


Selection of Short-Day Strawberry Genotypes through Multivariate Analysis
 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Genotypes Selection
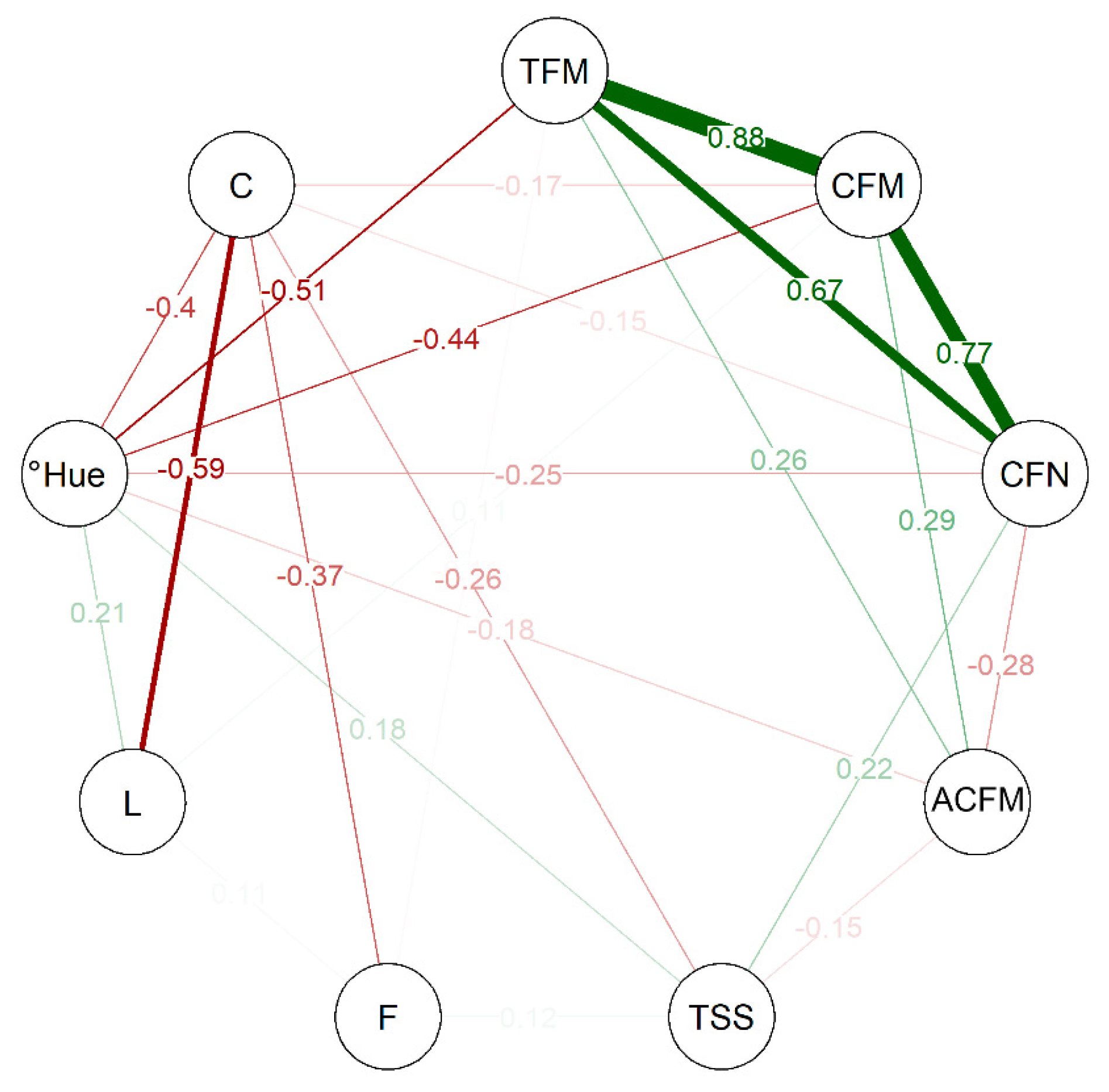
2.2. Multivariate Analyses among Selected Genotypes
3. Discussion
4. Materials and Methods
4.1. Experimental Hybrids Obtaining
4.2. Experimental Field
4.3. Production and Post-Harvest Traits Assessment
4.4. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Giampieri, F.; Alvarez-Suarez, J.M.; Battino, M. Strawberry and Human Health: Effects beyond Antioxidant Activity. J. Agric. Food Chem. 2014, 62, 3867–3876. [Google Scholar] [CrossRef] [PubMed]
- Tazzo, I.F.; Fagherazi, A.F.; Lerin, S.; Kretzschmar, A.A.; Rufato, L. Exigência Térmica de Duas Seleções e Quatro Cultivares de Morangueiro Cultivado No Planalto Catarinense. Rev. Bras. Frutic. 2015, 37, 550–558. [Google Scholar] [CrossRef] [Green Version]
- Lieten, P. Advances in Strawberry Substrate Culture during the Last Twenty Years in the Netherlands and Belgium. Int. J. Fruit Sci. 2013, 13, 84–90. [Google Scholar] [CrossRef]
- Neri, D.; Baruzzi, G.; Massetani, F.; Faedi, W. Strawberry Production in Forced and Protected Culture in Europe as a Response to Climate Change. Can. J. Plant Sci. 2012, 92, 1021–1036. [Google Scholar] [CrossRef] [Green Version]
- Flanagan, R.D.; Samtani, J.B.; Manchester, M.A.; Romelczyk, S.; Johnson, C.S.; Lawrence, W.; Pattison, J. On-Farm Evaluation of Strawberry Cultivars in Coastal Virginia. HortTechnology 2020, 30, 789–796. [Google Scholar] [CrossRef]
- Zeist, A.R.; de Resende, J.T.; Lima, R.B.; Gabriel, A.; Henschel, J.M.; da Silva, I.F. Phenology and Agronomic Components of First and Second-Cycle Strawberry. Hortic. Bras. 2019, 37, 29–34. [Google Scholar] [CrossRef]
- Barth, E.; de Resende, J.T.V.; Moreira, A.F.P.; Mariguele, K.H.; Zeist, A.R.; Silva, M.B.; Stulzer, G.C.G.; Mafra, J.G.M.; Simões Azeredo Gonçalves, L.; Roberto, S.R. Selection of Experimental Hybrids of Strawberry Using Multivariate Analysis. Agronomy 2020, 10, 598. [Google Scholar] [CrossRef] [Green Version]
- Barth, E.; de Resende, J.T.V.; Mariguele, K.H.; de Resende, M.D.V.; da Silva, A.L.B.R.; Ru, S. Multivariate Analysis Methods Improve the Selection of Strawberry Genotypes with Low Cold Requirement. Sci. Rep. 2022, 12, 11458. [Google Scholar] [CrossRef]
- Corrêa, J.V.W.; Weber, G.G.; Zeist, A.R.; Resende, J.T.V.; Silva, P.R. ISSR Analysis Reveals High Genetic Variation in Strawberry Three-Way Hybrids Developed for Tropical Regions. Plant Mol. Biol. Rep. 2021, 39, 566–576. [Google Scholar] [CrossRef]
- Moreira, A.F.P.; de Resende, J.T.V.; Shimizu, G.D.; Hata, F.T.; do Nascimento, D.; Oliveira, L.V.B.; Zanin, D.S.; Mariguele, K.H. Characterization of Strawberry Genotypes with Low Chilling Requirement for Cultivation in Tropical Regions. Sci. Hortic. 2022, 292, 110629. [Google Scholar] [CrossRef]
- Barth, E.; Resende, J.; Zeist, A.; Mariguele, K.; Zeist, R.; Gabriel, A.; Camargo, C.; Piran, F. Yield and Quality of Strawberry Hybrids under Subtropical Conditions. Genet. Mol. Res. 2019, 18, GMR18156. [Google Scholar] [CrossRef]
- Galvão, A.G.; Resende, L.V.; Maluf, W.R.; de Resende, J.T.V.; Ferraz, A.K.L.; Marodin, J.C. Breeding New Improved Clones for Strawberry Production in Brazil. Acta Sci. Agron. 2017, 39, 149–155. [Google Scholar] [CrossRef] [Green Version]
- Vergauwen, D.; Smet, I.D. The Strawberry Tales: Size Matters. Trends Plant Sci. 2019, 24, 1–3. [Google Scholar] [CrossRef]
- Mulamba, N.; Mock, J. Improvement of Yield Potential of the ETo Blanco Maize (Zea mays L.) Population by Breeding for Plant Traits. Egypt. J. Genet. Cytol. 1978, 7, 40–51. [Google Scholar]
- Chiomento, J.L.T.; Júnior, E.P.L.; D’Agostini, M.; Nardi, F.S.; Trentin, T.d.S.; Dornelles, A.G.; Huzar-Novakowiski, J.; Calvete, E.O. Horticultural Potential of Nine Strawberry Cultivars by Greenhouse Production in Brazil: A View through Multivariate Analysis. Sci. Hortic. 2021, 279, 109738. [Google Scholar] [CrossRef]
- Cassani, L.; Santos, M.; Gerbino, E.; Moreira, M.d.R.; Gómez-Zavaglia, A. A Combined Approach of Infrared Spectroscopy and Multivariate Analysis for the Simultaneous Determination of Sugars and Fructans in Strawberry Juices during Storage. J. Food Sci. 2018, 83, 631–638. [Google Scholar] [CrossRef]
- Khan, M.N.; Sarwar, A.; Bhutto, S.; Wahab, M.F. Physicochemical Characterization of the Strawberry Samples on Regional Basis Using Multivariate Analysis. Int. J. Food Prop. 2010, 13, 789–799. [Google Scholar] [CrossRef] [Green Version]
- Hervé, M.R.; Nicolè, F.; Cao, K.-A.L. Multivariate Analysis of Multiple Datasets: A Practical Guide for Chemical Ecology. J. Chem. Ecol. 2018, 44, 215–234. [Google Scholar] [CrossRef]
- Lapshin, V.; Yakovenko, V.; Shcheglov, S. Evaluation of the Breeding and Genetic Potential of Strawberry Varieties and Hybrids Based on Economically Valuable Traits. EDP Sci. 2021, 254, 01020. [Google Scholar] [CrossRef]
- Costa, M.M.; Mauro, A.O.D.; Unêda-Trevisoli, S.H.; Arriel, N.H.C.; Bárbaro, I.M.; Muniz, F.R.S. Ganho Genético Por Diferentes Critérios de Seleção Em Populações Segregantes de Soja. Pesqui. Agropecu. Bras. 2004, 39, 1095–1102. [Google Scholar] [CrossRef] [Green Version]
- Shaw, D.V. Heterogeneity of Segregation Ratios from Selfed Progenies Demonstrate Polygenic Inheritance for Day Neutrality in Strawberry (Fragaria × ananassa Duch.). J. Am. Soc. Hortic. Sci. 2003, 128, 504–507. [Google Scholar] [CrossRef]
- Weebadde, C.K.; Wang, D.; Finn, C.E.; Lewers, K.S.; Luby, J.J.; Bushakra, J.; Sjulin, T.M.; Hancock, J.F. Using a Linkage Mapping Approach to Identify QTL for Day-Neutrality in the Octoploid Strawberry. Plant Breed. 2008, 127, 94–101. [Google Scholar] [CrossRef]
- Gai, J.; Lu, J. Quantitative Genetics. In Brenner’s Encyclopedia of Genetics; Elsevier: Amsterdam, The Netherlands, 2013; pp. 13–17. ISBN 978-0-08-096156-9. [Google Scholar]
- Vieira, S.D.; de Souza, D.C.; Martins, I.A.; Ribeiro, G.H.M.R.; Resende, L.; Ferraz, A.; Galvão, A.G.; Resende, J.T.V. Selection of Experimental Strawberry (Fragaria × ananassa) Hybrids Based on Selection Indices. Genet. Mol. Res. 2017, 16, gmr16019052. [Google Scholar] [CrossRef] [PubMed]
- Camargo, L.K.P.; de Resende, J.T.V.; Camargo, C.K.; Kurchaidt, S.M.; Resende, N.C.V.; Botelho, R.V. Post-Harvest Characterization of Strawberry Hybrids Obtained from the Crossing between Commercial Cultivars. Rev. Bras. Frutic. 2018, 40, /10.1590/0100–29452018821. [Google Scholar] [CrossRef] [Green Version]
- Vieira, S.D.; Araujo, A.L.; Souza, D.C.; Resende, L.V.; Leite, M.E.; Resende, J.T. Heritability and Combining Ability Studies in Strawberry Population. J. Agric. Sci. 2019, 11, 57–469. [Google Scholar] [CrossRef]
- Morales, R.G.F.; de Resende, J.T.V.; Faria, M.V.; da Silva, P.R.; Figueiredo, A.S.T.; Carminatti, R. Divergência Genética Em Cultivares de Morangueiro, Baseada Em Caracteres Morfoagronômicos. Rev. Ceres 2011, 58, 323–329. [Google Scholar] [CrossRef]
- Costa, A.F.; Ribeiro, L.P.; Teodoro, P.E.; Bhering, L.L.; Tardin, F.D.; Fornazier, M.J.; Costa, H.; Martins, D.d.S.; Junior, J.S.Z. Genetic Divergence of Strawberry Cultivars under Different Managements. Biosci. J. 2018, 34, 129–137. [Google Scholar] [CrossRef] [Green Version]
- Murti, R.H.; Kim, H.Y.; Yeoung, Y.R. Heritability of Fruit Quality in the Progenies of Day-Neutral and Short Day Hybrid Strawberry Cultivars. AGRIVITA J. Agric. Sci. 2012, 34, 105–114. [Google Scholar] [CrossRef]
- Yamamoto, E.; Kataoka, S.; Shirasawa, K.; Noguchi, Y.; Isobe, S. Genomic Selection for F1 Hybrid Breeding in Strawberry (Fragaria × ananassa). Front. Plant Sci. 2021, 12, 308. [Google Scholar] [CrossRef]
- Hancock, J.F.; Luby, J.J.; Dale, A.; Callow, P.W.; Serçe, S.; El-Shiek, A. Utilizing Wild Fragaria Virginiana in Strawberry Cultivar Development: Inheritance of Photoperiod Sensitivity, Fruit Size, Gender, Female Fertility and Disease Resistance. Euphytica 2002, 126, 177–184. [Google Scholar] [CrossRef]
- Resende, M.D.V. Genética Biométrica e Estatística No Melhoramento de Plantas Perenes; Embrapa Florestas: Colombo, Sri Lanka, 2002; ISBN 85-7383-161-8. [Google Scholar]
- Vencovsky, R. Herança Quantitativa. In Melhoramento e Produção do Milho; Fundação Cargill: Piracicaba, Brazil, 1987; pp. 137–214. [Google Scholar]
- Singh, A.; Singh, B.; Deka, B.; Sanwal, S.; Patel, R.; Verma, M. The Genetic Variability, Inheritance and Inter-Relationships of Ascorbic Acid, β-Carotene, Phenol and Anthocyanin Content in Strawberry (Fragaria × ananassa Duch.). Sci. Hortic. 2011, 129, 86–90. [Google Scholar] [CrossRef]
- Chaves, V.C.; Calvete, E.; Reginatto, F.H. Quality Properties and Antioxidant Activity of Seven Strawberry (Fragaria × ananassa Duch) Cultivars. Sci. Hortic. 2017, 225, 293–298. [Google Scholar] [CrossRef]
- Gabriel, A.; de Resende, J.T.; Zeist, A.R.; Resende, L.V.; Resende, N.C.; Zeist, R.A. Estabilidade Fenotípica de Cultivares de Morangueiro Com Base Em Características Físico-Químicas de Frutos. Hortic. Bras. 2019, 37, 75–81. [Google Scholar] [CrossRef]
- Osorio, L.; Pattison, J.; Peres, N.; Whitaker, V. Genetic Variation and Gains in Resistance of Strawberry to Colletotrichum Gloeosporioides. Phytopathology 2014, 104, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Coman, M.; Popescu, A. Inheritance of Some Strawberry Quantitative Traits. In Proceedings of the Third International Strawberry Symposium, Veldhoven, The Netherlands, 29 April–4 May 1996; pp. 81–88. [Google Scholar] [CrossRef]
- Chandler, C.K. Strawberry. In Fruit Breeding; Springer: New York, NY, USA, 2012; pp. 305–325. ISBN 978-1-4419-0763-9. [Google Scholar]
- Galvão, A.G.; Resende, L.V.; Guimaraes, R.M.; Ferraz, A.K.L.; Morales, R.G.F.; Marodin, J.C.; Catão, H.C.R.M. Overcoming Strawberry Achene Dormancy for Improved Seedling Production in Breeding Programs. Idesia 2014, 32, 57–62. [Google Scholar] [CrossRef] [Green Version]
- Alvares, C.A.; Stape, J.; Sentelhas, P.; Gonçalves, J.; Sparovek, G. Köppen’s Climate Classification Map for Brazil. Meteorol. Z. 2013, 22, 711–728. [Google Scholar] [CrossRef]
- IDR-Paraná. Dados Meteorológicos Históricos e Atuais; Instituto de Desenvolvimento Rural do Paraná—IAPAR/EMATER: Curitiba, Brazil, 2021. [Google Scholar]
- Federer, W. Augmented (or Hoonuiaku) Designs Hawaiian Planters Record. Hawaian Plant. Rec. Aica 1956, 55, 191–208. [Google Scholar]
- McGuire, R.G. Reporting of Objective Color Measurements. HortScience 1992, 27, 1254–1255. [Google Scholar] [CrossRef] [Green Version]
- Cruz, C.D. Programa GENES: Aplicativo Computacional Em Genética e Estatística Versão Windows; UFV: Viçosa, Brazil, 2006; ISBN 85-7269-245-2. [Google Scholar]
- Zimmermann, F.J.P. Estatística Aplicada à Pesquisa Agrícola, 2nd ed.; Embrapa: Brasília, Brazil, 2014; ISBN 85-7437-020-7. [Google Scholar]
- Cruz, C.D. GENES—A Software Package for Analysis in Experimental Statistics and Quantitative Genetics. Acta Sci. Agron. 2013, 35, 271–276. [Google Scholar] [CrossRef]
- Cruz, C.D. Genes Software-Extended and Integrated with the R, Matlab and Selegen. Acta Sci. 2016, 38, 547–552. [Google Scholar] [CrossRef] [Green Version]
- RStudio Team. RStudio: Integrated Development for R; RStudio PBC: Boston, MA, USA, 2020. [Google Scholar]
- Galili, T. Dendextend: An R Package for Visualizing, Adjusting and Comparing Trees of Hierarchical Clustering. Bioinformatics 2015, 31, 3718–3720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, Z.; Eils, R.; Schlesner, M. Complex Heatmaps Reveal Patterns and Correlations in Multidimensional Genomic Data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses. Available online: https://cran.microsoft.com/snapshot/2016-11-30/web/packages/factoextra/factoextra.pdf (accessed on 13 July 2021).
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Epskamp, S.; Cramer, A.O.J.; Waldorp, L.J.; Schmittmann, V.D.; Borsboom, D. Qgraph: Network Visualizations of Relationships in Psychometric Data. J. Stat. Softw. 2012, 48, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Wei, T.; Simko, V. R Package “Corrplot”: Visualization of a Correlation Matrix (Version 0.90). Available online: https://github.com/taiyun/corrplot (accessed on 4 November 2021).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genetic Parameters | TFM ** | AFM ** | CFN ** | ACFM ** | TSS ** | F ** | L ** | °Hue ** | C ** |
|---|---|---|---|---|---|---|---|---|---|
| µ * | 975.89 | 69.8 | 14.49 | 769.8 | 49.89 | 18.64 | 6.68 | 0.51 | 39.48 |
| µF | 972.72 | 69.58 | 14.78 | 781.73 | 50.1 | 19.02 | 6.66 | 9.62 | 39.85 |
| CVcontrols/genotypes (%) | 11.90 | 18.96 | 10.89 | 13.78 | 16.12 | 7.39 | 5.69 | 5.95 | 2.28 |
| CVcontrols (%) | 16.09 | 17.85 | 16.67 | 19.33 | 18.03 | 11.31 | 5.49 | 7.51 | 2.68 |
| CVgenotypes (%) | 11.67 | 19.05 | 10.62 | 13.49 | 15.99 | 7.21 | 5.71 | 5.86 | 2.26 |
| 29,096.89 | 24,564.06 | 160.38 | 7.63 | 0.31 | 0.98 | 114.72 | 344.01 | 80.25 | |
| 12,997.73 | 11,262.51 | 64.69 | 1.9 | 0.14 | 0.32 | 0.81 | 30.83 | 5.05 | |
| 16,098.97 | 13,301.55 | 95.69 | 5.73 | 0.17 | 0.66 | 113.91 | 313.17 | 75.7 | |
| (%) | 55.32 | 54.15 | 59.66 | 75.09 | 54.02 | 67.49 | 99.29 | 91.03 | 93.7 |
| (%) | 12.99 | 14.68 | 19.45 | 12.51 | 6.19 | 8.45 | 26.73 | 32.31 | 26.64 |
| 1.11 | 1.08 | 1.21 | 1.73 | 1.08 | 1.44 | 11.84 | 3.19 | 3.86 | |
| Xo | 976.67 | 785.21 | 50.19 | 19.12 | 6.66 | 9.66 | 39.91 | 54.84 | 32.55 |
| Xs | 1219.45 | 1047.56 | 65.68 | 20.07 | 6.68 | 9.94 | 52.98 | 71.66 | 37.65 |
| GG | 134.33 | 142.06 | 9.24 | 0.71 | 0.01 | 0.19 | 12.98 | 15.31 | 4.78 |
| GS% | 13.75 | 18.09 | 18.42 | 3.72 | 0.16 | 1.98 | 32.52 | 27.91 | 14.68 |
| Population | Female Parent | Male Parent | Individuals |
|---|---|---|---|
| RVFS07CR | ‘Camino Real’ | RVFS07 (‘Festival’ × ‘Sweet Charlie’) | 194 |
| RVFS06CR | ‘Camino Real’ | RVFS06 (‘Festival’ × ‘Sweet Charlie’) | 171 |
| RVCA16CR | ‘Camino Real’ | RVCA16 (‘Camarosa’ × ‘Aromas’) | 163 |
| RVCS44CR | ‘Camino Real’ | RVCS44 (‘Camarosa’ × ‘Sweet Charlie’) | 152 |
| RVDA11CR | ‘Camino Real’ | RVDA11 (‘Dover’ × ‘Aromas’) | 190 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rutz, T.; de Resende, J.T.V.; Mariguele, K.H.; Zeist, R.A.; da Silva, A.L.B.R. Selection of Short-Day Strawberry Genotypes through Multivariate Analysis. Plants 2023, 12, 2650. https://doi.org/10.3390/plants12142650
Rutz T, de Resende JTV, Mariguele KH, Zeist RA, da Silva ALBR. Selection of Short-Day Strawberry Genotypes through Multivariate Analysis. Plants. 2023; 12(14):2650. https://doi.org/10.3390/plants12142650
Chicago/Turabian StyleRutz, Thiago, Juliano Tadeu Vilela de Resende, Keny Henrique Mariguele, Ricardo Antônio Zeist, and Andre Luiz Biscaia Ribeiro da Silva. 2023. "Selection of Short-Day Strawberry Genotypes through Multivariate Analysis" Plants 12, no. 14: 2650. https://doi.org/10.3390/plants12142650
APA StyleRutz, T., de Resende, J. T. V., Mariguele, K. H., Zeist, R. A., & da Silva, A. L. B. R. (2023). Selection of Short-Day Strawberry Genotypes through Multivariate Analysis. Plants, 12(14), 2650. https://doi.org/10.3390/plants12142650