

In Vitro Polyploidization of Brassolaeliocattleya Hybrid Orchid

 , and
, and
Abstract
:1. Introduction
2. Results
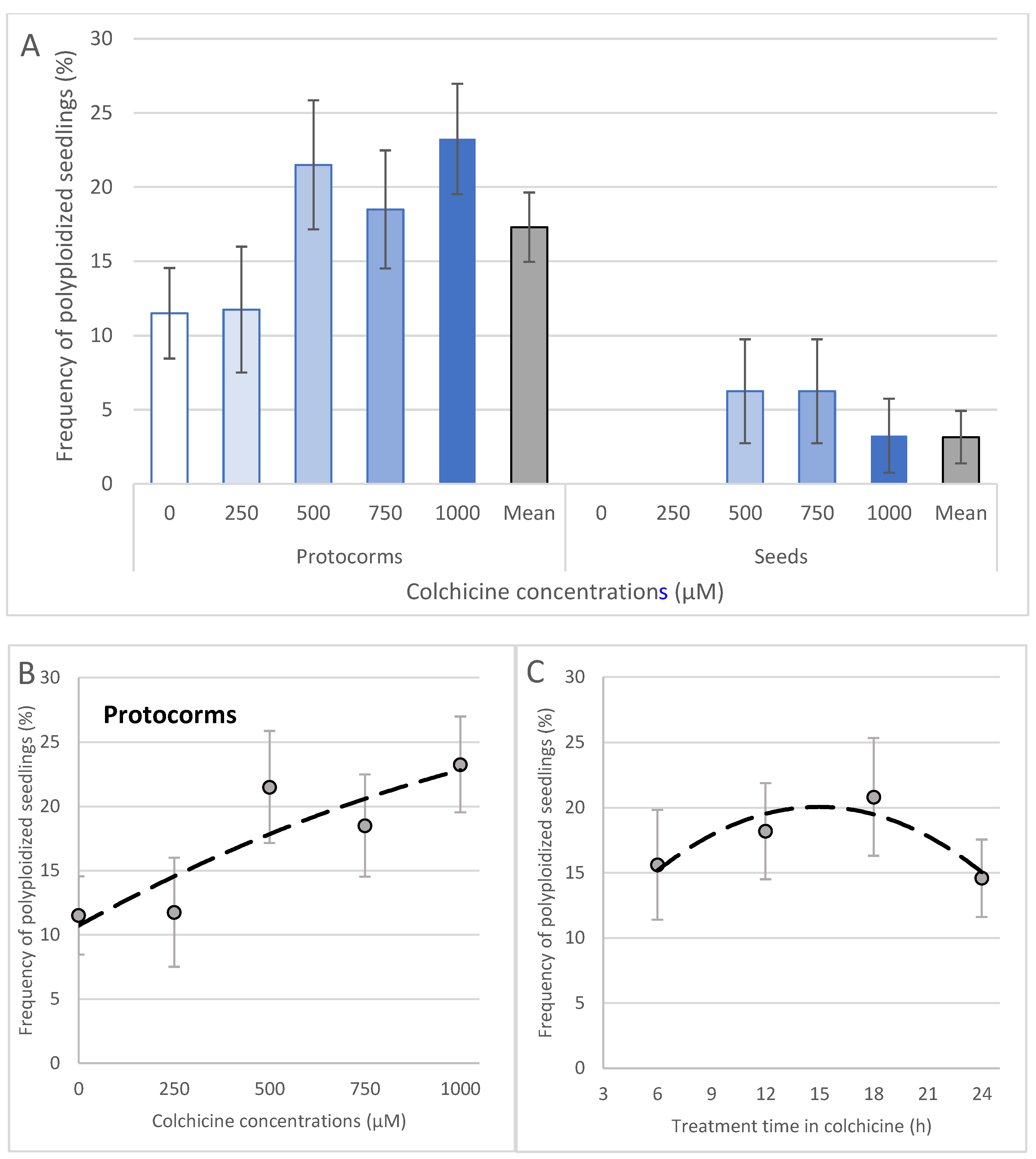
2.1. Colchicine Effects on Explant Development
- (a)
- Seeds
- (b)
- Protocorms
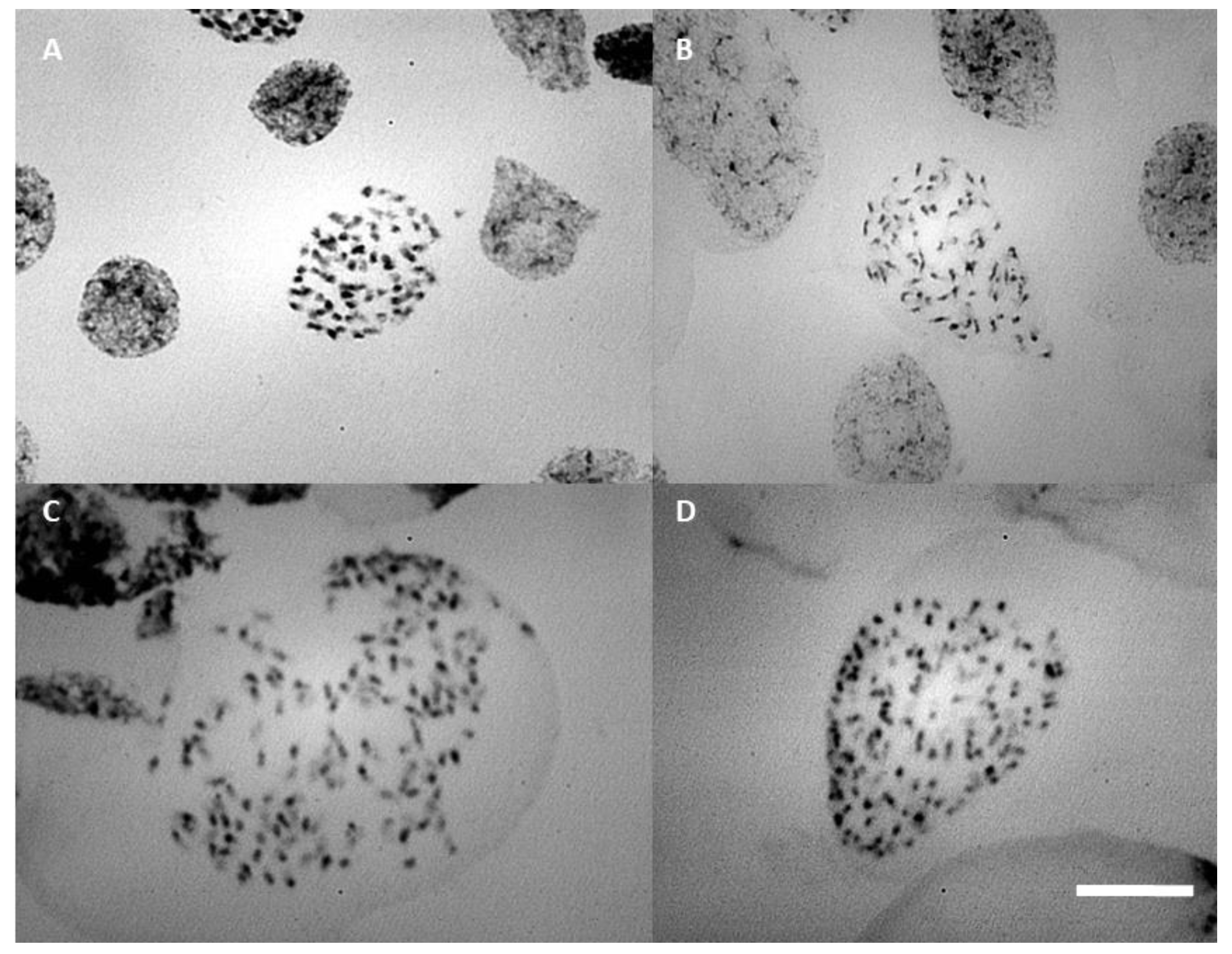
2.2. Flow Cytometry and Cytogenetic Analysis
2.3. Protocorms Resulted in the Best Polyploidization Rates Compared to Seeds
3. Discussion
3.1. Colchicine Switched the In Vitro Development of Blc. Orchid
3.2. Colchicine Is Efficient at Inducing Polyploidization in Blc. Orchid
3.3. Ploidy Levels and Chromosome Counting in Blc. Orchids
4. Material and Methods
4.1. Plant Material, In Vitro Establishment and Growth
4.2. Colchicine Treatment Procedures
4.3. Effects of Colchicine on Seeds and Protocorms
- (a)
- Seeds: Using the counting method, the percentage of seeds was evaluated by the presence or absence of the embryo inside the testa with the aid of an optical microscope (Nikon Eclipse e200, Nikon Instruments, Japan) with a 4X objective lens (Figure 1B). This included the percentage of germination 180 days after seeding; the percentage of embryos that developed into protocorms and/or seedlings based on fresh mass calculation of each type of development after germination; the total fresh weight of plant tissue obtained per vial.
- (b)
- Protocorms: After 180 days of culture following the treatment with colchicine, the percentages of survival and death of protocorms, the percentages of PLBs proliferation and the regeneration into plantlets and the total fresh weight obtained per vial were recorded.
4.4. Flow Cytometry Analysis
4.5. Chromosome Counting
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- AOS. The Showy Cattleya, Queen of the Orchids. 2013. Available online: http://www.aos.org/Default.aspx?id=%0A443 (accessed on 16 May 2022).
- Cavero, M.; Collantes, B.; Patroni, C. Orquídeas Del Peru; Centro de Datos para la Conservación del Perú: La Molina, Peru, 1991. [Google Scholar]
- Cardoso, J.C.; Martinelli, A.P.; da Silva, J.A.T. A novel approach for the selection of Cattleya hybrids for precocious and season-independent flowering. Euphytica 2016, 210, 143–150. [Google Scholar] [CrossRef]
- Takane, R.J.; Yanagisawa, S.S.; Pivetta, K.F.L. Cultivo Moderno de Orquídeas: Cattleya e Seus Híbridos; UFC: Fortaleza, Brazil, 2010. [Google Scholar]
- Cardoso, J.C.; Ono, E.O.; Rodrigues, J.D. Gibberellic acid and water regime in the flowering induction of Brassocattleya and Cattleya hybrid orchids. Hortic. Bras. 2010, 28, 395–398. [Google Scholar] [CrossRef] [Green Version]
- Vilcherrez-Atoche, J.A.; Iiyama, C.M.; Cardoso, J.C. Polyploidization in Orchids: From Cellular Changes to Breeding Applications. Plants 2022, 11, 469. [Google Scholar] [CrossRef]
- Soltis, P.S.; Marchant, D.B.; Van de Peer, Y.; Soltis, D.E. Polyploidy and Genome Evolution in Plants. Curr. Opin. Genet. Dev. 2015, 35, 119–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jolanta, M.; Kolano, B.; Sas-Nowosielska, H. Endopolyploidy in Plants. In Plant Genome Diversity; Springer: Vienna, Brazil, 2013; Volume 2, pp. 99–119. [Google Scholar]
- Ranney, T.G. Polyploidy: From Evolution to New Plant Development. Comb. Proc. Int. Plant Propagators’ Soc. 2006, 56, 137–142. [Google Scholar]
- Dhooghe, E.; Van Laere, K.; Eeckhaut, T.; Leus, L.; Van Huylenbroeck, J. Mitotic chromosome doubling of plant tissues in vitro. Plant Cell Tissue Organ Cult. (PCTOC) 2011, 104, 359–373. [Google Scholar] [CrossRef]
- Mondin, M.; Neto, A.D. Citogenética Vegetal Enfatizando a Família Orchidaceae. Orchidstudium 2006, 4, 24–54. [Google Scholar]
- Manzoor, A.; Ahmad, T.; Bashir, M.A.; Hafiz, I.A.; Silvestri, C. Studies on Colchicine Induced Chromosome Doubling for Enhancement of Quality Traits in Ornamental Plants. Plants 2019, 8, 194. [Google Scholar] [CrossRef] [Green Version]
- Chung, M.Y.; Kim, C.Y.; Min, J.S.; Lee, D.-J.; Naing, A.H.; Chung, J.D.; Kim, C.K. In Vitro Induction of Tetraploids in an Interspecific Hybrid of Calanthe (Calanthe Discolor× Calanthe Sieboldii) through Colchicine and Oryzalin Treatments. Plant Biotechnol. Rep. 2014, 8, 251–257. [Google Scholar] [CrossRef]
- Kobayashi, N.; Yamashita, S.; Ohta, K.; Hosoki, T. Morphological Characteristics and Their Inheritance in Colchicine-induced Salvia Polyploids. J. Jpn. Soc. Hortic. Sci. 2008, 77, 186–191. [Google Scholar] [CrossRef] [Green Version]
- Manzoor, A.; Ahmad, T.; Bashir, M.A.; Baig, M.M.Q.; Quresh, A.A.; Shah, M.K.N.; Hafiz, I.A. Induction and Identification of Colchicine Induced Polyploidy in Gladiolus Grandiflorus ‘White Prosperity’. Folia Hortic. 2018, 30, 307–319. [Google Scholar] [CrossRef] [Green Version]
- Kushwah, K.S.; Verma, R.C.; Patel, S.; Jain, N. Colchicine Induced Polyploidy in Chrysanthemum carinatum L. J. Phylogenet. Evol. Biol. 2018, 6, 2. [Google Scholar] [CrossRef]
- Huy, N.P.; Tam, D.T.T.; Luan, V.Q.; Tung, H.T.; Hien, V.T.; Ngan, H.T.M.; Duy, P.N.; Nhut, D.T. In vitro polyploid induction of Paphiopedilum villosum using colchicine. Sci. Hortic. 2019, 252, 283–290. [Google Scholar] [CrossRef]
- Chen, W.-H.; Tang, C.-Y. A Protocol for the Induction of Polyploids in Phalaenopsis Orchids by In Vitro Method Without Using Anti-Microtubule Agents. In Orchid Propagation: From Laboratories to Greenhouses—Methods and Protocols; Lee, Y.I., Yeung, E.T., Eds.; Humana Press: New York, NY, USA, 2018; pp. 317–330. [Google Scholar]
- Ji, B.X.; Chen, D.W.; Zhang, C.C.; Min, D.; Huang, W.J.; Wang, Y. High Efficient Polyploid Induction of Cymbidium Hybridium. Bull. Bot. Res. 2011, 31, 558–562. [Google Scholar]
- Vichiato, M.R.M.; Vichiato, M.; Pasqual, M.; Castro, D.M.; Dutra, L.F. Indução e Identificação de Tetraplóides Em Dendrobium Nobile Lindl.(Orchidaceae). Rev. Ciência Agronômica 2007, 38, 385–390. [Google Scholar]
- Cui, G.R.; Zhang, Z.X.; Zhang, C.Y.; Hu, N.B.; Sui, Y.H.; Li, J.Q. Polyploid Induction and Identification of Oncidium. Acta Prataculturae Sin. 2010, 19, 184–190. [Google Scholar]
- Menezes-Sá, T.S.A.; de Fátima Arrigoni-Blank, M.; da Costa, A.S.; de Almeida Santos-Serejo, J.; Blank, A.F.; Soares, C.A.; Moura, G.M.S. Chromosome Doubling in Cattleya Tigrina, A. Rich. Sci. Plena 2019, 15, 11. [Google Scholar] [CrossRef] [Green Version]
- Silva, P.A.K.X.; Callegari-Jacques, S.; Bodanese-Zanettini, M.H. Induction and Identification of Polyploids in Cattleya Intermedia Lindl.(Orchidaceae) by in Vitro Techniques. Ciência Rural 2000, 30, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Wibisono, K.; Syarifah, I.A.; Waras, N.; Suhest, S. Sensitivity in Callus Tissue of Plectranthus Amboinicus (L.) through Mutation Induction with Colchicine. AGRIVITA J. Agric. Sci. 2021, 44, 82–95. [Google Scholar] [CrossRef]
- Li, M.; Ding, B.; Huang, W.; Pan, J.; Ding, Z.; Jiang, F. Induction and Characterization of Tetraploids from Seeds of Bletilla Striata (Thunb.) Reichb. F. BioMed Res. Int. 2018, 2018, 3246398. [Google Scholar]
- Lone, A.B.; Unemoto, L.K.; Takahashi, L.S.A.; Faria, R.T. Sobrevivência e desenvolvimento in vitro de Cattleya (Orchidaceae) submetida a tratamentos com colchicina. Semin. Ciências Agrárias 2010, 31, 1337. [Google Scholar] [CrossRef] [Green Version]
- Mustika, T.; Indrianto, A. Improvement of Orchid Vanda Hybrid (Vanda Limbata Blume x Vanda Tricolor Lindl. Var. Suavis) by Colchicines Treatment in Vitro. Modern. Appl. Sci. 2016, 10, 83–89. [Google Scholar]
- Thao, N.T.P.; Ureshino, K.; Miyajima, I.; Ozaki, Y.; Okubo, H. Induction of tetraploids in ornamental Alocasia through colchicine and oryzalin treatments. Plant Cell Tissue Organ Cult. (PCTOC) 2003, 72, 19–25. [Google Scholar] [CrossRef]
- Heo, J.-Y.; Jeong, S.-H.; Choi, H.-R.; Park, S.-M. Polyploid Production in Lilium Leichtlinii Var. Maximowiczii Using Colchicine. JAPS J. Anim. Plant Sci. 2016, 26. [Google Scholar]
- Song, L.; Yang, J.; Liu, D.; Li, Z.; Wang, Y. Polyploid Induction in Cymbidium Sinenthese’Lv Mosu’× Cymbidium Hybridum’Shijieheping’F1 Generation. Guangxi Zhiwu/Guihaia 2018, 38, 188–194. [Google Scholar]
- Atichart, P. Polyploid Induction by Colchicine Treatments and Plant Regeneration of Dendrobium Chrysotoxum. Thai J. Agric. Sci. 2013, 46, 59–63. [Google Scholar]
- Griesbach, R.J. Colchicine-induced polyploidy in phalaenopsis orchids. Plant Cell Tissue Organ Cult. (PCTOC) 1981, 1, 103–107. [Google Scholar] [CrossRef]
- De, L.C.; Rao, A.N.; Rajeevan, P.K.; Pathak, P. ORCHID IMPROVEMENT-AN OVERVIEW. J. Orchid. Soc. India 2014, 28, 35–45. [Google Scholar]
- Chen, W.H.; Tang, C.Y.; Kao, Y.L. Ploidy doubling by in vitro culture of excised protocorms or protocorm-like bodies in Phalaenopsis species. Plant Cell Tissue Organ Cult. (PCTOC) 2009, 98, 229–238. [Google Scholar] [CrossRef]
- Souza, B.C.Q.; Souza, A.C.M.; Almeida, E.M.; Felix, L.P. Orchidaceae-IAPT/IOPB Chromosome Data 26. Taxon 2017, 66, 1487–1499. [Google Scholar]
- Eng, W.-H.; Ho, W.-S. Polyploidization Using Colchicine in Horticultural Plants: A Review. Sci. Hortic. 2019, 246, 604–617. [Google Scholar] [CrossRef]
- Grosso, V.; Farina, A.; Giorgi, D.; Nardi, L.; Diretto, G.; Lucretti, S. A high-throughput flow cytometry system for early screening of in vitro made polyploids in Dendrobium hybrids. Plant Cell Tissue Organ Cult. (PCTOC) 2018, 132, 57–70. [Google Scholar] [CrossRef]
- Yenchon, S.; Te-Chato, S. Polyploidy Induction of Dendrobium Formosum by Colchicine Treatment in Vitro. Acta Hortic. 2014, 1025, 81–88. [Google Scholar] [CrossRef]
- Xuejiao, L.; Zhilin, L.; Lipin, H. Induction and Identification of Polyploids in Wild Cymbidium Lowianum. Chin. Agric. Sci. Bull. 2010, 13, 60. [Google Scholar]
- Rahayu, E.M.D.; Sukma, D.; Syukur, M.; Irawati, D. Polyploid Induction of Phalaenopsis Amabilis (L.) Blume and Phalaenopsis Amboinensis, J. J. Smith by Colchicine in In Vitro Culture. Indones. J. Agron. 2015, 43, 219–226. [Google Scholar] [CrossRef] [Green Version]
- IOF. Orchid Hybrid: Brassolaeliocattleya Goldenzelle. ORCHIDS.ORG. 2022a. Available online: https://www.orchids.org/grexes/brassolaeliocattleya-goldenzelle (accessed on 25 January 2022).
- IOF. Orchid Hybrid: Brassolaeliocattleya Haw Yuan Beauty. ORCHIDS.ORG. 2022b. Available online: https://www.orchids.org/grexes/brassolaeliocattleya-haw-yuan-beauty (accessed on 25 January 2022).
- Querino, B.C.; Ferraz, M.E.; Mata-Sucre, Y.; Souza, G.; Felix, L.P. Cytomolecular diversity of the subtribe Laeliinae (Epidendroidae, Orchidaceae) suggests no relationship between genome size and heterochromatin abundance. Plant Syst. Evol. 2020, 306, 19. [Google Scholar] [CrossRef]
- Souza, B.C.Q. Citogenética Da Subtribo Laeliinae (Orchidaceae: Epidendroideae): Regiões Heterocromáticas e Localização Do DNA Ribossomal. In Dissertação de Mestrado; Universidade Federal da Paraıba: Paraıba, Brazil, 2011. [Google Scholar]
- Felix, L.P.; Guerra, M. Variation in chromosome number and the basic number of subfamily Epidendroideae (Orchidaceae). Bot. J. Linn. Soc. 2010, 163, 234–278. [Google Scholar] [CrossRef]
- Moraes, A.P.; Chinaglia, M.; Palma-Silva, C.; Pinheiro, F. Interploidy Hybridization in Sympatric Zones: The Formation of E Pidendrum Fulgens× E. Puniceoluteum Hybrids (E Pidendroideae, O Rchidaceae). Ecol. Evol. 2013, 3, 3824–3837. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bioassays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Galbraith, D.W.; Harkins, K.R.; Maddox, J.M.; Ayres, N.M.; Sharma, D.P.; Firoozabady, E. Rapid Flow Cytometric Analysis of the Cell Cycle in Intact Plant. Tissues. Science 1983, 220, 1049–1051. [Google Scholar] [CrossRef]
- Otto, F. Chapter 11 DAPI Staining of Fixed Cells for High-Resolution Flow Cytometry of Nuclear DNA. In Methods in Cell Biology; Academic Press: Cambridge, MA, USA, 1990; Volume 33, pp. 105–110. [Google Scholar] [CrossRef]
- Praça-Fontes, M.M.; Carvalho, C.R.; Clarindo, W.R.; Cruz, C.D. Revisiting the DNA C-values of the genome size-standards used in plant flow cytometry to choose the “best primary standards”. Plant Cell Rep. 2011, 30, 1183–1191. [Google Scholar] [CrossRef] [PubMed]
- Mondin, M.; Aguiar-Perecin, M.L.R. Coloração Pelo Método de Feulgen. 2009. Available online: http://www.genetica.esalq.usp.br/citogenetica/protocolos/Feulgen.pdf (accessed on 10 October 2022).










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Seeds | ||||
| Colchicine Treatment Time (h) | GP (%) | PrD (%) | SeD (%) | TFW (g) |
| 6 | 44.86 a | 54.99 c | 45.01 a | 13.62 a |
| 12 | 31.53 c | 62.51 ab | 39.15 ab | 12.40 a |
| 18 | 29.94 c | 58.25 bc | 41.75 a | 12.26 a |
| 24 | 36.96 b | 68.36 a | 31.64 b | 13.42 a |
| Colchicine concentrations (μM) | ||||
| 0.00 | 49.42 a | 62.88 a | 37.10 b | 14.52 a |
| 250 | 36.21 b | 60.38 ab | 39.62 ab | 12.76 ab |
| 500 | 34.86 b | 67.21 a | 34.86 b | 13.08 ab |
| 750 | 30.64 c | 61.75 a | 38.25 b | 12.50 ab |
| 1000 | 27.97 d | 52.91 b | 47.09 a | 11.77 b |
| F Colchicine treatment time (h) | 59.15 ** | 12.91 ** | 10.06 ** | 2.49 ns |
| F Colchicine concentrations (%) | 69.89 ** | 7.36 ** | 4.78 ** | 3.85 ** |
| F Interaction | 5.13 ** | 4.1 ** | 3.10 ** | 1.64 ns |
| CV (%) | 3.28 | 24.19 | 4.05 | 5.96 |
| Protocorm | ||||
| Colchicine Treatment Time (h) | PS (%) | PLBP (%) | RIP (%) | TFW (g) |
| 6 | 64.67 a | 59.38 a | 40.62 b | 10.07 ab |
| 12 | 59.50 a | 36.94 b | 63.06 a | 11.70 ab |
| 18 | 63.34 a | 46.91 ab | 53.09 ab | 8.92 b |
| 24 | 61.67 a | 40.92 b | 59.08 a | 12.30 a |
| Colchicine concentrations (μM) | ||||
| 0.00 | 72.92 a | 44.19 a | 55.80 a | 12.75 a |
| 250 | 71.25 a | 47.21 a | 52.79 a | 11.01 ab |
| 500 | 66.67 ab | 43.47 a | 56.53 a | 11.23 ab |
| 750 | 51.04 bc | 46.33 a | 53.67 a | 10.07 ab |
| 1000 | 49.58 c | 48.97 a | 51.03 a | 8.66 b |
| F treatment time | 0.49 ns | 5.0 ** | 5.25 ** | 4.07 * |
| F concentrations | 7.66 ** | 0.21 ns | 0.22 ns | 2.30 * |
| F Interaction | 1.92 * | 0.67 ns | 0.70 ns | 0.96 ns |
| CV (%) | 15.97 | 41.11 | 33.36 | 22.43 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vilcherrez-Atoche, J.A.; Silva, J.C.; Clarindo, W.R.; Mondin, M.; Cardoso, J.C. In Vitro Polyploidization of Brassolaeliocattleya Hybrid Orchid. Plants 2023, 12, 281. https://doi.org/10.3390/plants12020281
Vilcherrez-Atoche JA, Silva JC, Clarindo WR, Mondin M, Cardoso JC. In Vitro Polyploidization of Brassolaeliocattleya Hybrid Orchid. Plants. 2023; 12(2):281. https://doi.org/10.3390/plants12020281
Chicago/Turabian StyleVilcherrez-Atoche, Joe Abdul, Jéssica Coutinho Silva, Wellington Ronildo Clarindo, Mateus Mondin, and Jean Carlos Cardoso. 2023. "In Vitro Polyploidization of Brassolaeliocattleya Hybrid Orchid" Plants 12, no. 2: 281. https://doi.org/10.3390/plants12020281
APA StyleVilcherrez-Atoche, J. A., Silva, J. C., Clarindo, W. R., Mondin, M., & Cardoso, J. C. (2023). In Vitro Polyploidization of Brassolaeliocattleya Hybrid Orchid. Plants, 12(2), 281. https://doi.org/10.3390/plants12020281