
Evaluation of Climate Change Impacts on the Potential Distribution of Wild Radish in East Asia



Abstract
:1. Introduction
2. Materials and Methods
2.1. Species Occurrence Data
2.2. Environmental Factors
2.3. MaxEnt Model Accuracy Verification
2.4. Suitable Habitat Grade Classification
3. Results
3.1. Evaluation of MaxEnt Model Prediction Accuracy
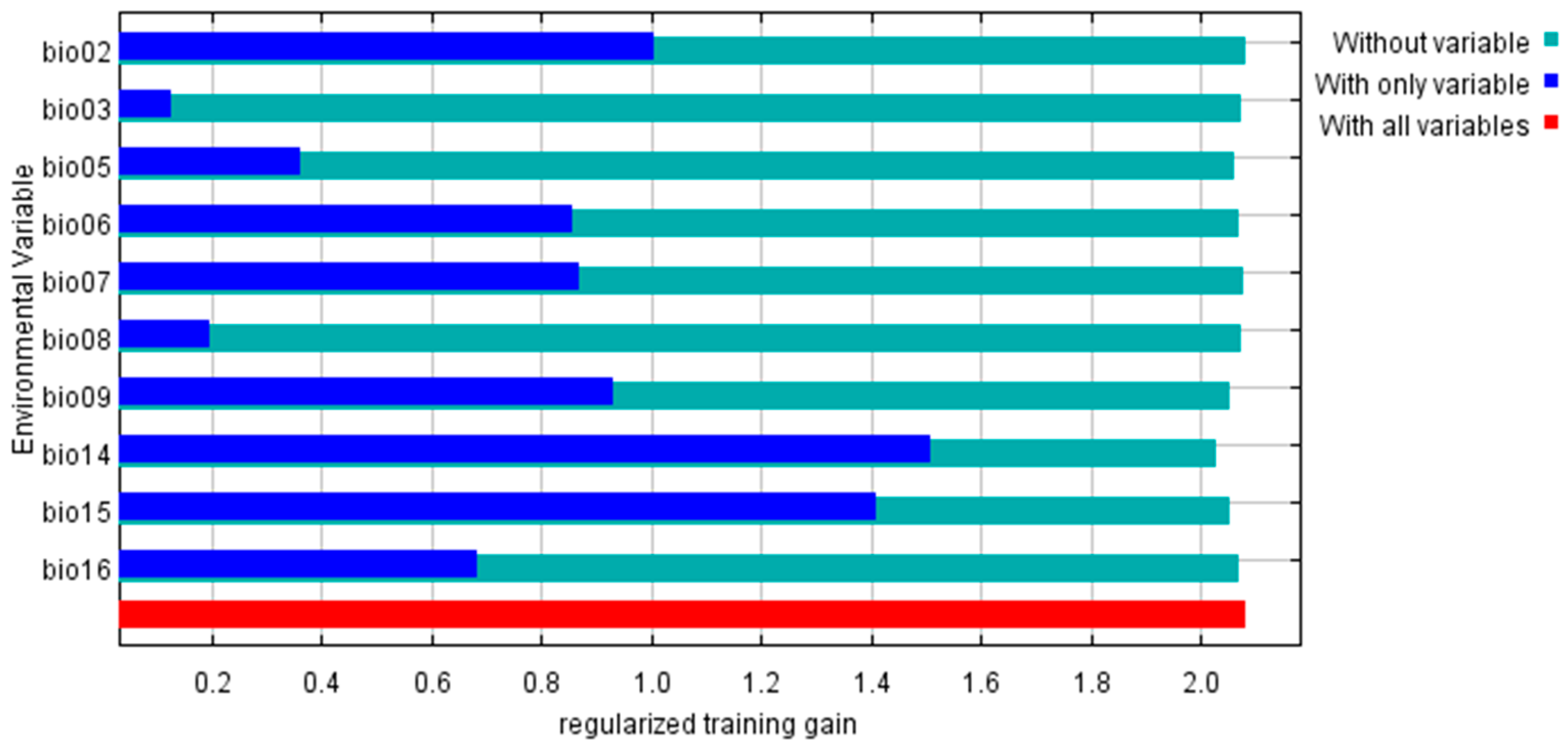
3.2. Evaluation of the Importance of Climatic Variables
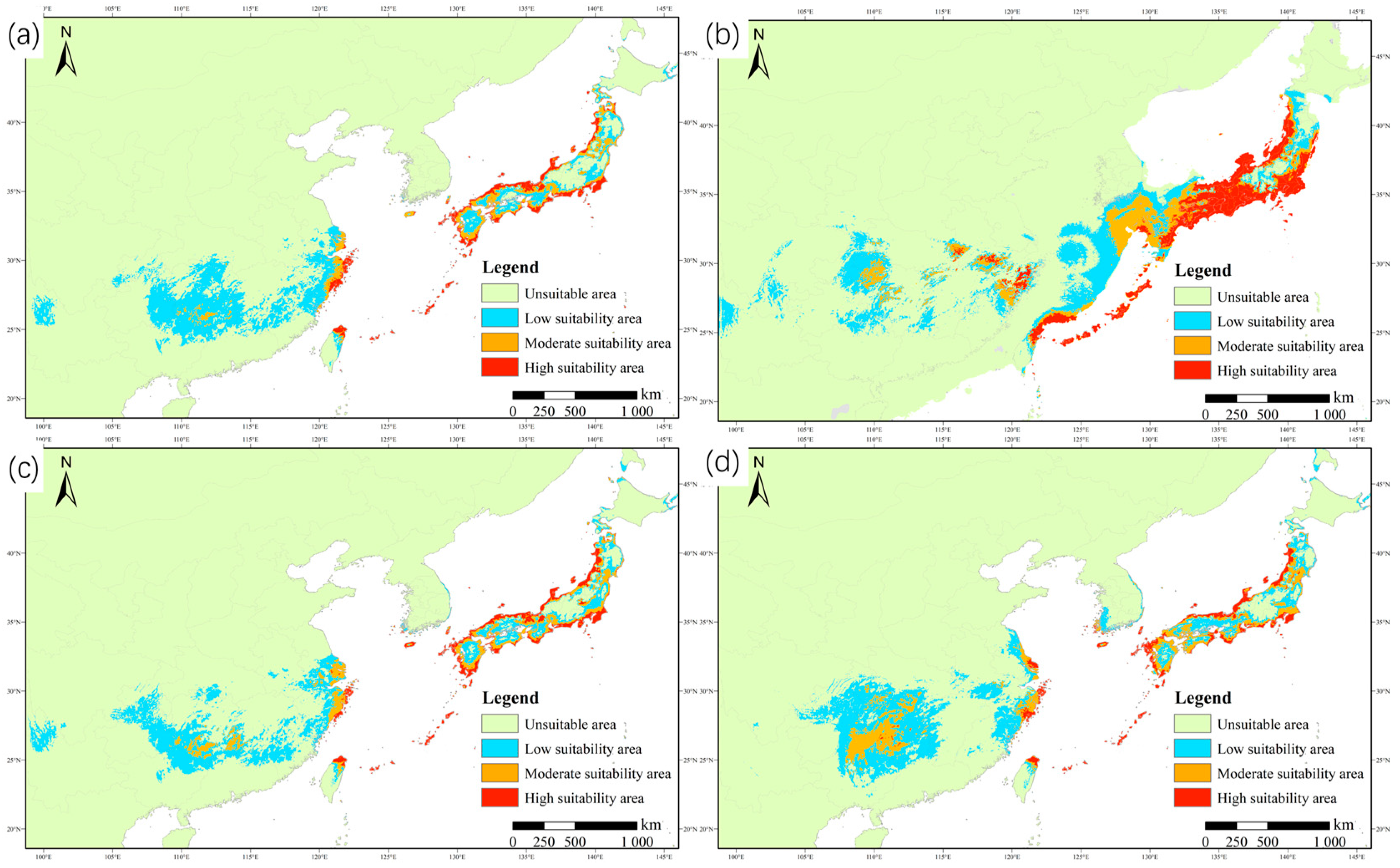
3.3. Distribution and Change in Potentially Suitable Habitat under Different Climate Conditions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Van der Wiel, K.; Bintanja, R. Contribution of climatic changes in mean and variability to monthly temperature and precipitation extremes. Commun. Earth Environ. 2021, 2, 1. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2023: Synthesis Report; IPCC: Geneva, Switzerland, 2023. [Google Scholar]
- Piao, S.; Wang, X. Biological systems under climate change: What do we learn from the IPCC AR6. Glob. Change Biol. 2023, 29, 5120–5121. [Google Scholar] [CrossRef] [PubMed]
- Lane, A.; Jarvis, A. Changes in climate will modify the geography of crop suitability: Agricultural biodiversity can help with adaptation. J. SAT Agric. Res. 2007, 4, 409–432. [Google Scholar] [CrossRef]
- Jarvis, A.; Lane, A.; Hijmans, R.J. The effect of climate change on crop wild relatives. Agric. Ecosyst. Environ. 2008, 126, 13–23. [Google Scholar] [CrossRef]
- Kapazoglou, A.; Gerakari, M.; Lazaridi, E.; Kleftogianni, K.; Sarri, E.; Tani, E.; Bebeli, P.J. Crop wild relatives: A valuable source of tolerance to various abiotic stresses. Plants 2023, 12, 328. [Google Scholar] [CrossRef] [PubMed]
- Castañeda-Álvarez, N.P.; Khoury, C.K.; Achicanoy, H.A.; Bernau, V.; Dempewolf, H.; Eastwood, R.J.; Guarino, L.; Harker, R.H.; Jarvis, A.; Maxted, N.; et al. Global conservation priorities for crop wild relatives. Nat. Plants 2016, 2, 16022. [Google Scholar] [CrossRef]
- Gutiérrez, R.M.P.; Perez, R.L. Raphanus sativus (Radish): Their chemistry and biology. Sci. World J. 2004, 4, 811–837. [Google Scholar] [CrossRef]
- Swaamy, K.R.M. Origin, distribution, genetic diversity and bressding of radish (Raphanus sativus L.). Int. J. Develop. Res. 2023, 13, 61657–61673. [Google Scholar]
- Govaerts, R.; Nic, L.E.; Black, N.; Turner, R.; Paton, A. The world checklist of vascular plants, a continuously updated resource for exploring global plant diversity. Sci. Data 2021, 8, 215. [Google Scholar] [CrossRef]
- Han, Q.; Higashi, H.; Mitsui, Y.; Setoguchi, H. Distinct phylogeographic structures of wild radish (Raphanus sativus L. var. raphanistroides Makino) in Japan. PLoS ONE 2015, 10, e0135132. [Google Scholar] [CrossRef]
- Manzoor, A.; Bashir, M.A.; Naveed, M.S.; Cheema, K.L.; Cardarelli, M. Role of different abiotic factors in inducing pre-harvest physiological disorders in radish (Raphanus sativus). Plants. 2021, 10, 2003. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, Y.; Matsuzawa, Y. Radish: Raphanus sativus L. In Genetic Improvement of Vegetable Crops; Elsevier: Amsterdam, The Netherlands, 1993; pp. 487–510. [Google Scholar] [CrossRef]
- Yukio, K.; Chiaki, K.T.; Sang, W.B.; Matsuzawa, Y. Genome Mapping and Molecular Breeding in Plants: Vegetates; Springer: Berlin/Heidelberg, Germany, 2006; Chapter 3; pp. 141–155. [Google Scholar]
- Han, Q.; Li, J.; Shen, H.; Sakaguchi, S.; Isagi, Y.; Setoguchi, H. Genetic diversity and population structure of wild radish in East Asia. J. Anim. Plant Sci. 2022, 32, 1110–1119. [Google Scholar] [CrossRef]
- Manivannan, A.; Kim, J.H.; Kim, D.S.; Lee, E.S.; Lee, H.E. Deciphering the nutraceutical potential of Raphanus sativus—A comprehensive overview. Nutrients 2019, 11, 402. [Google Scholar] [CrossRef] [PubMed]
- Prakash, P.; Alien, M.R.M.; Poniesha, M.; Sai, N.R.; Masilamani, S.M.; Thirugnanasambandam, R.; Stanley, A.L. Effect of humic acid on seed germination of Raphanus sativus L. Int. J. Chemtech Res. 2014, 6, 4180–4185. [Google Scholar]
- Beevi, S.S.; Mangamoori, L.N.; Dhand, V.; Ramakrishna, D.S. Isothiocyanate profile and selective antibacterial activity of root, stem, and leaf extracts derived from Raphanus sativus L. Foodborne Pathog. Dis. 2009, 6, 129–136. [Google Scholar] [CrossRef]
- Rakhmawati, R.; Anggarwulan, E.; Retnaningtyas, E. Potency of Lobak leaves (Raphanus sativus L. var. Hortensis Back) as anticancer and antimicrobial candidates. Biodiversitas J. Biol. Divers. 2009, 10, 158–162. [Google Scholar] [CrossRef]
- Pocasap, P.; Weerapreeyakul, N.; Barusrux, S. Cancer preventive effect of Thai rat-tailed radish (Raphanus sativus L. var. caudatus Alef). J. Funct. Foods 2013, 5, 1372–1381. [Google Scholar] [CrossRef]
- Kim, S.; Woo, M.; Kim, M.; Noh, J.S.; Song, Y.O. Hot water extracts of pressure-roasted dried radish attenuates hepatic oxidative stress via Nrf2 up-regulation in mice fed high-fat diet. Food Sci. Biotechnol. 2017, 26, 1063–1069. [Google Scholar] [CrossRef]
- Baenas, N.; Piegholdt, S.; Schloesser, A.; Moreno, D.A.; García-Viguera, C.; Rimbach, G.; Wagner, A.E. Metabolic activity of radish sprouts derived isothiocyanates in drosophila melanogaster. Int. J. Mol. Sci. 2016, 17, 251. [Google Scholar] [CrossRef]
- Siddiq, A.; Younus, I. The Radish, Raphanus sativus L. Var. caudatus reduces anxiety-like behavior in mice. Metab. Brain Dis. 2018, 33, 1255–1260. [Google Scholar] [CrossRef]
- Do, M.H.; Kim, M.; Choi, S.Y.; Lee, P.; Kim, Y.; Hur, J. Wild radish (Raphanus sativus var. hortensis f. raphanistroides) root extract protects neuronal cells by inhibiting microglial activation. Appl. Biol. Chem. 2021, 64, 31. [Google Scholar] [CrossRef]
- Ishizuka, W.; Hikosaka, K.; Ito, M.; Morinaga, S.I. Temperature-related cline in the root mass fraction in East Asian wild radish along the Japanese archipelago. Breed. Sci. 2020, 70, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Austin, M. Species distribution models and ecological theory: A critical assessment and some possible new approaches. Ecol. Modell. 2007, 200, 1–19. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Modell. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Zhang, K.; Yao, L.; Meng, J.; Tao, J. Maxent modeling for predicting the potential geographical distribution of two peony species under climate change. Sci. Total Environ. 2018, 634, 1326–1334. [Google Scholar] [CrossRef]
- Qin, A.; Liu, B.; Guo, Q.; Bussmann, R.W.; Ma, F.; Jian, Z.; Xu, G.; Pei, S. Maxent modeling for predicting impacts of climate change on the potential distribution of Thuja sutchuenensis Franch., an extremely endangered conifer from southwestern China. Glob. Ecol. Conserv. 2017, 10, 139–146. [Google Scholar] [CrossRef]
- Li, J.; Fan, G.; He, Y. Predicting the current and future distribution of three Coptis herbs in China under climate change conditions, using the MaxEnt model and chemical analysis. Sci. Total Environ. 2020, 698, 134141. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Popp, A.; Calvin, K.; Fujimori, S.; Havlik, P.; Humpenöder, F.; Stehfest, E.; Bodirsky, B.L.; Dietrich, J.P.; Doelmann, J.C.; Gusti, M.; et al. Land-use futures in the shared socio-economic pathways. Glob. Environ. Change 2017, 42, 331–345. [Google Scholar] [CrossRef]
- Xin, X. Performance of BCC-CSM2-MR in simulating summer climate changes in East Asia. Geophys. Res. Abstr. 2019, 21, 1. [Google Scholar]
- Merow, C.; Smith, M.J.; Silander, J.A., Jr. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Vitasse, Y.; Signarbieux, C.; Fu, Y.H. Global warming leads to more uniform spring phenology across elevations. Proc. Natl. Acad. Sci. USA 2018, 115, 1004–1008. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Swets, J.A. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.Q.; Kushwaha, S.P.S.; Saran, S.; Xu, J.; Roy, P.S. Maxent modeling for predicting the potential distribution of medicinal plant, Justicia adhatoda L. in Lesser Himalayan foothills. Ecol. Eng. 2013, 51, 83–87. [Google Scholar] [CrossRef]
- Evans, J.M.; Fletcher, J.R.J.; Alavalapati, J. Using species distribution models to identify suitable areas for biofuel feedstock production. GCB Bioenergy 2010, 2, 63–78. [Google Scholar] [CrossRef]
- Saatchi, S.; Buermann, W.; Ter Steege, H.; Mori, S.; Smith, T.B. Modeling distribution of Amazonian tree species and diversity using remote sensing measurements. Remote Sens. Environ. 2008, 112, 2000–2017. [Google Scholar] [CrossRef]
- Yamagishi, H. Speciation and diversification of radish. In The Radish Genome; Nishio, T., Kitashiba, H., Eds.; Springer: Cham, Switzerland, 2017; pp. 11–30. [Google Scholar]
- Luo, L.Q.; Chen, Y.F.; Li, Q.Q.; Wang, W.Q.; Hou, X.L. New distribution records of one genus and six Plants from coastal zones, Fujian Province. J. Fujian Forestry Sci. Technol. 2017, 44, 82–86, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Li, H.R.; Wang, Y.; Yan, X.L.; Wang, Z.H.; Yan, J.; Ma, J.S. New data of Shanghai flora. J. East China Norm. Univ. Nat. Sci. 2017, 1, 132–138, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Ma, K. Flora of Wenzhou, a practical demo for local flora in China. Biodiv. Sci. 2017, 25, 1350–1351, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Stagnari, F.; Galieni, A.; D’Egidio, S.; Pagnani, G.; Pisante, M. Responses of radish (Raphanus sativus) to drought stress. Ann. Appl. Biol. 2018, 172, 170–186. [Google Scholar] [CrossRef]
- Eslami, S.V.; Gill, G.S.; McDonald, G. Effect of water stress during seed development on morphometric characteristics and dormancy of wild radish (Raphanus raphanistrum L.) seeds. Int. J. Plant Prod. 2010, 4, 159–168. [Google Scholar]
- Huang, Z.; Xie, L.; Wang, H.; Zhong, J.; Li, Y.; Liu, J.; Ou, Z.; Liang, X.; Li, Y.; Huang, H.; et al. Geographic distribution and impacts of climate change on the suitable habitats of Zingiber species in China. Ind. Crop. Prod. 2019, 138, 111429. [Google Scholar] [CrossRef]
- Han, Q.; Higashi, H.; Mitsui, Y.; Setoguchi, H. Lineage isolation in the face of active gene flow in the coastal plant wild radish is reinforced by differentiated vernalisation responses. BMC Evol. Biol. 2016, 16, 84. [Google Scholar] [CrossRef] [PubMed]
- Blackshaw, R.E.; Lemerle, D.; Mailer, R.; Young, K.R. Influence of wild radish on yield and quality of canola. Weed Sci. 2002, 50, 344–349. [Google Scholar] [CrossRef]
- Kang, E.S.; Ha, S.M.; Ko, H.C.; Yu, H.-J.; Chae, W.B. Reproductive traits and molecular evidence related to the global distribution of cultivated radish (Raphanus sativus L.). Plant Syst. Evol. 2016, 302, 1367–1380. [Google Scholar] [CrossRef]
- Engelen-Eigles, G.; Erwin, J.E. A model plant for vernalization studies. Sci. Hortic. 1997, 70, 197–202. [Google Scholar] [CrossRef]
- Han, Q.; Sakaguchi, S.; Wakabayashi, T.; Setoguchi, H. Association between RsFT, RsFLC and RsCOL5 (A&B) expression and flowering regulation in Japanese wild radish. AoB Plants 2021, 13, plab039. [Google Scholar] [CrossRef] [PubMed]
- Jiang, K.; Tong, X.; Ding, Y.Q.; Wang, Z.W.; Miao, L.Y.; Xiao, Y.E.; Huang, W.C.; Hu, Y.H.; Chen, X.Y. Shifting roles of the East China Sea in the phylogeography of red nanmu in East Asia. J. Biogeogr. 2021, 48, 2486–2501. [Google Scholar] [CrossRef]
- Qi, X.S.; Chen, C.; Comes, H.P.; Sakaguchi, S.; Liu, Y.H.; Tanaka, N.; Sakio, H.; Qiu, Y.X. Molecular data and ecological niche modelling reveal a highly dynamic evolutionary history of the East Asian Tertiary relict Cercidiphyllum (Cercidiphyllaceae). New Phytol. 2012, 196, 617–630. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Sun, J. Predicting the distribution of Stipa purpurea across the Tibetan Plateau via the MaxEnt model. BMC Ecol. 2018, 18, 10. [Google Scholar] [CrossRef]
- Khalil, T.; Asad, S.A.; Khubaib, N.; Baig, A.; Atif, S.; Umar, M.; Kropp, J.P.; Pradhan, P.; Baig, S. Climate change and potential distribution of potato (Solanum tuberosum) crop cultivation in Pakistan using Maxent. AIMS Agric. Food 2021, 6, 663–676. [Google Scholar] [CrossRef]
- Aguirre-Liguori, J.A.; Ramírez-Barahona, S.; Tiffin, P.; Eguiarte, L.E. Climate change is predicted to disrupt patterns of local adaptation in wild and cultivated maize. Proc. R. Soc. B 2019, 286, 20190486. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, R.; Lenoir, J.; Piedallu, C.; Riofrío-Dillon, G.; De Ruffray, P.; Vidal, C.; Pierrat, J.C.; Gégout, J.C. Changes in plant community composition lag behind climate warming in lowland forests. Nature 2011, 479, 517–520. [Google Scholar] [CrossRef] [PubMed]
- Lukac, M.; Calfapietra, C.; Lagomarsino, A.; Loreto, F. Global climate change and tree nutrition: Effects of elevated CO2 and temperature. Tree Physiol. 2010, 30, 1209–1220. [Google Scholar] [CrossRef] [PubMed]
- Bhargava, S.; Mitra, S. Elevated atmospheric CO2 and the future of crop plants. Plant Breed. 2021, 140, 1–11. [Google Scholar] [CrossRef]
- Gamage, D.; Thompson, M.; Sutherland, M.; Hirotsu, N.; Makino, A.; Seneweera, S. New insights into the cellular mechanisms of plant growth at elevated atmospheric carbon dioxide concentrations. Plant Cell Environ. 2018, 41, 1233–1246. [Google Scholar] [CrossRef] [PubMed]
- Osborne, C.P. Crop yields: CO2 fertilization dries up. Nat. Plants 2016, 2, 16138. [Google Scholar] [CrossRef] [PubMed]
- Makowski, D.; Marajo-Petitzon, E.; Durand, J.L.; Ben-Ari, T. Quantitative synthesis of temperature, CO2, rainfall, and adaptation effects on global crop yields. Eur. J. Agron. 2020, 115, 126041. [Google Scholar] [CrossRef]
- Dusenge, M.E.; Duarte, A.G.; Way, D.A. Plant carbon metabolism and climate change: Elevated CO2 and temperature impacts on photosynthesis, photorespiration and respiration. New Phytol. 2019, 221, 32–49. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Description | Units |
|---|---|---|
| bio01 | Annual Mean Temperature | °C |
| bio02 | Mean Diurnal Range (Mean of the monthly (maximum–minimum temperatures)) | °C |
| bio03 | Isothermality (bio02/bio07) (×100) | - |
| bio04 | Temperature Seasonality (Standard Deviation ×100) | - |
| bio05 | Maximum Temperature of the Warmest Month | °C |
| bio06 | Minimum Temperature of the Coldest Month | °C |
| bio07 | Temperature Annual Range (bio05-bio06) | °C |
| bio08 | Mean Temperature of the Wettest Quarter | °C |
| bio09 | Mean Temperature of the Driest Quarter | °C |
| bio10 | Mean Temperature of the Warmest Quarter | °C |
| bio11 | Mean Temperature of the Coldest Quarter | °C |
| bio12 | Annual Precipitation | mm |
| bio13 | Precipitation of the Wettest Month | mm |
| bio14 | Precipitation of the Driest Month | mm |
| bio15 | Precipitation Seasonality (Coefficient of Variation) | - |
| bio16 | Precipitation of the Wettest Quarter | mm |
| bio17 | Precipitation of the Driest Quarter | mm |
| bio18 | Precipitation of the Warmest Quarter | mm |
| bio19 | Precipitation of the Coldest Quarter | mm |
| Time Period | Area of Each Suitable Habitat (The Change in Area Compared with the Current Period) 104 km2 | |||
|---|---|---|---|---|
| Lowly Suitable Habitat | Moderately Suitable Habitat | Highly Suitable Habitat | Total Suitable Habitat | |
| LGM | 83.0800 (−9.3200) | 35.4804 (−19.7044) | 42.2017 (−29.1812) | 160.763 (−58.2055) |
| Current | 73.7610 (0.0000) | 15.7760 (0.0000) | 13.0204 (0.0000) | 102.5574 (0.0000) |
| 2070 (SSP1-2.6) | 67.9276 (−5.8334) | 18.8107 (3.0348) | 13.0591 (0.0387) | 99.7975 (−2.7599) |
| 2070 (SSP5-8.5) | 84.2481 (10.4871) | 28.8238 (13.0479) | 11.8967 (−1.1238) | 124.9686 (22.4112) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, Q.; Liu, Y.; Jiang, H.; Chen, X.; Feng, H. Evaluation of Climate Change Impacts on the Potential Distribution of Wild Radish in East Asia. Plants 2023, 12, 3187. https://doi.org/10.3390/plants12183187
Han Q, Liu Y, Jiang H, Chen X, Feng H. Evaluation of Climate Change Impacts on the Potential Distribution of Wild Radish in East Asia. Plants. 2023; 12(18):3187. https://doi.org/10.3390/plants12183187
Chicago/Turabian StyleHan, Qingxiang, Ye Liu, Hongsheng Jiang, Xietian Chen, and Huizhe Feng. 2023. "Evaluation of Climate Change Impacts on the Potential Distribution of Wild Radish in East Asia" Plants 12, no. 18: 3187. https://doi.org/10.3390/plants12183187
APA StyleHan, Q., Liu, Y., Jiang, H., Chen, X., & Feng, H. (2023). Evaluation of Climate Change Impacts on the Potential Distribution of Wild Radish in East Asia. Plants, 12(18), 3187. https://doi.org/10.3390/plants12183187