


Establishing a Virus-Induced Gene Silencing System in Lycoris chinensis

 and
and
Abstract
:1. Introduction
2. Results
2.1. Comparison of Agrobacterium Infection Efficiency of Different Infiltration Methods
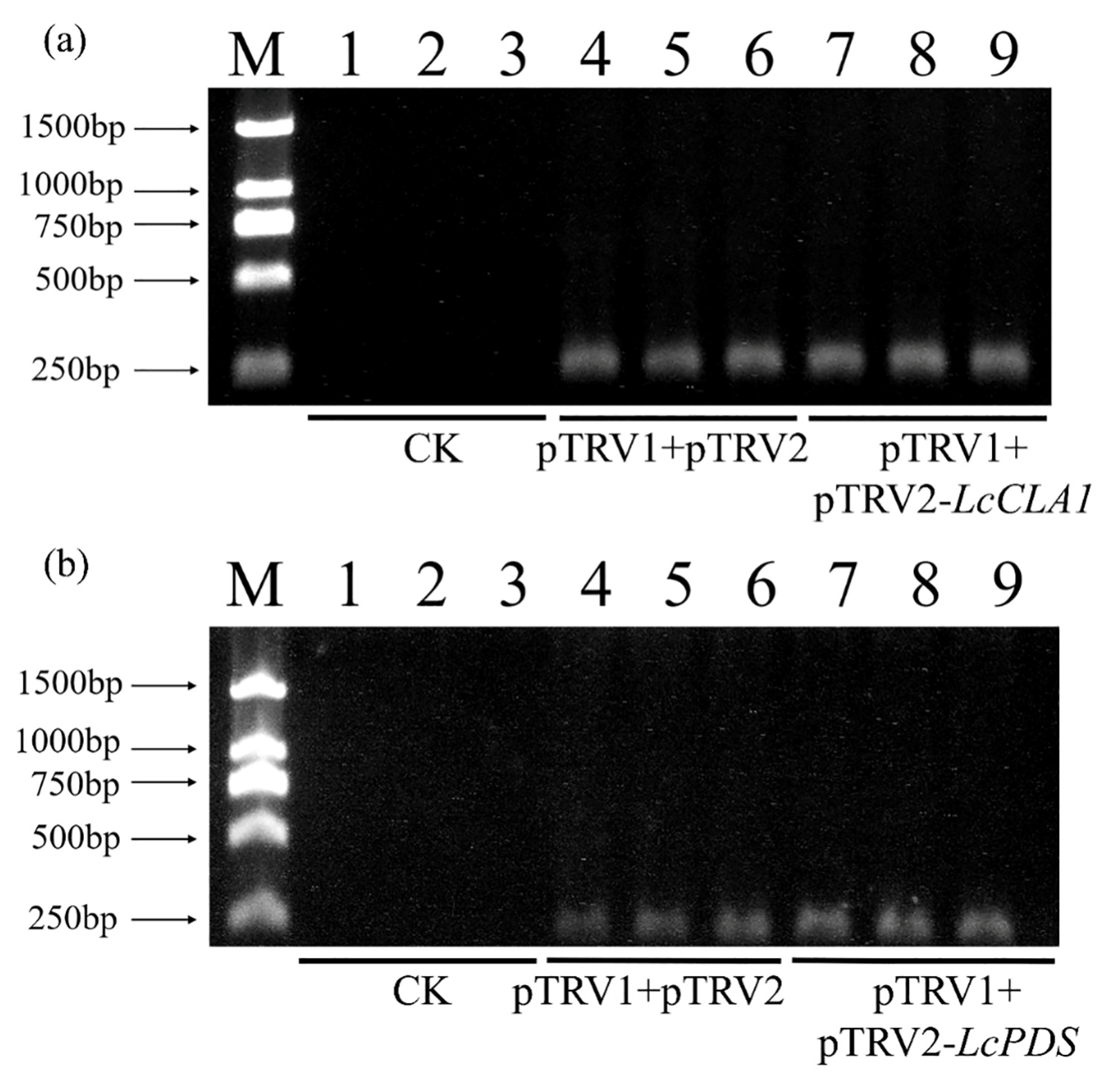
2.2. Silencing Efficiency of LcCLA1 and LcPDS in L. chinensis after Infection
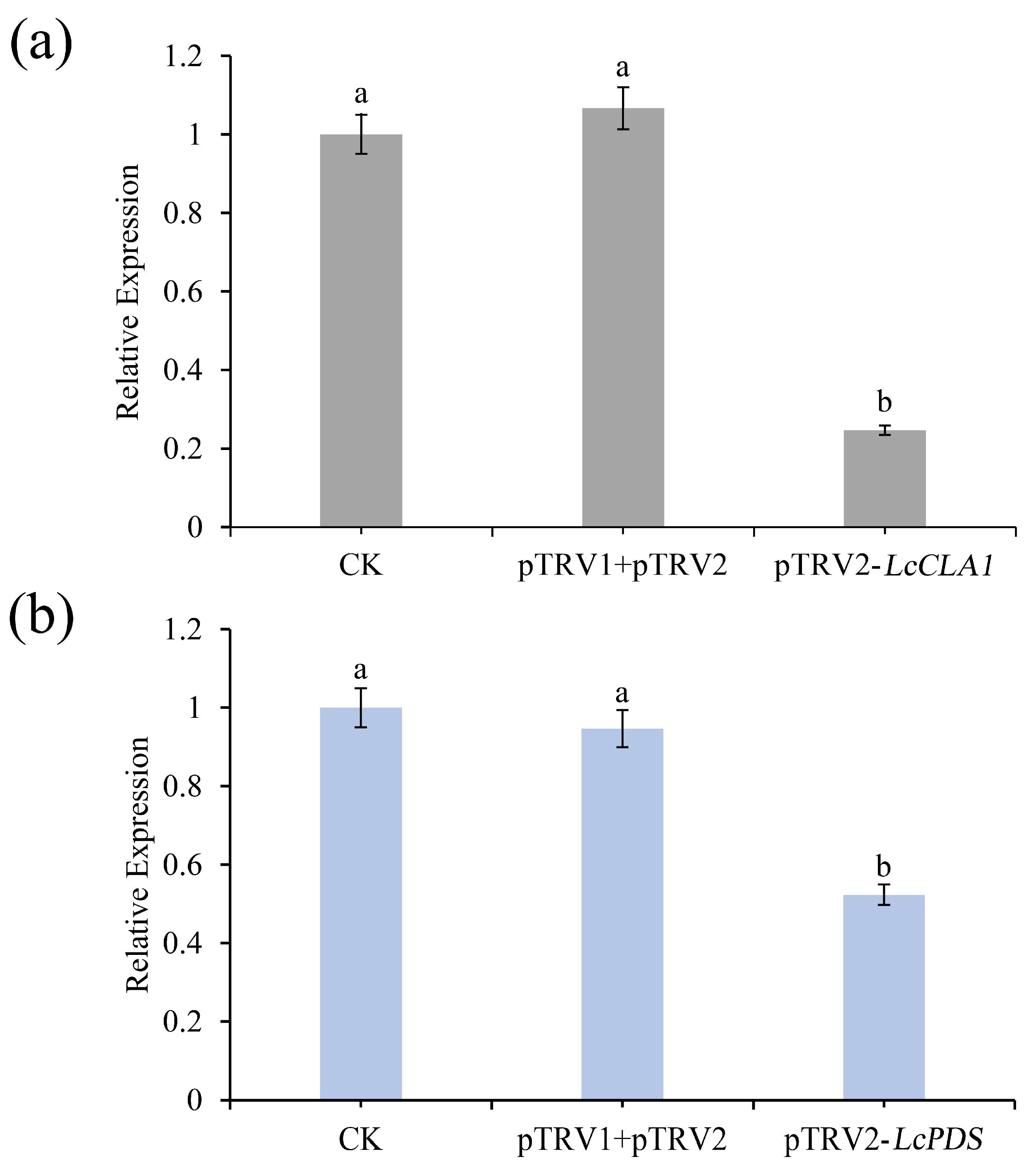
2.3. qRT-PCR Analysis of LcCLA1 and LcPDS Silencing Level
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Infection Methods
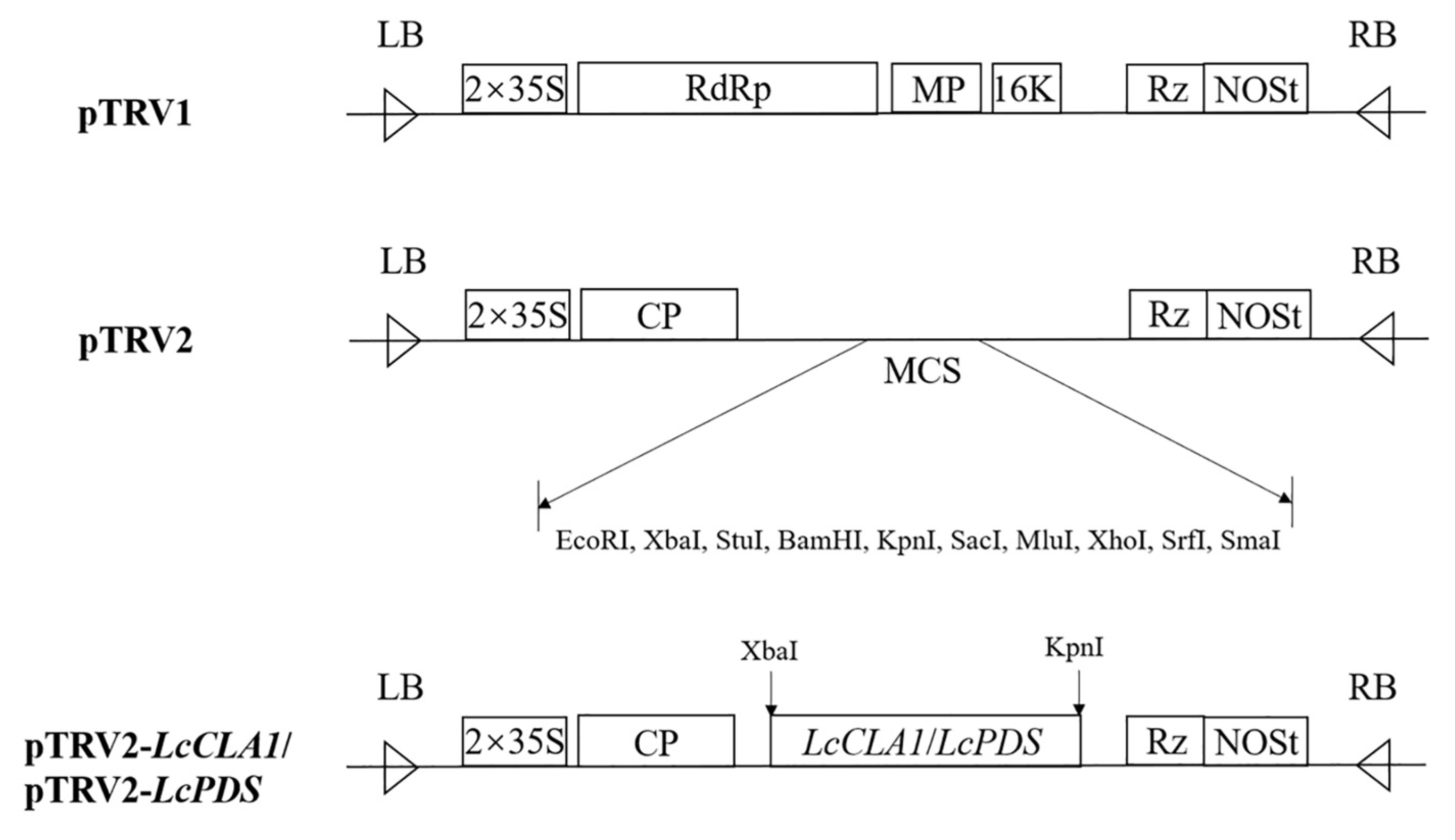
4.2. Cloning of the LcCLA1 and LcPDS Genes and VIGS Vector Construction
4.3. Preparation of Agroinfiltration Infection Solution
4.4. Expression Analysis of LcCLA1 and LcPDS Using qRT-PCR
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kurita, S. Variation and evolution on the karyotype of Lycoris, Amaryllidaceae II. Karyotype analysis of ten taxa among which seven are native in China. Cytologia 1987, 52, 19–40. [Google Scholar] [CrossRef] [Green Version]
- Tae, K.H.; Ko, S.C. A taxonomic study on the genus Lycoris (Amaryllidaceae). Kor. J. Plant Tax. 1996, 26, 19–35. [Google Scholar] [CrossRef]
- Cahlikova, L.; Breiterova, K.; Opletal, L. Chemistry and Biological Activity of Alkaloids from the Genus Lycoris (Amaryllidaceae). Molecules 2020, 25, 4797. [Google Scholar] [CrossRef]
- Yeo, H.J.; Kim, Y.J.; Nguyen, B.V.; Park, Y.E.; Park, C.H.; Kim, H.H.; Kim, J.K.; Park, S.U. Comparison of Secondary Metabolite Contents and Metabolic Profiles of Six Lycoris Species. Horticulturae 2021, 7, 5. [Google Scholar] [CrossRef]
- Sun, B.; Wang, P.; Wang, R.; Li, Y.; Xu, S. Molecular Cloning and Characterization of a meta/para-O-Methyltransferase from Lycoris aurea. Int. J. Mol. Sci. 2018, 19, 1911. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Cheng, G.; Shu, X.; Wang, N.; Wang, Z. Transcriptome Analysis of Lycoris chinensis Bulbs Reveals Flowering in the Age-Mediated Pathway. Biomolecules 2022, 12, 899. [Google Scholar] [CrossRef]
- Wang, N.; Song, G.; Zhang, F.; Shu, X.; Cheng, G.; Zhuang, W.; Wang, T.; Li, Y.; Wang, Z. Characterization of the WRKY Gene Family Related to Anthocyanin Biosynthesis and the Regulation Mechanism under Drought Stress and Methyl Jasmonate Treatment in Lycoris radiata. Int. J. Mol. Sci. 2023, 24, 2423. [Google Scholar] [CrossRef]
- Cheng, G.; Zhang, F.; Shu, X.; Wang, N.; Wang, T.; Zhuang, W.; Wang, Z. Identification of Differentially Expressed Genes Related to Floral Bud Differentiation and Flowering Time in Three Populations of Lycoris radiata. Int. J. Mol. Sci. 2022, 23, 14036. [Google Scholar] [CrossRef]
- Cai, J.; Fan, J.; Wei, X.; Zhang, D.; Ren, J.; Zhang, L. Differences in floral development between Lycoris radiata and Lycoris sprengeri. Scienceasia 2020, 46, 271. [Google Scholar] [CrossRef]
- Senthil-Kumar, M.; Mysore, K.S. New dimensions for VIGS in plant functional genomics. Trends Plant Sci. 2011, 16, 656–665. [Google Scholar] [CrossRef]
- Liu, Y.; Schiff, M.; Dinesh-Kumar, S.P. Virus-induced gene silencing in tomato. Plant J. 2002, 31, 777–786. [Google Scholar] [CrossRef] [PubMed]
- Burch-Smith, T.M.; Anderson, J.C.; Martin, G.B.; Dinesh-Kumar, S.P. Applications and advantages of virus-induced gene silencing for gene function studies in plants. Plant J. 2004, 39, 734–746. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, P.M.; Wang, M.B.; Lough, T. Gene silencing as an adaptive defence against viruses. Nature 2001, 411, 834–842. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Ye, S.; Wang, L.; Duan, Y.; Lu, W.; Liu, H.; Fan, D.; Zhang, F.; Luo, K. Heterologous gene silencing induced by tobacco rattle virus (TRV) is efficient for pursuing functional genomics studies in woody plants. Plant Cell Tiss. Org. 2014, 116, 163–174. [Google Scholar] [CrossRef]
- Burch-Smith, T.M.; Schiff, M.; Liu, Y.; Dinesh-Kumar, S.P. Efficient virus-induced gene silencing in Arabidopsis. Plant Physiol. 2006, 142, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Chung, E.; Seong, E.; Kim, Y.C.; Chung, E.J.; Oh, S.K.; Lee, S.; Park, J.M.; Joung, Y.H.; Choi, D. A Method of High Frequency Virus-induced Gene Silencing in Chili Pepper (Capsicum annuum L. cv. Bukang). Mol. Cells 2004, 17, 377–380. [Google Scholar]
- Xu, H.; Xu, L.; Yang, P.; Cao, Y.; Tang, Y.; He, G.; Yuan, S.; Lei, J.; Ming, J. Virus-induced Phytoene Desaturase (PDS) Gene Silencing Using Tobacco Rattle Virus in Lilium × formolongi. Hortic. Plant J. 2019, 5, 31–38. [Google Scholar] [CrossRef]
- Kumagai, M.H.; Donson, J.; Della-Cioppa, G.; Harvey, D.; Hanley, K.; Grill, L. Cytoplasmic inhibition of carotenoid biosynthesis with virus-derived RNA. Proc. Natl. Acad. Sci. USA 1995, 92, 1679–1683. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, M.T.; Voinnet, O.; Baulcombe, D.C. Initiation and Maintenance of Virus-Induced Gene Silencing. Plant Cell 1998, 10, 937–946. [Google Scholar] [CrossRef]
- Peele, C.; Jordan, C.V.; Muangsan, N.; Turnage, M.; Egelkrout, E.; Eagle, P.; Robertson, D. Silencing of a meristematic gene using geminivirus-derived vectors. Plant J. 2001, 27, 357–366. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Schiff, M.; Marathe, R.; Dinesh-Kumar, S.P. Tobacco Rar1, EDS1 and NPR1/NIM1 like genes are required for N-mediated resistance to tobacco mosaic virus. Plant J. 2002, 30, 415–429. [Google Scholar] [CrossRef]
- Igarashi, A.; Yamagata, K.; Sugai, T.; Takahashi, Y.; Sugawara, E.; Tamura, A.; Yaegashi, H.; Yamagishi, N.; Takahashi, T.; Isogai, M.; et al. Apple latent spherical virus vectors for reliable and effective virus-induced gene silencing among a broad range of plants including tobacco, tomato, Arabidopsis thaliana, cucurbits, and legumes. Virology 2009, 386, 407–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, X.; Zhang, Y.; Zhang, L.; Wang, B.; Zhao, Y.; Irfan, M.; Chen, L.; Feng, Y. Regulation of MYB Transcription Factors of Anthocyanin Synthesis in Lily Flowers. Front. Plant Sci. 2021, 12, 761668. [Google Scholar] [CrossRef]
- Zhou, P.; Peng, J.; Zeng, M.; Wu, L.; Fan, Y.; Zeng, L. Virus-induced gene silencing (VIGS) in Chinese narcissus and its use in functional analysis of NtMYB3. Hortic. Plant J. 2021, 7, 565–572. [Google Scholar] [CrossRef]
- Zhao, Y.; Yang, Z.; Ding, Y.; Liu, L.; Han, X.; Zhan, J.; Wei, X.; Diao, Y.; Qin, W.; Wang, P.; et al. Over-expression of an R2R3 MYB Gene, GhMYB73, increases tolerance to salt stress in transgenic Arabidopsis. Plant Sci. 2019, 286, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Zhu, L.; Zhang, X.; Guan, Q.; Xiao, S.; Min, L.; Zhang, X. GhCPK33 Negatively Regulates Defense against Verticillium dahliae by Phosphorylating GhOPR3. Plant Physiol. 2018, 178, 876–889. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Zhang, Y.; Wang, X.; Wang, W.; Li, Z.; Wu, J.; Wang, G.; Wu, L.; Zhang, G.; Ma, Z. HyPRP1 performs a role in negatively regulating cotton resistance to V. dahliae via the thickening of cell walls and ROS accumulation. BMC Plant Biol. 2018, 18, 339. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Feng, J.; Ma, W.; Zhou, Y.; Ma, Z. GhCLCg-1, a Vacuolar Chloride Channel, Contributes to Salt Tolerance by Regulating Ion Accumulation in Upland Cotton. Front. Plant Sci. 2021, 12, 765173. [Google Scholar] [CrossRef] [PubMed]
- Manhães, A.M.E.D.A.; de Oliveira, M.V.V.; Shan, L. Establishment of an Efficient Virus-Induced Gene Silencing (VIGS) Assay in Arabidopsis by Agrobacterium-Mediated Rubbing Infection. In Plant Gene Silencing: Methods and Protocols; Mysore, K.S., Senthil-Kumar, M., Eds.; Springer: New York, NY, USA, 2015; pp. 235–241. [Google Scholar]
- Kirigia, D.; Runo, S.; Alakonya, A. A virus-induced gene silencing (VIGS) system for functional genomics in the parasitic plant Striga hermonthica. Plant Methods 2014, 10, 16. [Google Scholar] [CrossRef] [Green Version]
- Xie, L.; Zhang, Q.; Sun, D.; Yang, W.; Hu, J.; Niu, L.; Zhang, Y. Virus-induced gene silencing in the perennial woody Paeonia ostii. PeerJ 2019, 7, e7001. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Xu, L.; Yang, P.; Cao, Y.; Tang, Y.; He, G.; Yuan, S.; Ming, J. Tobacco rattle virus-induced PHYTOENE DESATURASE (PDS) and Mg-chelatase H subunit (ChlH) gene silencing in Solanum pseudocapsicum L. PeerJ 2018, 6, e7001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sung, Y.C.; Lin, C.P.; Chen, J.C. Optimization of virus-induced gene silencing in Catharanthus roseus. Plant Pathol. 2014, 63, 1159–1167. [Google Scholar] [CrossRef]
- Nishii, K.; Fei, Y.; Hudson, A.; Moller, M.; Molnar, A. Virus-induced Gene Silencing in Streptocarpus rexii (Gesneriaceae). Mol. Biotechnol. 2020, 62, 317–325. [Google Scholar] [CrossRef]
- Zhou, J.; Hunter, D.A.; Lewis, D.H.; McManus, M.T.; Zhang, H. Insights into carotenoid accumulation using VIGS to block different steps of carotenoid biosynthesis in petals of California poppy. Plant Cell Rep. 2018, 37, 1311–1323. [Google Scholar] [CrossRef]
- Vaia, G.; Pavese, V.; Moglia, A.; Cristofori, V.; Silvestri, C. Knockout of phytoene desaturase gene using CRISPR/Cas9 in highbush blueberry. Front. Plant Sci. 2022, 13, 1074541. [Google Scholar] [CrossRef] [PubMed]
- Lindbo, J.A. High-efficiency protein expression in plants from agroinfection-compatible Tobacco mosaic virus expression vectors. BMC Biotechnol. 2007, 7, 52. [Google Scholar] [CrossRef] [Green Version]
- Pang, J.; Zhu, Y.; Li, Q.; Liu, J.; Tian, Y.; Liu, Y.; Wu, J. Development of Agrobacterium-mediated virus-induced gene silencing and performance evaluation of four marker genes in Gossypium barbadense. PLoS ONE 2013, 8, e73211. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Fu, D.; Zhu, B.; Yan, H.; Shen, X.; Zuo, J.; Zhu, Y.; Luo, Y. Virus-induced Gene Silencing in Eggplant (Solanum melongena). J. Integr. Plant Biol. 2012, 54, 422–429. [Google Scholar] [CrossRef]
- Liu, J.; Huang, Y.; Kong, L.; Yu, X.; Feng, B.; Liu, D.; Zhao, B.; Mendes, G.C.; Yuan, P.; Ge, D.; et al. The malectin-like receptor-like kinase LETUM1 modulates NLR protein SUMM2 activation via MEKK2 scaffolding. Nat. Plants 2020, 6, 1106–1115. [Google Scholar] [CrossRef]
- Cao, Z.; Zhao, T.; Wang, L.; Han, J.; Chen, J.; Hao, Y.; Guan, X. The lincRNA XH123 is involved in cotton cold-stress regulation. Plant Mol. Biol. 2021, 106, 521–531. [Google Scholar] [CrossRef]
- Wan, K.; Lu, K.; Gao, M.; Zhao, T.; He, Y.; Yang, D.L.; Tao, X.; Xiong, G.; Guan, X. Functional analysis of the cotton CLE polypeptide signaling gene family in plant growth and development. Sci. Rep. 2021, 11, 5060. [Google Scholar] [CrossRef]
- Cai, Y.; Cai, X.; Wang, Q.; Wang, P.; Zhang, Y.; Cai, C.; Xu, Y.; Wang, K.; Zhou, Z.; Wang, C.; et al. Genome sequencing of the Australian wild diploid species Gossypium australe highlights disease resistance and delayed gland morphogenesis. Plant Biotechnol. J. 2020, 18, 814–828. [Google Scholar] [CrossRef] [Green Version]
- Liu, E.; Page, J.E. Optimized cDNA libraries for virus-induced gene silencing (VIGS) using tobacco rattle virus. Plant Methods 2008, 4, 5–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bredow, M.; Natukunda, M.I.; Beernink, B.M.; Chicowski, A.S.; Salas-Fernandez, M.G.; Whitham, S.A. Characterization of a foxtail mosaic virus vector for gene silencing and analysis of innate immune responses in Sorghum bicolor. Mol. Plant Pathol. 2023, 24, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Na, R.; Nowak, J.S.; Qiu, Y.; Lu, Q.S.; Yang, C.; Marsolais, F.; Tian, L. Development of a Csy4-processed guide RNA delivery system with soybean-infecting virus ALSV for genome editing. BMC Plant Biol. 2021, 21, 419. [Google Scholar] [CrossRef]
- Yan, H.X.; Fu, D.Q.; Zhu, B.Z.; Liu, H.P.; Shen, X.Y.; Luo, Y.B. Sprout vacuum-infiltration: A simple and efficient agroinoculation method for virus-induced gene silencing in diverse solanaceous species. Plant Cell Rep. 2012, 31, 1713–1722. [Google Scholar] [CrossRef]
- Deng, X.; Elomaa, P.; Nguyen, C.X.; Hytonen, T.; Valkonen, J.P.; Teeri, T.H. Virus-induced gene silencing for Asteraceae--a reverse genetics approach for functional genomics in Gerbera hybrida. Plant Biotechnol. J. 2012, 10, 970–978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinesh-Kumar, S.; Anandalakshmi, R.; Marathe, R.; Schiff, M.; Liu, Y. Virus-Induced Gene Silencing; Humana Press: Totowa, NJ, USA, 2003; pp. 287–293. [Google Scholar]
- Zhang, J.; Wang, F.; Zhang, C.; Zhang, J.; Chen, Y.; Liu, G.; Zhao, Y.; Hao, F.; Zhang, J. A novel VIGS method by agroinoculation of cotton seeds and application for elucidating functions of GhBI-1 in salt-stress response. Plant Cell Rep. 2018, 37, 1091–1100. [Google Scholar] [CrossRef]
- Zhang, J.; Yu, D.; Zhang, Y.; Liu, K.; Xu, K.; Zhang, F.; Wang, J.; Tan, G.; Nie, X.; Ji, Q.; et al. Vacuum and Co-cultivation Agroinfiltration of (Germinated) Seeds Results in Tobacco Rattle Virus (TRV) Mediated Whole-Plant Virus-Induced Gene Silencing (VIGS) in Wheat and Maize. Front. Plant Sci. 2017, 8, 393. [Google Scholar] [CrossRef] [Green Version]
- Ratcliff, F.; Martin-Hernandez, A.M.; Baulcombe, D.C. Technical advance: Tobacco rattle virus as a vector for analysis of gene function by silencing. Plant J. 2001, 25, 237–245. [Google Scholar] [CrossRef]
- Höfgen, R.; Willmitzer, L. Storage of competent cells for Agrobacterium transformation. Nucleic Acids Res. 1988, 16, 9877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chong, X.; Wang, Y.; Xu, X.; Zhang, F.; Wang, C.; Zhou, Y.; Zhou, T.; Li, Y.; Lu, X.; Chen, H. Efficient Virus-Induced Gene Silencing in Ilex dabieshanensis Using Tobacco Rattle Virus. Forests 2023, 14, 488. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence |
|---|---|
| EXP1-RT-F | TTGATGTTGACAAGGTAAGGTGC |
| EXP1-RT-R | AGGCAGGAAATCTCCAAAGC |
| LcCLA1-F | TTGACGGGCAGGAGGGAC |
| LcCLA1-R | GCGGAGGAAGCAGTTTAGGC |
| LcCLA1-XbaI-F | CTAGTCTAGATTGACGGGCAGGAGGGAC |
| LcCLA1-KpnI-R | CGGGGTACCGCGGAGGAAGCAGTTTAGGC |
| LcCLA1-RT-F | GTTGCTCATTTCTTGGCACTCA |
| LcCLA1-RT-R | CAGCACCAACGGTCTCCACT |
| LcPDS-F | GTTAGGTCAGTTTCTGCTGTTTGTC |
| LcPDS-R | GTTGTTCCTCAAGATAGCCCATA |
| LcPDS-XbaI-F | CTAGTCTAGAGTTAGGTCAGTTTCTGCTGTTTGTC |
| LcPDS-KpnI-R | CGGGGTACCGTTGTTCCTCAAGATAGCCCATA |
| LcPDS-RT-F | AAAACCGTACCCGACTGTGAG |
| LcPDS-RT-R | CGGCTGTAGACACTTTCTTGCT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, G.; Shu, X.; Wang, Z.; Wang, N.; Zhang, F. Establishing a Virus-Induced Gene Silencing System in Lycoris chinensis. Plants 2023, 12, 2458. https://doi.org/10.3390/plants12132458
Cheng G, Shu X, Wang Z, Wang N, Zhang F. Establishing a Virus-Induced Gene Silencing System in Lycoris chinensis. Plants. 2023; 12(13):2458. https://doi.org/10.3390/plants12132458
Chicago/Turabian StyleCheng, Guanghao, Xiaochun Shu, Zhong Wang, Ning Wang, and Fengjiao Zhang. 2023. "Establishing a Virus-Induced Gene Silencing System in Lycoris chinensis" Plants 12, no. 13: 2458. https://doi.org/10.3390/plants12132458
APA StyleCheng, G., Shu, X., Wang, Z., Wang, N., & Zhang, F. (2023). Establishing a Virus-Induced Gene Silencing System in Lycoris chinensis. Plants, 12(13), 2458. https://doi.org/10.3390/plants12132458