. Comparative Functional Genomics Analysis of Bhlh Gene Family in Rice, Maize and Wheat. BMC Plant Biol. 2018, 18, 309. [Google Scholar] [CrossRef]
- Li, X.; Duan, X.; Jiang, H.; Sun, Y.; Tang, Y.; Yuan, Z.; Guo, J.; Liang, W.; Chen, L.; Yin, J.; et al. Genome-Wide Analysis of Basic/Helix-Loop-Helix Transcription Factor Family in Rice and Arabidopsis. Plant Physiol. 2006, 141, 1167–1184. [Google Scholar] [CrossRef]
- Miyamoto, K.; Shimizu, T.; Mochizuki, S.; Nishizawa, Y.; Minami, E.; Nojiri, H.; Yamane, H.; Okada, K. Stress-Induced Expression of the Transcription Factor Rerj1 Is Tightly Regulated in Response to Jasmonic Acid Accumulation in Rice. Protoplasma 2013, 250, 241–249. [Google Scholar] [CrossRef]
- Valea, I.; Motegi, A.; Kawamura, N.; Kawamoto, K.; Miyao, A.; Ozawa, R.; Takabayashi, J.; Gomi, K.; Nemoto, K.; Nozawa, A.; et al. The Rice Wound-Inducible Transcription Factor Rerj1 Sharing Same Signal Transduction Pathway with Osmyc2 Is Necessary for Defense Response to Herbivory and Bacterial Blight. Plant Mol. Biol. 2022, 109, 651–666. [Google Scholar] [CrossRef]
- Hozain, M.; Abdelmageed, H.; Lee, J.; Kang, M.; Fokar, M.; Allen, R.D.; Holaday, A.S. Expression of Atsap5 in Cotton Up-Regulates Putative Stress-Responsive Genes and Improves the Tolerance to Rapidly Developing Water Deficit and Moderate Heat Stress. J. Plant Physiol. 2012, 169, 1261–1270. [Google Scholar] [CrossRef]
- Kothari, K.S.; Dansana, P.K.; Giri, J.; Tyagi, A.K. Rice Stress Associated Protein 1 (Ossap1) Interacts with Aminotransferase (Osamtr1) and Pathogenesis-Related 1a Protein (Osscp) and Regulates Abiotic Stress Responses. Front. Plant Sci. 2016, 7, 1057. [Google Scholar] [CrossRef]
- Kapoor, R.; Kumar, G.; Pawar, L.; Salvi, P.; Devanna, B.N.; Singh, K.; Sharma, T.R. Stress Responsive Oshyprp16 Promoter Driven Early Expression of Resistance Gene PI54 Potentiate the Resistance against Magnaporthe oryzae in Transgenic Rice. Plant Sci. 2022, 324, 111413. [Google Scholar] [CrossRef]
- Huang, J.; Zhao, X.; Weng, X.; Wang, L.; Xie, W. The Rice B-Box Zinc Finger Gene Family: Genomic Identification, Characterization, Expression Profiling and Diurnal Analysis. PLoS ONE 2012, 7, e48242. [Google Scholar] [CrossRef] [PubMed]
- Chowrasia, S.; Panda, A.K.; Rawal, H.C.; Kaur, H.; Mondal, T.K. Identification of Jumonjic Domain Containing Gene Family among the Oryza Species and Their Expression Analysis in Fl478, a Salt Tolerant Rice Genotype. Plant Physiol. Biochem. 2018, 130, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Zhou, D.-X. Rice Jmjc Domain-Containing Gene Jmj706 Encodes H3k9 Demethylase Required for Floral Organ Development. Proc. Natl. Acad. Sci. USA 2008, 105, 13679–13684. [Google Scholar] [CrossRef]
- Ye, H.; Du, H.; Tang, N.; Li, X.; Xiong, L. Identification and Expression Profiling Analysis of Tify Family Genes Involved in Stress and Phytohormone Responses in Rice. Plant Mol. Biol. 2009, 71, 291–305. [Google Scholar] [CrossRef]
- Seo, J.-S.; Joo, J.; Kim, M.-J.; Kim, Y.-K.; Nahm, B.H.; Song, S.I.; Cheong, J.-J.; Lee, J.S.; Kim, J.-K.; Choi, Y.D. Osbhlh148, a Basic Helix-Loop-Helix Protein, Interacts with Osjaz Proteins in a Jasmonate Signaling Pathway Leading to Drought Tolerance in Rice. Plant J. 2011, 65, 907–921. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Li, D.; Mao, D.; Liu, X.; Ji, C.; Li, X.; Zhao, X.; Cheng, Z.; Chen, C.; Zhu, L. Overexpression of Microrna319 Impacts Leaf Morphogenesis and Leads to Enhanced Cold Tolerance in Rice (Oryza sativa L.). Plant Cell Environ. 2013, 36, 2207–2218. [Google Scholar] [CrossRef]
- Mondragón-Palomino, M.; Trontin, C. High Time for a Roll Call: Gene Duplication and Phylogenetic Relationships of TCP-Like Genes in Monocots. Ann. Bot. 2011, 107, 1533–1544. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, K.; Kurata, N.; Ohyanagi, H.; Hake, S. Genome-Wide Study of Knox Regulatory Network Reveals Brassinosteroid Catabolic Genes Important for Shoot Meristem Function in Rice. Plant Cell 2014, 26, 3488–3500. [Google Scholar] [CrossRef] [PubMed]
- Jasinski, S.; Piazza, P.; Craft, J.; Hay, A.; Woolley, L.; Rieu, I.; Phillips, A.; Hedden, P.; Tsiantis, M. Knox Action in Arabidopsis Is Mediated by Coordinate Regulation of Cytokinin and Gibberellin Activities. Curr. Biol. 2005, 15, 1560–1565. [Google Scholar] [CrossRef]
- Yanai, O.; Shani, E.; Dolezal, K.; Tarkowski, P.; Sablowski, R.; Sandberg, G.; Samach, A.; Ori, N. Arabidopsis Knoxi Proteins Activate Cytokinin Biosynthesis. Curr. Biol. 2005, 15, 1566–1571. [Google Scholar] [CrossRef]
- Sato, Y.; Sentoku, N.; Nagato, Y.; Matsuoka, M. Isolation and Characterization of a Rice Homebox Gene, Osh15. Plant Mol. Biol. 1998, 38, 983–997. [Google Scholar] [CrossRef]
- Niu, M.; Wang, H.; Yin, W.; Meng, W.; Xiao, Y.; Liu, D.; Zhang, X.; Dong, N.; Liu, J.; Yang, Y.; et al. Rice Dwarf and Low-Tillering and the Homeodomain Protein Osh15 Interact to Regulate Internode Elongation via Orchestrating Brassinosteroid Signaling and Metabolism. Plant Cell 2022, 34, 3754–3772. [Google Scholar] [CrossRef]
- Huang, L.; Jia, J.; Zhao, X.; Zhang, M.; Huang, X.; Ji, E.; Ni, L.; Jiang, M. The Ascorbate Peroxidase Apx1 Is a Direct Target of a Zinc Finger Transcription Factor Zfp36 and a Late Embryogenesis Abundant Protein Oslea5 Interacts with Zfp36 to Co-Regulate Osapx1 in Seed Germination in Rice. Biochem. Biophys. Res. Commun. 2018, 495, 339–345. [Google Scholar] [CrossRef]
- Agalou, A.; Purwantomo, S.; Övernäs, E.; Johannesson, H.; Zhu, X.; Estiati, A.; De Kam, R.J.; Engström, P.; Slamet-Loedin, I.; Zhu, Z.; et al. A Genome-Wide Survey of Hd-Zip Genes in Rice and Analysis of Drought-Responsive Family Members. Plant Mol. Biol. 2008, 66, 87–103. [Google Scholar] [CrossRef]
- Wu, W.; Zheng, X.-M.; Chen, D.; Zhang, Y.; Ma, W.; Zhang, H.; Sun, L.; Yang, Z.; Zhao, C.; Zhan, X.; et al. Oscol16, Encoding a Constans-Like Protein, Represses Flowering by Up-Regulating Ghd7 Expression in Rice. Plant Sci. 2017, 260, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Shalmani, A.; Jing, X.Q.; Shi, Y.; Muhammad, I.; Zhou, M.R.; Wei, X.Y.; Chen, Q.Q.; Li, W.Q.; Liu, W.T.; Chen, K.M. Characterization of B-Box Gene Family and Their Expression Profiles under Hormonal, Abiotic and Metal Stresses in Poaceae Plants. BMC Genom. 2019, 20, 27. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Wang, S.; Zhang, B.; Shang-Guan, K.; Shi, Y.; Zhang, D.; Liu, X.; Wu, K.; Xu, Z.; Fu, X.; et al. A Gibberellin-Mediated Della-Nac Signaling Cascade Regulates Cellulose Synthesis in Rice. Plant Cell 2015, 27, 1681–1696. [Google Scholar] [CrossRef] [PubMed]
- Wani, S.H.; Anand, S.; Singh, B.; Bohra, A.; Joshi, R. Wrky Transcription Factors and Plant Defense Responses: Latest Discoveries and Future Prospects. Plant Cell Rep. 2021, 40, 1071–1085. [Google Scholar] [CrossRef] [PubMed]
- Abdullah-Zawawi, M.-R.; Ahmad-Nizammuddin, N.-F.; Govender, N.; Harun, S.; Mohd-Assaad, N.; Mohamed-Hussein, Z.-A. Comparative Genome-Wide Analysis of Wrky, Mads-Box and Myb Transcription Factor Families in Arabidopsis and Rice. Sci. Rep. 2021, 11, 19678. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Hu, L.; Zhang, S.; Zhang, M.; Jiang, W.; Wu, T.; Du, X. Rice Oswrky50 Mediates Aba-Dependent Seed Germination and Seedling Growth, and Aba-Independent Salt Stress Tolerance. Int. J. Mol. Sci. 2021, 22, 8625. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Mei, E.; He, M.; Bu, Q.; Tian, X. Functions of Oswrky24, Oswrky70 and Oswrky53 in Regulating Grain Size in Rice. Planta 2022, 255, 92. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Zhang, Z.L.; Zou, X.; Yang, G.; Komatsu, S.; Shen, Q.J. Interactions of Two Abscisic-Acid Induced Wrky Genes in Repressing Gibberellin Signaling in Aleurone Cells. Plant J. 2006, 46, 231–242. [Google Scholar] [CrossRef]
- Zhang, Z.-L.; Xie, Z.; Zou, X.L.; Casaretto, J.; Ho, T.-H.D.; Shen, Q.J. A Rice Wrky Gene Encodes a Transcriptional Repressor of the Gibberellin Signaling Pathway in Aleurone Cells. Plant Physiol. 2004, 134, 1500–1513. [Google Scholar] [CrossRef]
- He, Q.; Lu, H.; Guo, H.; Wang, Y.; Zhao, P.; Li, Y.; Wang, F.; Xu, J.; Mo, X.; Mao, C. Osbhlh6 Interacts with Osspx4 and Regulates the Phosphate Starvation Response in Rice. Plant J. 2021, 105, 649–667. [Google Scholar] [CrossRef]
- Li, Z.; Liang, Y.; Yuan, Y.; Wang, L.; Meng, X.; Xiong, G.; Zhou, J.; Cai, Y.; Han, N.; Hua, L.; et al. OsBRXL4 regulates shoot gravitropism and rice tiller angle through affecting LAZY1 nuclear localization. Mol. Plant. 2019, 12, 1143–1156. [Google Scholar] [CrossRef]
- Papatheodorou, I.; Moreno, P.; Manning, J.; Fuentes, A.M.-P.; George, N.; Fexova, S.; Fonseca, N.A.; Füllgrabe, A.; Green, M.; Huang, N.; et al. Expression Atlas Update: From Tissues to Single Cells. Nucleic Acids Res. 2020, 48, D77–D83. [Google Scholar] [CrossRef]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food Security: The Challenge of Feeding 9 Billion People. Science 2010, 327, 812–818. [Google Scholar] [CrossRef]
- Miro, B.; Ismail, A.M. Tolerance of Anaerobic Conditions Caused by Flooding during Germination and Early Growth in Rice (Oryza sativa L.). Front. Plant Sci. 2013, 4, 269. [Google Scholar] [CrossRef]
- Haque, A.; Rafii, M.Y.; Yusoff, M.M.; Ali, N.S.; Yusuff, O.; Arolu, F.; Anisuzzaman, M. Flooding Tolerance in Rice: Adaptive Mechanism and Marker-Assisted Selection Breeding Approaches. Mol. Biol. Rep. 2023, 50, 2795–2812. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Singh, Y.; Xalaxo, S.; Verulkar, S.; Yadav, N.; Singh, S.; Singh, N.; Prasad, K.; Kondayya, K.; Rao, P.R.; et al. From Qtl to Variety-Harnessing the Benefits of Qtls for Drought, Flood and Salt Tolerance in Mega Rice Varieties of India through a Multi-Institutional Network. Plant Sci. 2016, 242, 278–287. [Google Scholar] [CrossRef]
- Fukao, T.; Xu, K.; Ronald, P.C.; Bailey-Serres, J. A Variable Cluster of Ethylene Response Factor–Like Genes Regulates Metabolic and Developmental Acclimation Responses to Submergence in Rice. Plant Cell 2006, 18, 2021–2034. [Google Scholar] [CrossRef]
- Fukao, T.; Bailey-Serres, J. Submergence Tolerance Conferred by Sub1A is Mediated by Slr1 and Slrl1 Restriction of Gibberellin Responses in Rice. Proc. Natl. Acad. Sci. USA 2008, 105, 16814–16819. [Google Scholar] [CrossRef] [PubMed]
- Locke, A.M.; Barding, G.A., Jr.; Sathnur, S.; Larive, C.K.; Bailey-Serres, J. Rice Sub1a Constrains Remodelling of the Transcriptome and Metabolome during Submergence to Facilitate Post-Submergence Recovery. Plant Cell Environ. 2018, 41, 721–736. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Biswas, S.; Kim, B.; Bailey-Serres, J.; Septiningsih, E.M. Improved Transformation and Regeneration of Indica Rice: Disruption of Sub1a as a Test Case Via Crispr-Cas9. Int. J. Mol. Sci. 2021, 22, 6989. [Google Scholar] [CrossRef]
- Hsu, S.-K.; Tung, C.-W. Genetic Mapping of Anaerobic Germination-Associated QTLs Controlling Coleoptile Elongation in Rice. Rice 2015, 8, 38. [Google Scholar] [CrossRef]
- Nghi, K.N.; Tondelli, A.; Valè, G.; Tagliani, A.; Marè, C.; Perata, P.; Pucciariello, C. Dissection of Coleoptile Elongation in Japonica Rice under Submergence through Integrated Genome-Wide Association Mapping and Transcriptional Analyses. Plant Cell Environ. 2019, 42, 1832–1846. [Google Scholar] [CrossRef]
- Harper, L.; Campbell, J.; Cannon, E.K.S.; Jung, S.; Poelchau, M.; Walls, R.; Andorf, C.; Arnaud, E.; Berardini, T.Z.; Birkett, C.; et al. Agbiodata Consortium Recommendations for Sustainable Genomics and Genetics Databases for Agriculture. Database 2018, 2018, bay088. [Google Scholar] [CrossRef]
- Naithani, S.; Gupta, P.; Preece, J.; Garg, P.; Fraser, V.; Padgitt-Cobb, L.K.; Martin, M.; Vining, K.; Jaiswal, P. Involving Community in Genes and Pathway Curation. Database 2019, 2019, bay146. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naithani, S.; Mohanty, B.; Elser, J.; D’Eustachio, P.; Jaiswal, P. Biocuration of a Transcription Factors Network Involved in Submergence Tolerance during Seed Germination and Coleoptile Elongation in Rice (Oryza sativa). Plants 2023, 12, 2146. https://doi.org/10.3390/plants12112146
Naithani S, Mohanty B, Elser J, D’Eustachio P, Jaiswal P. Biocuration of a Transcription Factors Network Involved in Submergence Tolerance during Seed Germination and Coleoptile Elongation in Rice (Oryza sativa). Plants. 2023; 12(11):2146. https://doi.org/10.3390/plants12112146
Chicago/Turabian StyleNaithani, Sushma, Bijayalaxmi Mohanty, Justin Elser, Peter D’Eustachio, and Pankaj Jaiswal. 2023. "Biocuration of a Transcription Factors Network Involved in Submergence Tolerance during Seed Germination and Coleoptile Elongation in Rice (Oryza sativa)" Plants 12, no. 11: 2146. https://doi.org/10.3390/plants12112146
APA StyleNaithani, S., Mohanty, B., Elser, J., D’Eustachio, P., & Jaiswal, P. (2023). Biocuration of a Transcription Factors Network Involved in Submergence Tolerance during Seed Germination and Coleoptile Elongation in Rice (Oryza sativa). Plants, 12(11), 2146. https://doi.org/10.3390/plants12112146