
Protective Activity of Rhizobium leguminosarum bv. viciae Strain 33504-Mat209 against Alfalfa Mosaic Virus Infection in Faba Bean Plants

 ,
, 
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Bacterial Isolation and Identification
2.2. The AMV-CP Accumulation Level and Disease Severity Evaluation
2.3. Evaluation of Plants’ Growth Parameters and Total Chlorophyll Contents
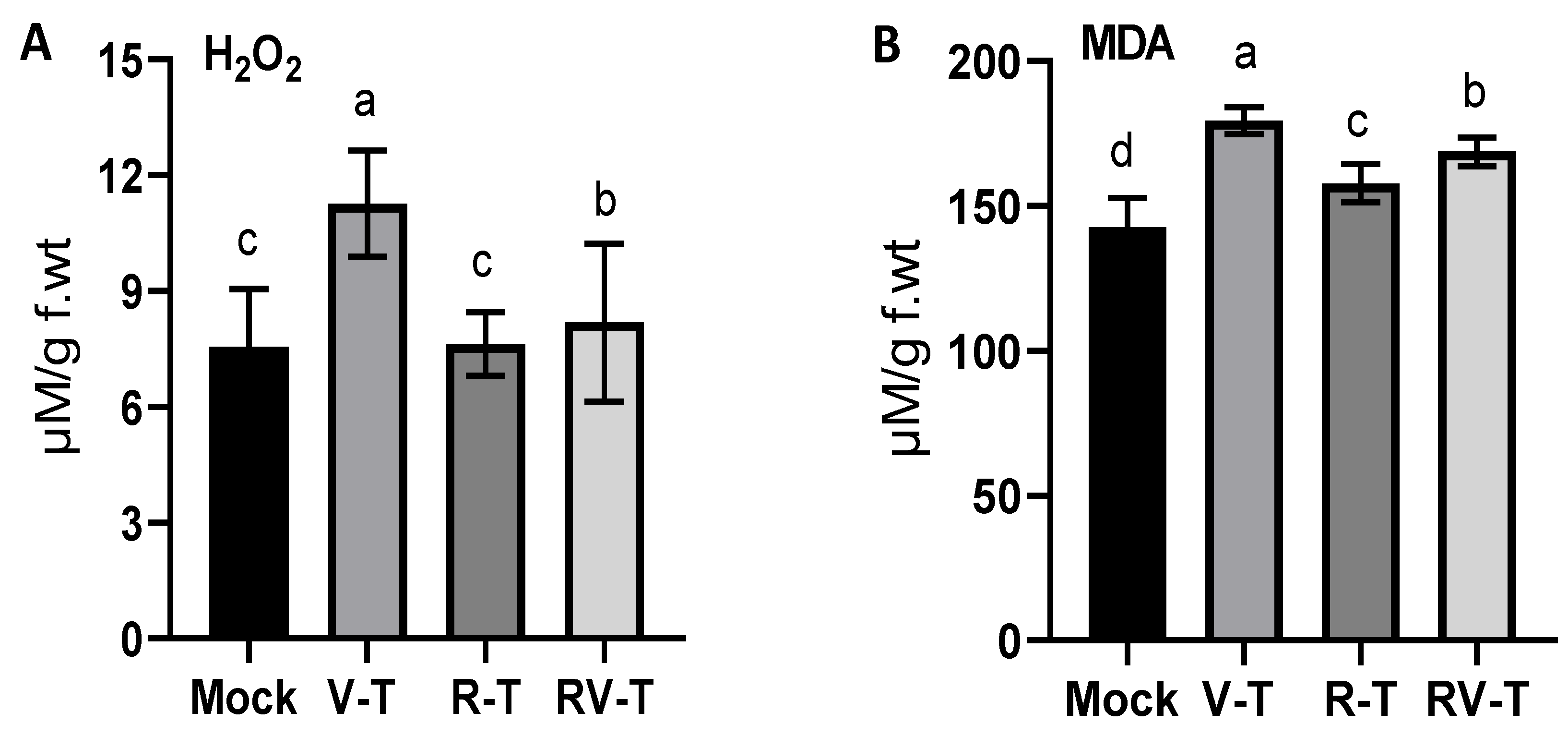
2.4. Evaluation of Oxidative Stress Markers
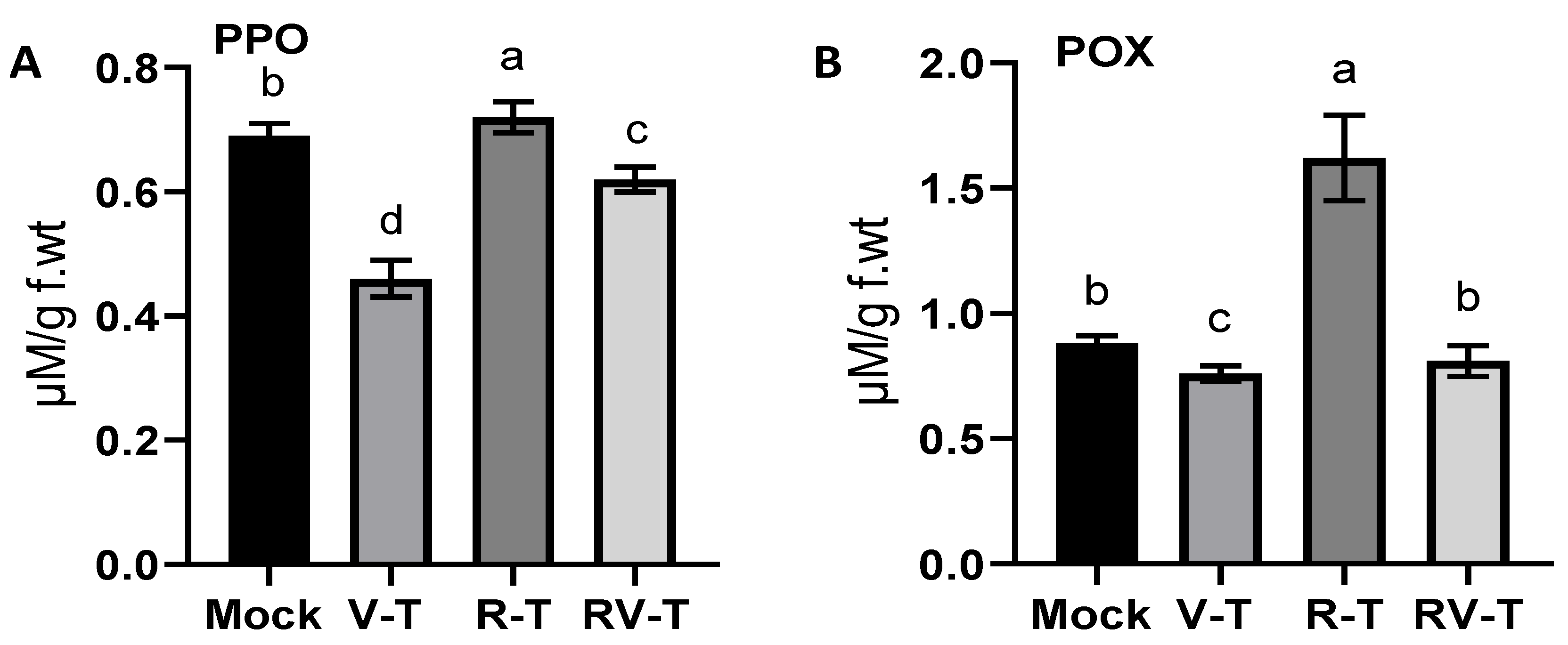
2.5. Antioxidant Enzymes Evaluation
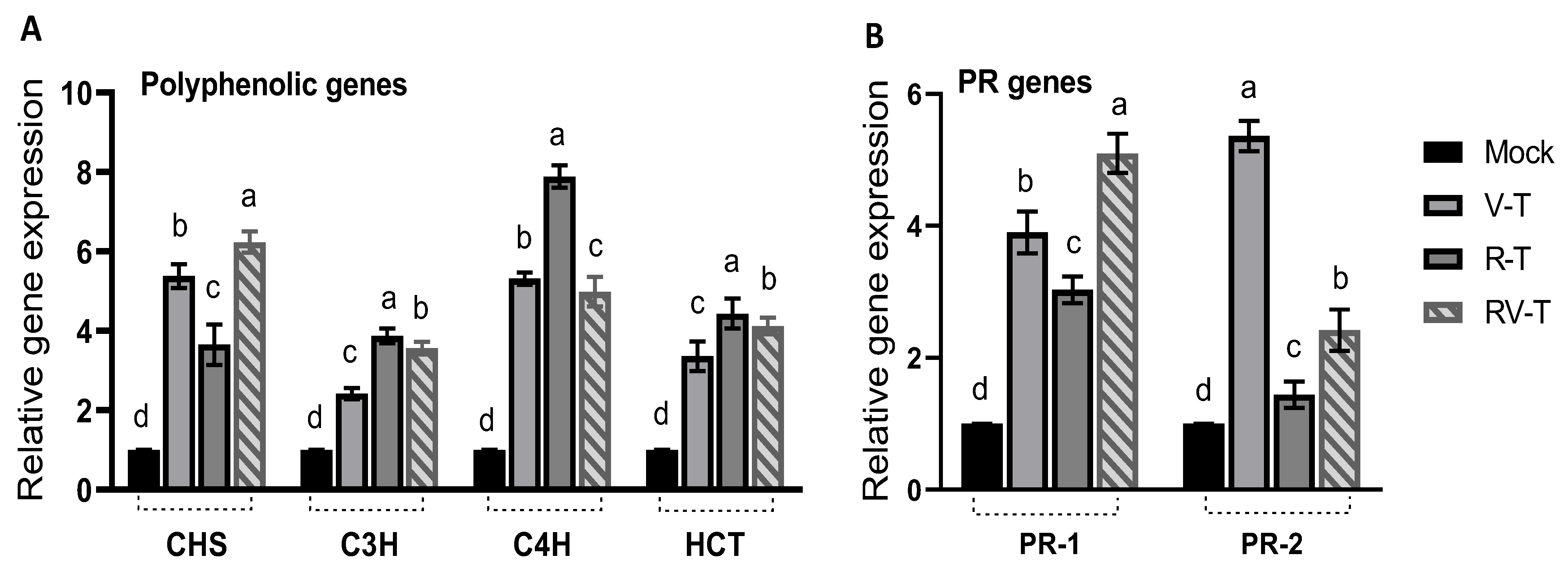
2.6. Effect of 33504-Mat209 on the Transcriptional Levels of Defense-Related Genes
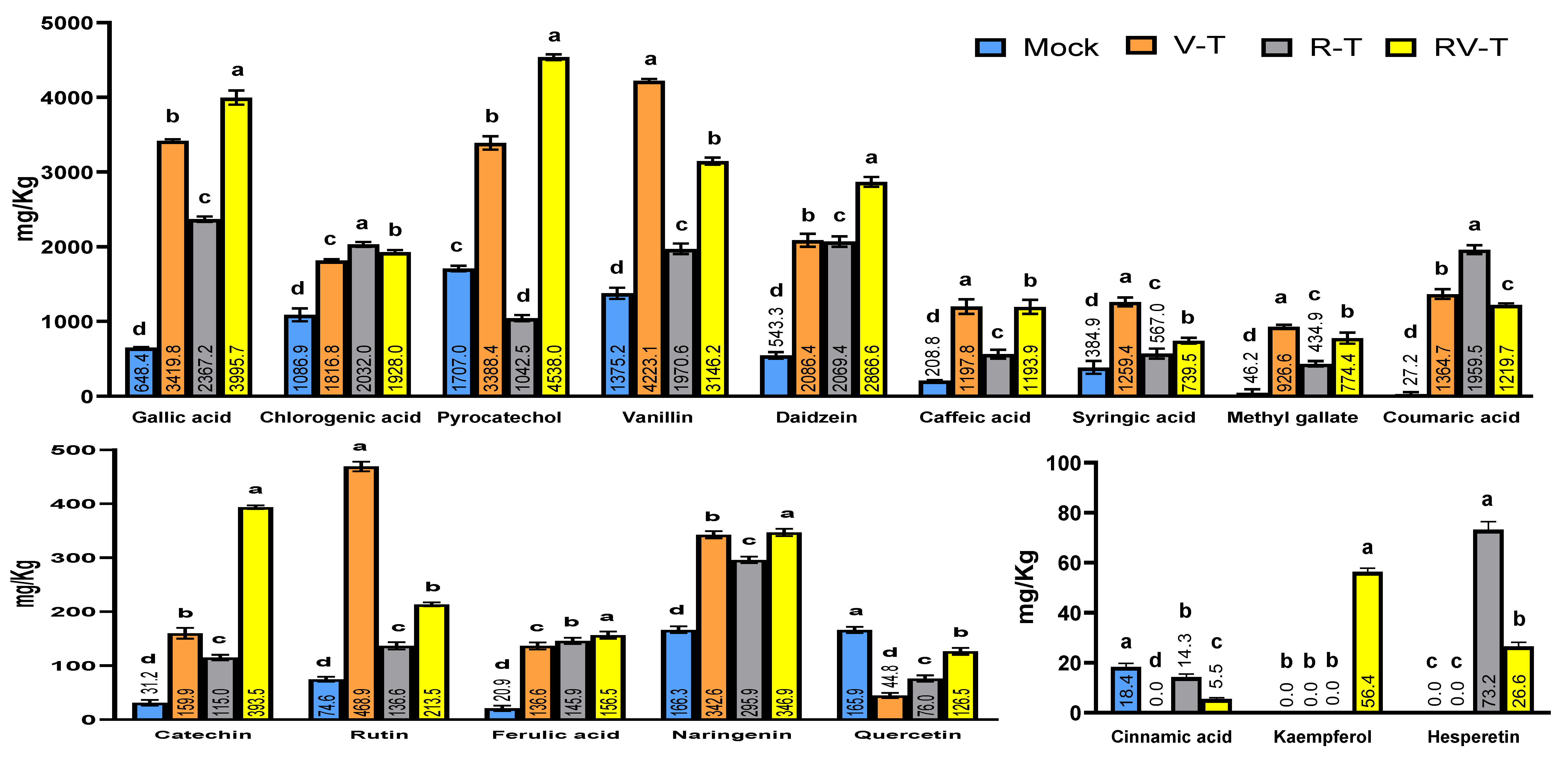
2.7. Polyphenolic Compounds Analysis through HPLC
3. Discussion
4. Materials and Methods
4.1. Plant Material and Viral Inoculum Source
4.2. Isolation and Characterization of the Most Potent Bacterial Isolate
4.3. Inoculum Preparation and Greenhouse Experimental Design
4.4. Virus Accumulation Level and Disease Assessment
4.5. Evaluation of Plants’ Growth Parameters
4.6. Total Chlorophyll Determination
4.7. Determination of Oxidation Stress Conditions (H2O2 and MDA)
4.8. Antioxidant Enzyme Evaluation
4.8.1. Peroxidase Activity (POX)
4.8.2. Polyphenol Oxidase Activity (PPO)
4.9. Evaluation of the Relative Expression of Some Defense-Related Genes
4.10. Evaluation of Polyphenolic Phytochemicals Contents through HPLC
4.11. Statistical Significance and Analysis
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cui, Y.; Ji, Y.; Liu, R.; Li, W.; Liu, Y.; Liu, Z.; Zong, X.; Yang, T. Faba Bean (Vicia faba L.) Yield Estimation Based on Dual-Sensor Data. Drones 2023, 7, 378. [Google Scholar] [CrossRef]
- Alnefaie, R.M.; El-Sayed, S.A.; Ramadan, A.A.; Elmezien, A.I.; El-Taher, A.M.; Randhir, T.O.; Bondok, A. Physiological and anatomical responses of faba bean plants infected with chocolate spot disease to chemical inducers. Life 2023, 13, 392. [Google Scholar] [CrossRef]
- Rose, T.J.; Rose, M.T.; Pariasca-Tanaka, J.; Heuer, S.; Wissuwa, M. The frustration with utilization: Why have improvements in internal phosphorus utilization efficiency in crops remained so elusive? Front. Plant Sci. 2011, 2, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashem, A.H.; Abdelaziz, A.M.; Askar, A.A.; Fouda, H.M.; Khalil, A.M.A.; Abd-Elsalam, K.A.; Khaleil, M.M. Bacillus megaterium-mediated synthesis of selenium nanoparticles and their antifungal activity against Rhizoctonia solani in faba bean plants. J. Fungi 2021, 7, 195. [Google Scholar] [CrossRef] [PubMed]
- Abdelkhalek, A.; Yassin, Y.; Abdel-Megeed, A.; Abd-Elsalam, K.A.; Moawad, H.; Behiry, S.I. Rhizobium leguminosarum bv. viciae-Mediated Silver Nanoparticles for Controlling Bean Yellow Mosaic Virus (BYMV) Infection in Faba Bean Plants. Plants 2023, 12, 45. [Google Scholar] [CrossRef] [PubMed]
- Chatzivassiliou, E.K. An annotated list of legume-infecting viruses in the light of metagenomics. Plants 2021, 10, 1413. [Google Scholar] [CrossRef]
- Abdelkhalek, A.; El-Gendi, H.; Al-Askar, A.A.; Maresca, V.; Moawad, H.; Elsharkawy, M.M.; Younes, H.A.; Behiry, S.I. Enhancing systemic resistance in faba bean (Vicia faba L.) to Bean yellow mosaic virus via soil application and foliar spray of nitrogen-fixing Rhizobium leguminosarum bv. viciae strain 33504-Alex1. Front. Plant Sci. 2022, 13, 933498. [Google Scholar] [CrossRef] [PubMed]
- Nair, R.M.; Habili, N.; Randles, J.W. Infection of Cullen australasicum (syn. Psoralea australasica) with Alfalfa mosaic virus. Australas. Plant Dis. Notes 2009, 4, 46–48. [Google Scholar]
- Mahmoud, S.Y.M.; Khaled, A.-S.G.A.; Petrzik, K. Differentiation study between Alfalfa Mosaic Virus and Red Clover Mottle Virus affecting broad bean by biological and molecular characterization. Int. J. Virol. 2010, 6, 224–239. [Google Scholar] [CrossRef]
- Abdelkhalek, A.; Hafez, E. Plant Viral Diseases in Egypt and Their Control. In Cottage Industry of Biocontrol Agents and Their Applications; Springer: Berlin/Heidelberg, Germany, 2020; pp. 403–421. [Google Scholar]
- Piccoli, C.; Cremonese, C.; Koifman, R.J.; Koifman, S.; Freire, C. Pesticide exposure and thyroid function in an agricultural population in Brazil. Environ. Res. 2016, 151, 389–398. [Google Scholar] [CrossRef]
- Bass, C.; Denholm, I.; Williamson, M.S.; Nauen, R. The global status of insect resistance to neonicotinoid insecticides. Pestic. Biochem. Physiol. 2015, 121, 78–87. [Google Scholar] [CrossRef] [Green Version]
- Franco-Andreu, L.; Gómez, I.; Parrado, J.; García, C.; Hernández, T.; Tejada, M. Behavior of two pesticides in a soil subjected to severe drought. Effects on soil biology. Appl. Soil Ecol. 2016, 105, 17–24. [Google Scholar] [CrossRef]
- Rizvi, A.; Ahmed, B.; Khan, M.S.; El-Beltagi, H.S.; Umar, S.; Lee, J. Bioprospecting Plant Growth Promoting Rhizobacteria for Enhancing the Biological Properties and Phytochemical Composition of Medicinally Important Crops. Molecules 2022, 27, 1407. [Google Scholar] [CrossRef] [PubMed]
- Karavidas, I.; Ntatsi, G.; Vougeleka, V.; Karkanis, A.; Ntanasi, T.; Saitanis, C.; Agathokleous, E.; Ropokis, A.; Sabatino, L.; Tran, F.; et al. Agronomic Practices to Increase the Yield and Quality of Common Bean (Phaseolus vulgaris L.): A Systematic Review. Agronomy 2022, 12, 271. [Google Scholar] [CrossRef]
- Al-Ani, R.A.; Adhab, M.A. Bean Yellow Mosaic Virus (BYMV) on broadbean: Characterization and resistance induced by Rhizobium leguminosarum. J. Pure Appl. Microbiol. 2013, 7, 135–142. [Google Scholar]
- Vejan, P.; Abdullah, R.; Khadiran, T.; Ismail, S.; Nasrulhaq Boyce, A. Role of plant growth promoting rhizobacteria in agricultural sustainability-A review. Molecules 2016, 21, 573. [Google Scholar] [CrossRef] [Green Version]
- Abdelkhalek, A.; Al-Askar, A.A.; Behiry, S.I. Bacillus licheniformis strain POT1 mediated polyphenol biosynthetic pathways genes activation and systemic resistance in potato plants against Alfalfa mosaic virus. Sci. Rep. 2020, 10, 16120. [Google Scholar] [CrossRef]
- Abdelkhalek, A.; Behiry, S.I.; Al-Askar, A.A. Bacillus velezensis PEA1 Inhibits Fusarium oxysporum Growth and Induces Systemic Resistance to Cucumber Mosaic Virus. Agronomy 2020, 10, 1312. [Google Scholar] [CrossRef]
- Moawad, H.; Abd el-Rahim, W.M.; Abd el-Aleem, D.; Abo Sedera, S.A. Persistence of two Rhizobium etli inoculant strains in clay and silty loam soils. J. Basic Microbiol. An Int. J. Biochem. Physiol. Genet. Morphol. Ecol. Microorg. 2005, 45, 438–446. [Google Scholar]
- Saeed, Q.; Xiukang, W.; Haider, F.U.; Kučerik, J.; Mumtaz, M.Z.; Holatko, J.; Naseem, M.; Kintl, A.; Ejaz, M.; Naveed, M.; et al. Rhizosphere bacteria in plant growth promotion, biocontrol, and bioremediation of contaminated sites: A comprehensive review of effects and mechanisms. Int. J. Mol. Sci. 2021, 22, 10529. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.G.; Makkouk, K.M. Virus diseases of faba bean (Vicia faba L.) in Asia and Africa. Plant Viruses 2007, 1, 93–105. [Google Scholar]
- Akgun, D.; Canci, H. Selection of Faba Bean (Vicia faba L.) Genotypes for High Yield, Essential Amino Acids and Low Anti-Nutritional Factors. Agriculture 2023, 13, 932. [Google Scholar] [CrossRef]
- Al-Saleh, M.A.; Amer, M.A. Biological and molecular variability of Alfalfa mosaic virus affecting alfalfa crop in Riyadh region. Plant Pathol. J. 2013, 29, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Abdelkhalek, A.; Al-Askar, A.A.; Hafez, E. Differential induction and suppression of the potato innate immune system in response to Alfalfa mosaic virus infection. Physiol. Mol. Plant Pathol. 2020, 110, 101485. [Google Scholar] [CrossRef]
- Logan, N.A.; De Vos, P. Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley & Sons: Hoboken, NJ, USA, 2015. [Google Scholar] [CrossRef]
- Elhelaly, S.H. The effectiveness of the bacteria Rhizobium leguminosarum against bean yellow mosaic (BYMV) potyvirus infecting faba bean (Vicia faba L.) Plants. Ann. Agric. Sci. Moshtohor 2022, 60, 181–190. [Google Scholar] [CrossRef]
- El-Gendi, H.; Al-Askar, A.A.; Király, L.; Samy, M.A.; Moawad, H.; Abdelkhalek, A. Foliar Applications of Bacillus subtilis HA1 Culture Filtrate Enhance Tomato Growth and Induce Systemic Resistance against Tobacco mosaic virus Infection. Horticulturae 2022, 8, 301. [Google Scholar] [CrossRef]
- Zhu, F.; Zhang, Q.P.; Che, Y.P.; Zhu, P.X.; Zhang, Q.Q.; Ji, Z.L. Glutathione contributes to resistance responses to TMV through a differential modulation of salicylic acid and reactive oxygen species. Mol. Plant Pathol. 2021, 22, 1668–1687. [Google Scholar] [CrossRef]
- Balal, R.M.; Khan, M.M.; Shahid, M.A.; Mattson, N.S.; Abbas, T.; Ashfaq, M.; Garcia-Sanchez, F.; Ghazanfer, U.; Gimeno, V.; Iqbal, Z. Comparative studies on the physiobiochemical, enzymatic, and ionic modifications in salt-tolerant and salt-sensitive citrus rootstocks under NaCl stress. J. Am. Soc. Hortic. Sci. 2012, 137, 86–95. [Google Scholar] [CrossRef] [Green Version]
- Alché, J. de D. A concise appraisal of lipid oxidation and lipoxidation in higher plants. Redox Biol. 2019, 23, 101136. [Google Scholar] [CrossRef]
- Mathioudakis, M.M.; Veiga, R.S.L.; Canto, T.; Medina, V.; Mossialos, D.; Makris, A.M.; Livieratos, I. P epino mosaic virus triple gene block protein 1 (TGBp1) interacts with and increases tomato catalase 1 activity to enhance virus accumulation. Mol. Plant Pathol. 2013, 14, 589–601. [Google Scholar] [CrossRef]
- Abdelkhalek, A.; Aseel, D.G.; Király, L.; Künstler, A.; Moawad, H.; Al-Askar, A.A. Induction of Systemic Resistance to Tobacco mosaic virus in Tomato through Foliar Application of Bacillus amyloliquefaciens Strain TBorg1 Culture Filtrate. Viruses 2022, 14, 1830. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, M.; Kazemi, H. Changes in peroxidase and polyphenol oxidase activities in susceptible and resistant wheat heads inoculated with Fusarium graminearum and induced resistance. Plant Sci. 2002, 162, 491–498. [Google Scholar] [CrossRef]
- Abdelkhalek, A.; Al-Askar, A.A.; Arishi, A.A.; Behiry, S.I. Trichoderma hamatum Strain Th23 Promotes Tomato Growth and Induces Systemic Resistance against Tobacco Mosaic Virus. J. Fungi 2022, 8, 228. [Google Scholar] [CrossRef] [PubMed]
- Van Loon, L.C.; Van Strien, E.A. The families of pathogenesis-related proteins, their activities, and comparative analysis of PR-1 type proteins. Physiol. Mol. Plant Pathol. 1999, 55, 85–97. [Google Scholar] [CrossRef]
- Dempsey, D.A.; Vlot, A.C.; Wildermuth, M.C.; Klessig, D.F.; D’Maris Amick Dempsey, A.C.; Vlot, M.C.W.; Daniel, F.K.; Dempsey, D.A.; Vlot, A.C.; Wildermuth, M.C.; et al. Salicylic acid biosynthesis and metabolism. Arab. Book/Am. Soc. Plant Biol. 2011, 9, e0156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoegen, E.; Strömberg, A.; Pihlgren, U.; Kombrink, E. Primary structure and tissue-specific expression of the pathogenesis-related protein PR-1b in potato. Mol. Plant Pathol. 2002, 3, 329–345. [Google Scholar] [CrossRef]
- Abdelkhalek, A.; Qari, S.H.; Abu-Saied, M.A.A.-R.; Khalil, A.M.; Younes, H.A.; Nehela, Y.; Behiry, S.I. Chitosan Nanoparticles Inactivate Alfalfa Mosaic Virus Replication and Boost Innate Immunity in Nicotiana glutinosa Plants. Plants 2021, 10, 2701. [Google Scholar] [CrossRef]
- Kawakami, S.; Watanabe, Y.; Beachy, R.N. Tobacco mosaic virus infection spreads cell to cell as intact replication complexes. Proc. Natl. Acad. Sci. USA 2004, 101, 6291–6296. [Google Scholar] [CrossRef]
- Otulak, K.; Garbaczewska, G. Cellular localisation of calcium ions during potato hypersensitive response to Potato virus Y. Micron 2011, 42, 381–391. [Google Scholar] [CrossRef]
- Abdelkhalek, A. Expression of tomato pathogenesis related genes in response to Tobacco mosaic virus. JAPS J. Anim. Plant Sci. 2019, 29, 1596–1602. [Google Scholar]
- Otulak-Kozieł, K.; Kozieł, E.; Lockhart, B. Plant cell wall dynamics in compatible and incompatible potato response to infection caused by Potato virus Y (PVYNTN). Int. J. Mol. Sci. 2018, 19, 862. [Google Scholar] [CrossRef] [Green Version]
- Akyol, H.; Riciputi, Y.; Capanoglu, E.; Caboni, M.; Verardo, V. Phenolic compounds in the potato and its byproducts: An overview. Int. J. Mol. Sci. 2016, 17, 835. [Google Scholar] [CrossRef] [Green Version]
- André, C.M.; Schafleitner, R.; Legay, S.; Lefèvre, I.; Aliaga, C.A.A.; Nomberto, G.; Hoffmann, L.; Hausman, J.-F.; Larondelle, Y.; Evers, D. Gene expression changes related to the production of phenolic compounds in potato tubers grown under drought stress. Phytochemistry 2009, 70, 1107–1116. [Google Scholar] [CrossRef]
- Abdelkhalek, A.; Dessoky, E.S.E.S.; Hafez, E. Polyphenolic genes expression pattern and their role in viral resistance in tomato plant infected with Tobacco mosaic virus. Biosci. Res. 2018, 15, 3349–3356. [Google Scholar]
- Yan, Q.; Si, J.; Cui, X.; Peng, H.; Chen, X.; Xing, H.; Dou, D. The soybean cinnamate 4-hydroxylase gene GmC4H1 contributes positively to plant defense via increasing lignin content. Plant Growth Regul. 2019, 88, 139–149. [Google Scholar] [CrossRef]
- Hoffmann, L.; Maury, S.; Martz, F.; Geoffroy, P.; Legrand, M. Purification, cloning, and properties of an acyltransferase controlling shikimate and quinate ester intermediates in phenylpropanoid metabolism. J. Biol. Chem. 2003, 278, 95–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leiss, K.A.; Maltese, F.; Choi, Y.H.; Verpoorte, R.; Klinkhamer, P.G.L. Identification of chlorogenic acid as a resistance factor for thrips in chrysanthemum. Plant Physiol. 2009, 150, 1567–1575. [Google Scholar] [CrossRef] [Green Version]
- Tsao, R.; Marvin, C.H.; Broadbent, A.B.; Friesen, M.; Allen, W.R.; Mcgarvey, B.D. Evidence for an isobutylamide associated with host-plant resistance to western flower thrips, Frankliniella occidentalis, in chrysanthemum. J. Chem. Ecol. 2005, 31, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Dao, T.T.H.; Linthorst, H.J.M.; Verpoorte, R. Chalcone synthase and its functions in plant resistance. Phytochem. Rev. 2011, 10, 397–412. [Google Scholar] [CrossRef] [Green Version]
- Martínez, G.; Regente, M.; Jacobi, S.; Del Rio, M.; Pinedo, M.; de la Canal, L. Chlorogenic acid is a fungicide active against phytopathogenic fungi. Pestic. Biochem. Physiol. 2017, 140, 30–35. [Google Scholar] [CrossRef]
- Kanwal, Q.; Hussain, I.; Latif Siddiqui, H.; Javaid, A. Antifungal activity of flavonoids isolated from mango (Mangifera indica L.) leaves. Nat. Prod. Res. 2010, 24, 1907–1914. [Google Scholar] [CrossRef] [PubMed]
- Likić, S.; Šola, I.; Ludwig-Müller, J.; Rusak, G. Involvement of kaempferol in the defence response of virus infected Arabidopsis thaliana. Eur. J. Plant Pathol. 2014, 138, 257–271. [Google Scholar] [CrossRef]
- Kubo, S.; Ikeda, T.; Imaizumi, S.; Takanami, Y.; Mikami, Y. A potent plant virus inhibitor found in Mirabilis jalapa L. Jpn. J. Phytopathol. 1990, 56, 481–487. [Google Scholar] [CrossRef]
- Somasegaran, P.; Hoben, H.J. Handbook for Rhizobia: Methods in Legume-Rhizobium Technology; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012; ISBN 1461383757. [Google Scholar]
- Strain, H.H.; Svec, W.A. Extraction, separation, estimation, and isolation of the chlorophylls. In The Chlorophylls; Elsevier: Amsterdam, The Netherlands, 1966; pp. 21–66. [Google Scholar]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Angelini, R.; Manes, F.; Federico, R. Spatial and functional correlation between diamine-oxidase and peroxidase activities and their dependence upon de-etiolation and wounding in chick-pea stems. Planta 1990, 182, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.K.; Ahn, H.K. Purification and characterization of polyphenol oxidase from potato: II. Inhibition and catalytic mechanism. J. Food Biochem. 1999, 23, 593–605. [Google Scholar] [CrossRef]
- AbdEl-Rahim, W.M.; Khalil, W.K.B.; Eshak, M.G. Evaluation of the gene expression changes in Nile tilapia (Oreochromis niloticus) as affected by the bio-removal of toxic textile dyes from aqueous solution in small-scale bioreactor. Environmentalist 2010, 30, 242–253. [Google Scholar] [CrossRef]
- Hafez, E.E.; Abdelkhalek, A.A.; Abd El-Wahab, A.S.E.D.; Galal, F.H. Altered gene expression: Induction/suppression in leek elicited by Iris Yellow Spot Virus infection (IYSV) Egyptian isolate. Biotechnol. Biotechnol. Equip. 2013, 27, 4061–4068. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author (s) and contributor (s) and not of MDPI and/or the editor (s). MDPI and/or the editor (s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Shoot | Root | Total Chlorophyll Content | ||||
|---|---|---|---|---|---|---|---|
| Length (cm) | Fresh Weight (g) | Dry Weight (g) | Length (cm) | Fresh Weight (g) | Dry Weight (g) | ||
| Mock | 37.2 ± 1.3 b | 7.23 ± 0.9 b | 2.98 ± 0.3 b | 20.1 ± 1.5 c | 5.89 ± 1.2 c | 2.21 ± 0.9 c | 26.20 ± 1.2 b |
| V-T | 29.9 ± 1.5 d | 6.32 ± 0.8 d | 2.11 ± 0.5 d | 14.3 ± 1.8 d | 4.58 ± 1.1 d | 1.67 ± 0.9 d | 20.18 ± 1.4 d |
| R-T | 42.3 ± 2.0 a | 8.14 ± 1.1 a | 3.16 ± 0.9 a | 29.2 ± 2.3 a | 6.76 ± 1.0 a | 2.97 ± 1.0 a | 30.81 ± 1.7 a |
| RV-T | 34.2 ± 1.6 c | 6.73 ± 1.1 c | 2.72 ± 0.4 c | 24.6 ± 2.1 b | 6.23 ± 1.1 b | 2.57 ± 0.8 b | 25.65 ± 1.6 c |
| Primer Name | Abbreviation | Direction | Nucleotide Sequence |
|---|---|---|---|
| 16S ribosomal RNA | 16S rRNA | Forward | AGAGTTTGATCCTGGCTCAG |
| Reverse | GGTTACCTTGTTACGACTT | ||
| Alfalfa mosaic virus-coat protein | AMV-CP | Forward | CCATCATGAGTTCTTCACAAAAG |
| Reverse | TCGTCACGTCATCAGTGAGAC | ||
| Pathogenesis related protein-1 | PR-1 | Forward | GTTCCTCCTTGCCACCTTC |
| Reverse | TATGCACCCCCAGCATAGTT | ||
| Endoglucanase | PR-2 | Forward | TATAGCCGTTGGAAACGAAG |
| Reverse | CAACTTGCCATCACATTCTG | ||
| Cinnamate 4-hydroxylase | C4H | Forward | CCCAGTTTTTG AAA TTG GCT TCA |
| Reverse | GCCCCATTCTAA GCA AGA GAA CAT C | ||
| Hydroxycinnamoyl transferase | HCT | Forward | TCT CCA ACC CCT TTT AAC GAACC |
| Reverse | CAA CTT GTC CTT CTA CCA CAG GGA A | ||
| p-coumarate 3-hydroxylase | C3H | Forward | TTG GTG GCT ACG ACA TTC CTA AGG |
| Reverse | GGT CTG AAC TCC AAT GGG TTA TTC C | ||
| Chalcone synthase | CHS | Forward | CAC CGT GGA GGA GTA TCG TAA GGC |
| Reverse | TGA TCA ACA CAG TTG GAA GGCG | ||
| Actin | Actin | Forward | GTTAGCAACTGGGATGACAT |
| Reverse | GTTACGACCACTAGCATAGAGTG | ||
| Elongation factor 1-alpha | EF1-α | Forward | GTGAAGCCCGGTATGCTTGT |
| Reverse | CTTGAGATCCTTGACTGCAACATT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdelkhalek, A.; Bashir, S.; El-Gendi, H.; Elbeaino, T.; El-Rahim, W.M.A.; Moawad, H. Protective Activity of Rhizobium leguminosarum bv. viciae Strain 33504-Mat209 against Alfalfa Mosaic Virus Infection in Faba Bean Plants. Plants 2023, 12, 2658. https://doi.org/10.3390/plants12142658
Abdelkhalek A, Bashir S, El-Gendi H, Elbeaino T, El-Rahim WMA, Moawad H. Protective Activity of Rhizobium leguminosarum bv. viciae Strain 33504-Mat209 against Alfalfa Mosaic Virus Infection in Faba Bean Plants. Plants. 2023; 12(14):2658. https://doi.org/10.3390/plants12142658
Chicago/Turabian StyleAbdelkhalek, Ahmed, Shimaa Bashir, Hamada El-Gendi, Toufic Elbeaino, Wafaa M. Abd El-Rahim, and Hassan Moawad. 2023. "Protective Activity of Rhizobium leguminosarum bv. viciae Strain 33504-Mat209 against Alfalfa Mosaic Virus Infection in Faba Bean Plants" Plants 12, no. 14: 2658. https://doi.org/10.3390/plants12142658
APA StyleAbdelkhalek, A., Bashir, S., El-Gendi, H., Elbeaino, T., El-Rahim, W. M. A., & Moawad, H. (2023). Protective Activity of Rhizobium leguminosarum bv. viciae Strain 33504-Mat209 against Alfalfa Mosaic Virus Infection in Faba Bean Plants. Plants, 12(14), 2658. https://doi.org/10.3390/plants12142658