

FKF1 Interacts with CHUP1 and Regulates Chloroplast Movement in Arabidopsis
 and
and 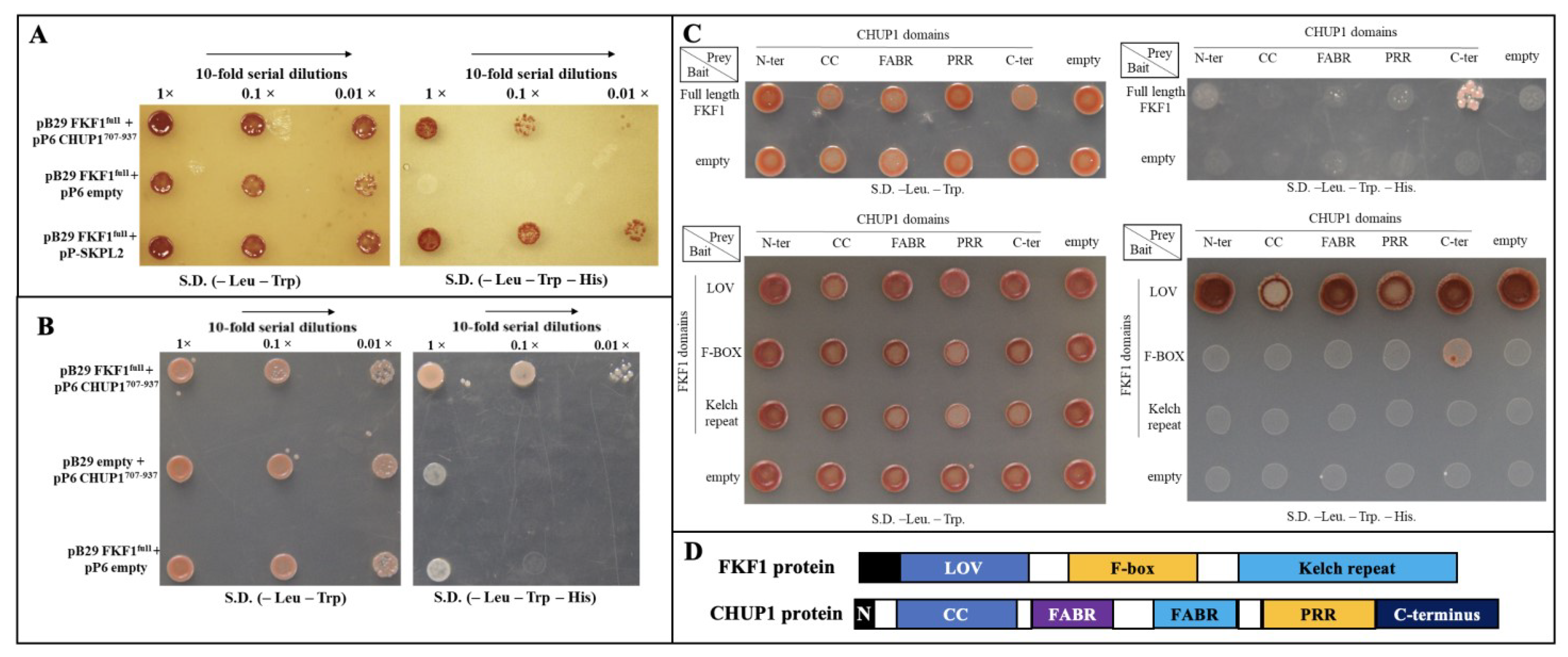{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. FKF1 Physically Interacts with CHUP1
2.2. F-Box Domain of FKF1 Interacts with the C-Terminal Domain of CHUP1
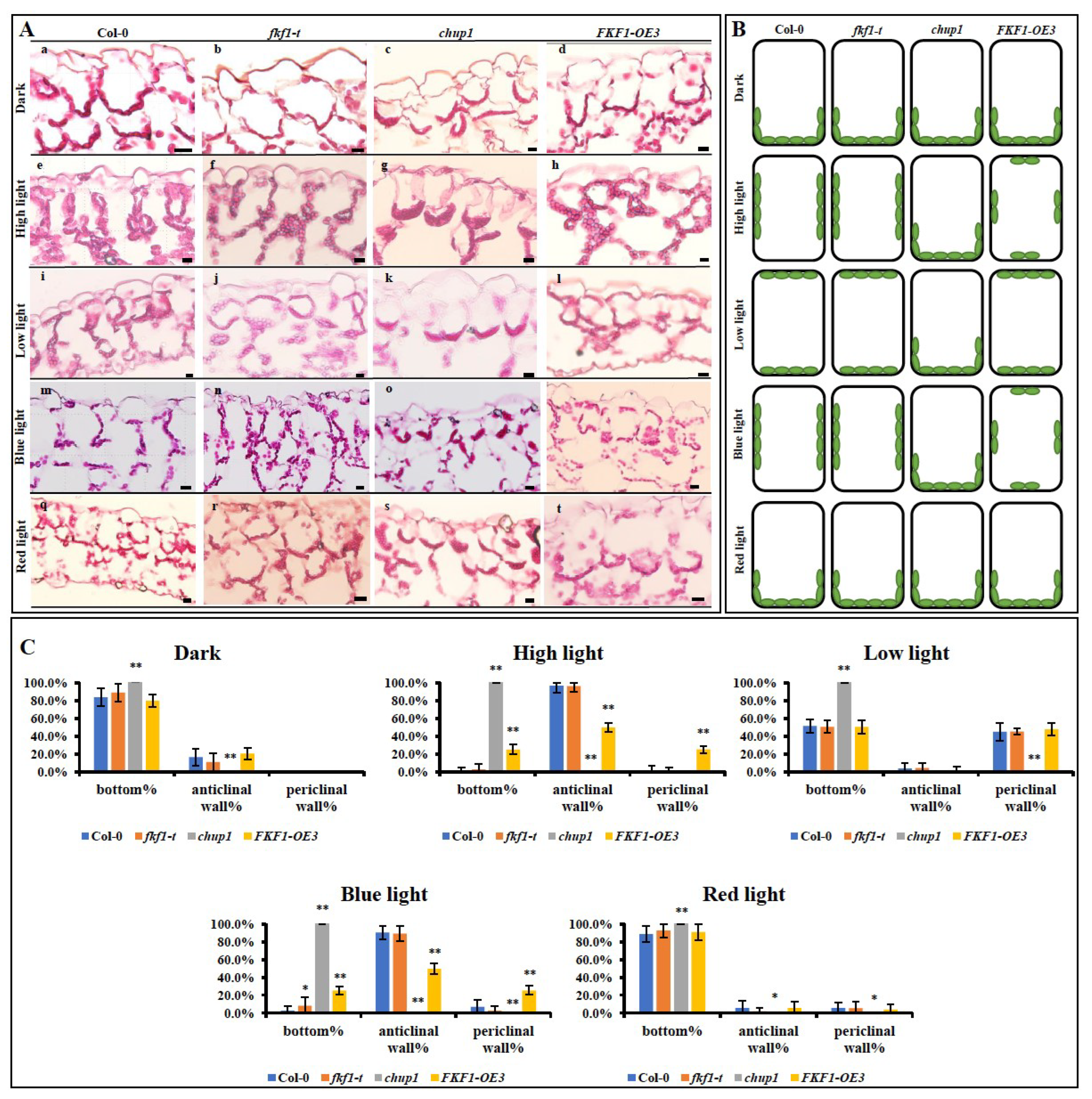
2.3. Overexpression of FKF1 Alters the Chloroplast Localization within the Cells
2.4. The Chloroplast Avoidance Response to Light Is Impaired in FKF1 Overexpression Lines
2.5. FKF1 Regulates CHUP1 Protein Abundance
2.6. FKF1 Alters the Expressions of Chloroplast Photorelocation Pathway Genes
2.7. Altering the Expression Levels of THRUMIN1 and JAC1 Did Not Rescue the Impaired Avoidance Response of FKF1-OE Plant
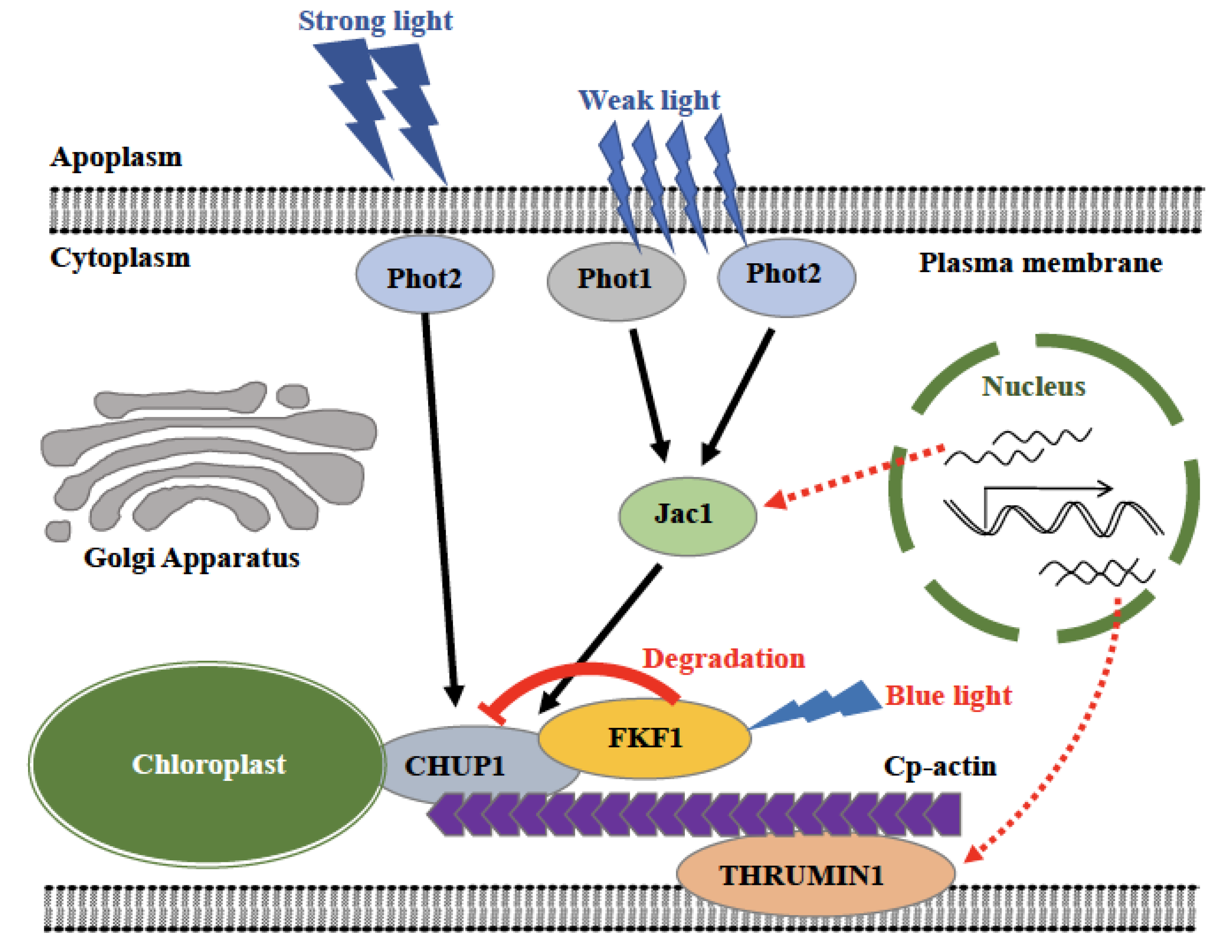
3. Discussion
4. Materials and Methods
4.1. Plant Growth and Light Conditions
4.2. Quantitative Analysis of Gene Expression
4.3. Plasmid Construction and Arabidopsis Transformation
4.4. Genome-Wide Yeast Two-Hybrid Screening to Identify FKF1 Interacting Proteins
4.5. Confirmation of FKF1 and CHUP1 Interaction
4.6. Bimolecular Fluorescence Complementation (BiFC)
4.7. Red-Light Transmittance Studies
4.8. Histochemical Analysis of Chloroplasts Positioning
4.9. Western Blot Analysis
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yuan, N.; Balasubramanian, V.K.; Chopra, R.; Mendu, V. The Photoperiodic Flowering Time Regulator FKF1 Negatively Regulates Cellulose Biosynthesis. Plant Physiol. 2019, 180, 2240–2253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawa, M.; Nusinow, D.A.; Kay, S.A.; Imaizumi, T. FKF1 and GIGANTEA complex formation is required for day-length measurement in Arabidopsis. Science 2007, 318, 261–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, D.C.; Lasswell, J.; Rogg, L.E.; Cohen, M.A.; Bartel, B. FKF1, a clock-controlled gene that regulates the transition to flowering in Arabidopsis. Cell 2000, 101, 331–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nawkar, G.M.; Maibam, P.; Park, J.H.; Sahi, V.P.; Lee, S.Y.; Kang, C.H. UV-Induced cell death in plants. Int. J. Mol. Sci. 2013, 14, 1608–1628. [Google Scholar] [CrossRef] [Green Version]
- Kasahara, M.; Kagawa, T.; Oikawa, K.; Suetsugu, N.; Miyao, M.; Wada, M. Chloroplast avoidance movement reduces photodamage in plants. Nature 2002, 420, 829–832. [Google Scholar] [CrossRef] [PubMed]
- Suetsugu, N.; Wada, M. Chloroplast Photorelocation Movement: A Sophisticated Strategy for Chloroplasts to Perform Efficient Photosynthesis. In Advances in Photosynthesis-Fundamental Aspects; Intechopen: London, UK, 2012; pp. 215–234. [Google Scholar] [CrossRef]
- Wada, M. Chloroplast movement. Plant Sci. 2013, 210, 177–182. [Google Scholar] [CrossRef]
- Lin, C. Blue Light Receptors and Signal Transduction. Plant Cell 2002, 14, S207–S225. [Google Scholar] [CrossRef] [Green Version]
- Sakai, T.; Kagawa, T.; Kasahara, M.; Swartz, T.E.; Christie, J.M.; Briggs, W.R.; Wada, M.; Okada, K. Arabidopsis nph1 and npl1: Blue light receptors that mediate both phototropism and chloroplast relocation. Proc. Natl. Acad. Sci. USA 2001, 98, 6969–6974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kagawa, T.; Sakai, T.; Suetsugu, N.; Oikawa, K.; Ishiguro, S.; Kato, T.; Tabata, S.; Okada, K.; Wada, M. Arabidopsis NPL1: A phototropin homolog controlling the chloroplast high-light avoidance response. Science 2001, 291, 2138–2141. [Google Scholar] [CrossRef]
- Jarillo, J.A.; Gabrys, H.; Capel, J.; Alonso, J.M.; Ecker, J.R.; Cashmore, A.R. Phototropin-related NPL1 controls chloroplast relocation induced by blue light. Nature 2001, 410, 952–954. [Google Scholar] [CrossRef]
- Quail, P.H. Photosensory perception and signalling in plant cells: New paradigms? Curr. Opin. Cell Biol. 2002, 14, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Suetsugu, N.; Wada, M. Chloroplast photorelocation movement mediated by phototropin family proteins in green plants. Biol. Chem. 2007, 388, 927–935. [Google Scholar] [CrossRef] [PubMed]
- Ishishita, K.; Higa, T.; Tanaka, H.; Inoue, S.; Chung, A.; Ushijima, T.; Matsushita, T.; Kinoshita, T.; Nakai, M.; Wada, M.; et al. Phototropin2 Contributes to the Chloroplast Avoidance Response at the Chloroplast-Plasma Membrane Interface(1)([CC-BY]). Plant Physiol. 2020, 183, 304–316. [Google Scholar] [CrossRef] [Green Version]
- Christie, J.M. Phototropin blue-light receptors. Annu. Rev. Plant Biol. 2007, 58, 21–45. [Google Scholar] [CrossRef] [Green Version]
- Kong, S.G.; Wada, M. Recent advances in understanding the molecular mechanism of chloroplast photorelocation movement. Biochim. Biophys. Acta 2014, 1837, 522–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schnabel, J.; Hombach, P.; Waksman, T.; Giuriani, G.; Petersen, J.; Christie, J.M. A chemical genetic approach to engineer phototropin kinases for substrate labeling. J. Biol. Chem. 2018, 293, 5613–5623. [Google Scholar] [CrossRef] [Green Version]
- Takemiya, A.; Shimazaki, K. Arabidopsis phot1 and phot2 phosphorylate BLUS1 kinase with different efficiencies in stomatal opening. J. Plant Res. 2016, 129, 167–174. [Google Scholar] [CrossRef]
- Kasahara, M.; Kagawa, T.; Sato, Y.; Kiyosue, T.; Wada, M. Phototropins mediate blue and red light-induced chloroplast movements in Physcomitrella patens. Plant Physiol. 2004, 135, 1388–1397. [Google Scholar] [CrossRef] [Green Version]
- Kagawa, T.; Kasahara, M.; Abe, T.; Yoshida, S.; Wada, M. Function analysis of phototropin2 using fern mutants deficient in blue light-induced chloroplast avoidance movement. Plant Cell Physiol. 2004, 45, 416–426. [Google Scholar] [CrossRef]
- Suetsugu, N.; Mittmann, F.; Wagner, G.; Hughes, J.; Wada, M. A chimeric photoreceptor gene, NEOCHROME, has arisen twice during plant evolution. Proc. Natl. Acad. Sci. USA 2005, 102, 13705–13709. [Google Scholar] [CrossRef] [Green Version]
- Suetsugu, N.; Wada, M. Phytochrome-dependent photomovement responses mediated by phototropin family proteins in cryptogam plants. Photochem. Photobiol. 2007, 83, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Suetsugu, N.; Wada, M. Photoreceptor gene families in lower plants. In Handbook of Photosensory Receptors; Wiley-VCH: Weinheim, Germany, 2005; pp. 349–369. [Google Scholar]
- Davis, P.A.; Hangarter, R.P. Chloroplast movement provides photoprotection to plants by redistributing PSII damage within leaves. Photosynth. Res. 2012, 112, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Trojan, A.; Gabrys, H. Chloroplast Distribution in Arabidopsis thaliana (L.) Depends on Light Conditions during Growth. Plant Physiol. 1996, 111, 419–425. [Google Scholar] [CrossRef] [Green Version]
- Nozue, K.; Kanegae, T.; Imaizumi, T.; Fukuda, S.; Okamoto, H.; Yeh, K.-C.; Lagarias, J.C.; Wada, M. A phytochrome from the fern Adiantum with features of the putative photoreceptor NPH1. Proc. Natl. Acad. Sci. USA 1998, 95, 15826–15830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuboi, H.; Wada, M. Chloroplasts continuously monitor photoreceptor signals during accumulation movement. J. Plant Res. 2013, 126, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Tsuboi, H.; Wada, M. Speed of signal transfer in the chloroplast accumulation response. J. Plant Res. 2010, 123, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Kagawa, T.; Wada, M. Blue light-induced chloroplast relocation in Arabidopsis thaliana as analyzed by microbeam irradiation. Plant Cell Physiol. 2000, 41, 84–93. [Google Scholar] [CrossRef] [Green Version]
- Kagawa, T.; Wada, M. Phytochrome- and blue-light-absorbing pigment-mediated directional movement of chloroplasts in dark-adapted prothallial cells of fern Adiantum as analyzed by microbeam irradiation. Planta 1996, 198, 488–493. [Google Scholar] [CrossRef]
- Oikawa, K.; Yamasato, A.; Kong, S.G.; Kasahara, M.; Nakai, M.; Takahashi, F.; Ogura, Y.; Kagawa, T.; Wada, M. Chloroplast outer envelope protein CHUP1 is essential for chloroplast anchorage to the plasma membrane and chloroplast movement. Plant Physiol. 2008, 148, 829–842. [Google Scholar] [CrossRef]
- Whippo, C.W.; Khurana, P.; Davis, P.A.; DeBlasio, S.L.; DeSloover, D.; Staiger, C.J.; Hangarter, R.P. THRUMIN1 Is a Light-Regulated Actin-Bundling Protein Involved in Chloroplast Motility. Curr. Biol. 2011, 21, 59–64. [Google Scholar] [CrossRef] [Green Version]
- Suetsugu, N.; Kagawa, T.; Wada, M. An auxilin-like J-domain protein, JAC1, regulates phototropin-mediated chloroplast movement in Arabidopsis. Plant Physiol. 2005, 139, 151–162. [Google Scholar] [CrossRef] [Green Version]
- DeBlasio, S.L.; Luesse, D.L.; Hangarter, R.P. A plant-specific protein essential for blue-light-induced chloroplast movements. Plant Physiol. 2005, 139, 101–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suetsugu, N.; Kagawa, T.; Wada, M. Analysis of all auxilin-like J-domain protein, JAC1, regulates phototropin-mediated chloroplast movement in Arabidopsis thaliana. Plant Cell Physiol. 2006, 47, S160. [Google Scholar]
- Luesse, D.R.; DeBlasio, S.L.; Hangarter, R.P. Plastid movement impaired 2, a new gene involved in normal blue-light-induced chloroplast movements in Arabidopsis. Plant Physiol. 2006, 141, 1328–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kodama, Y.; Suetsugu, N.; Kong, S.G.; Wada, M. Two interacting coiled-coil proteins, WEB1 and PMI2, maintain the chloroplast photorelocation movement velocity in Arabidopsis. Proc. Natl. Acad. Sci. USA 2010, 107, 19591–19596. [Google Scholar] [CrossRef] [Green Version]
- Majumdar, A.; Kar, R.K. Chloroplast avoidance movement: A novel paradigm of ROS signalling. Photosynth. Res. 2020, 144, 109–121. [Google Scholar] [CrossRef]
- Aggarwal, C.; Labuz, J.; Gabrys, H. Phosphoinositides Play Differential Roles in Regulating Phototropin1-and Phototropin2-Mediated Chloroplast Movements in Arabidopsis. PLoS ONE 2013, 8, e55393. [Google Scholar] [CrossRef] [Green Version]
- Oikawa, K.; Kasahara, M.; Kiyosue, T.; Kagawa, T.; Suetsugu, N.; Takahashi, F.; Kanegae, T.; Niwa, Y.; Kadota, A.; Wada, M. CHLOROPLAST UNUSUAL POSITIONING1 is essential for proper chloroplast positioning. Plant Cell 2003, 15, 2805–2815. [Google Scholar] [CrossRef] [Green Version]
- von Braun, S.S.; Schleiff, E. The chloroplast outer membrane protein CHUP1 interacts with actin and profilin. Planta 2008, 227, 1151–1159. [Google Scholar] [CrossRef]
- Usami, H.; Maeda, T.; Fujii, Y.; Oikawa, K.; Takahashi, F.; Kagawa, T.; Wada, M.; Kasahara, M. CHUP1 mediates actin-based light-induced chloroplast avoidance movement in the moss Physcomitrella patens. Planta 2012, 236, 1889–1897. [Google Scholar] [CrossRef]
- Kong, S.G.; Arai, Y.; Suetsugu, N.; Yanagida, T.; Wada, M. Rapid Severing and Motility of Chloroplast-Actin Filaments Are Required for the Chloroplast Avoidance Response in Arabidopsis. Plant Cell 2013, 25, 572–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suetsugu, N.; Yamada, N.; Kagawa, T.; Yonekura, H.; Uyeda, T.Q.P.; Kadota, A.; Wada, M. Two kinesin-like proteins mediate actin-based chloroplast movement in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2010, 107, 8860–8865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Y.H.; Ito, S.; Imaizumi, T. Flowering time regulation: Photoperiod-and temperature-sensing in leaves. Trends Plant Sci. 2013, 18, 575–583. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.H.; Smith, R.W.; To, B.J.; Millar, A.J.; Imaizumi, T. FKF1 conveys timing information for CONSTANS stabilization in photoperiodic flowering. Science 2012, 336, 1045–1049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, Y.; Weigel, D. Move on up, it’s time for change—Mobile signals controlling photoperiod-dependent flowering. Genes Dev. 2007, 21, 2371–2384. [Google Scholar] [CrossRef] [Green Version]
- de Montaigu, A.; Tóth, R.; Coupland, G. Plant development goes like clockwork. Trends Genet. 2010, 26, 296–306. [Google Scholar] [CrossRef]
- Amasino, R. Seasonal and developmental timing of flowering. Plant J. 2010, 61, 1001–1013. [Google Scholar] [CrossRef]
- Valverde, F.; Mouradov, A.; Soppe, W.; Ravenscroft, D.; Samach, A.; Coupland, G. Photoreceptor regulation of CONSTANS protein in photoperiodic flowering. Science 2004, 303, 1003–1006. [Google Scholar] [CrossRef] [Green Version]
- Imaizumi, T.; Tran, H.G.; Swartz, T.E.; Briggs, W.R.; Kay, S.A. FKF1 is essential for photoperiodic-specific light signalling in Arabidopsis. Nature 2003, 426, 302–306. [Google Scholar] [CrossRef]
- Zoltowski, B.D.; Imaizumi, T. Chapter Nine-Structure and Function of the ZTL/FKF1/LKP2 Group Proteins in Arabidopsis. In The Enzymes; Machida, Y., Lin, C., Tamanoi, F., Eds.; Academic Press: Cambridge, MA, USA, 2014; Volume 35, pp. 213–239. [Google Scholar]
- Demarsy, E.; Fankhauser, C. Higher plants use LOV to perceive blue light. Curr. Opin. Plant Biol. 2009, 12, 69–74. [Google Scholar] [CrossRef]
- Takahashi, N.; Kuroda, H.; Kuromori, T.; Hirayama, T.; Seki, M.; Shinozaki, K.; Shimada, H.; Matsui, M. Expression and Interaction Analysis of Arabidopsis Skp1-Related Genes. Plant Cell Physiol. 2004, 45, 83–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gehl, C.; Waadt, R.; Kudla, J.; Mendel, R.R.; Hansch, R. New GATEWAY vectors for High Throughput Analyses of Protein-Protein Interactions by Bimolecular Fluorescence Complementation. Mol. Plant 2009, 2, 1051–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, Y.-H.; Panzeri, D.; Kadota, Y.; Huang, Y.-C.; Huang, P.-Y.; Tao, C.-N.; Roux, M.; Chien, H.-C.; Chin, T.-C.; Chu, P.-W. The Arabidopsis malectin-like/LRR-RLK IOS1 is critical for BAK1-dependent and BAK1-independent pattern-triggered immunity. Plant Cell 2016, 28, 1701–1721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, N.; Yuan, S.; Li, Z.; Zhou, M.; Wu, P.; Hu, Q.; Wang, L.; Mendu, V.; Luo, H. STRESS INDUCED FACTOR 2, a leucine-rich repeat kinase regulates basal plant pathogen defense. Plant Physiol. 2018, 176, 01266–02017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso, J.M.; Stepanova, A.N.; Leisse, T.J.; Kim, C.J.; Chen, H.; Shinn, P.; Stevenson, D.K.; Zimmerman, J.; Barajas, P.; Cheuk, R.; et al. Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 2003, 301, 653–657. [Google Scholar] [CrossRef] [Green Version]
- Wada, M.; Kong, S.-G. Analysis of Chloroplast Movement and Relocation in Arabidopsis. In Chloroplast Research in Arabidopsis: Methods and Protocols, Volume I; Jarvis, R.P., Ed.; Humana Press: Totowa, NJ, USA, 2011; pp. 87–102. [Google Scholar]
- Ito, S.; Song, Y.H.; Imaizumi, T. LOV Domain-Containing F-Box Proteins: Light-Dependent Protein Degradation Modules in Arabidopsis. Mol. Plant 2012, 5, 573–582. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.D.; Kim, M.R.; Kang, M.Y.; Cha, J.Y.; Han, S.H.; Nawkar, G.M.; Sakuraba, Y.; Lee, S.Y.; Imaizumi, T.; McClung, C.R.; et al. The F-box protein FKF1 inhibits dimerization of COP1 in the control of photoperiodic flowering. Nat. Commun. 2017, 8, 2259. [Google Scholar] [CrossRef] [Green Version]
- Kubota, A.; Kita, S.; Ishizaki, K.; Nishihama, R.; Yamato, K.T.; Kohchi, T. Co-option of a photoperiodic growth-phase transition system during land plant evolution. Nat. Commun. 2014, 5, 3668. [Google Scholar] [CrossRef] [Green Version]
- Kagawa, T.; Wada, M. Velocity of chloroplast avoidance movement is fluence rate dependent. Photochem. Photobiol. Sci. 2004, 3, 592–595. [Google Scholar] [CrossRef]
- Tsuboi, H.; Yamashita, H.; Wada, M. Chloroplasts do not have a polarity for light-induced accumulation movement. J. Plant Res. 2009, 122, 131–140. [Google Scholar] [CrossRef]
- Sztatelman, O.; Łabuz, J.; Hermanowicz, P.; Banaś, A.K.; Zgłobicki, P.; Aggarwal, C.; Nadzieja, M.; Krzeszowiec, W.; Strzałka, W.; Gabryś, H. Fine tuning chloroplast movements through physical interactions between phototropins. J. Exp. Bot. 2016, 67, 4963–4978. [Google Scholar] [CrossRef] [PubMed]
- Hogewoning, S.W.; Trouwborst, G.; Maljaars, H.; Poorter, H.; van Ieperen, W.; Harbinson, J. Blue light dose-responses of leaf photosynthesis, morphology, and chemical composition of Cucumis sativus grown under different combinations of red and blue light. J. Exp. Bot. 2010, 61, 3107–3117. [Google Scholar] [CrossRef] [PubMed]
- Johkan, M.; Shoji, K.; Goto, F.; Hashida, S.-N.; Yoshihara, T. Blue light-emitting diode light irradiation of seedlings improves seedling quality and growth after transplanting in red leaf lettuce. HortScience 2010, 45, 1809–1814. [Google Scholar] [CrossRef] [Green Version]
- Terfa, M.T.; Solhaug, K.A.; Gislerød, H.R.; Olsen, J.E.; Torre, S. A high proportion of blue light increases the photosynthesis capacity and leaf formation rate of Rosa× hybrida but does not affect time to flower opening. Physiol. Plant. 2013, 148, 146–159. [Google Scholar] [CrossRef] [PubMed]
- Rao, X.; Huang, X.; Zhou, Z.; Lin, X. An improvement of the 2^(-delta delta CT) method for quantitative real-time polymerase chain reaction data analysis. Biostat. Bioinform. Biomath. 2013, 3, 71–85. [Google Scholar]
- Vojtek, A.B.; Hollenberg, S.M. Ras-Raf interaction: Two-hybrid analysis. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1995; Volume 255, pp. 331–342. [Google Scholar]
- Béranger, F.; Aresta, S.; de Gunzburg, J.; Camonis, J. Getting more from the two-hybrid system: N-terminal fusions to LexA are efficient and sensitive baits for two-hybrid studies. Nucleic Acids Res. 1997, 25, 2035–2036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartel, P.L.; Fields, S. Analyzing protein-protein interactions using two-hybrid system. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1995; Volume 254, pp. 241–263. [Google Scholar]
- Fromont-Racine, M.; Rain, J.-C.; Legrain, P. Toward a functional analysis of the yeast genome through exhaustive two-hybrid screens. Nat. Genet. 1997, 16, 277–282. [Google Scholar] [CrossRef]
- Matsushima, R.; Maekawa, M.; Kusano, M.; Kondo, H.; Fujita, N.; Kawagoe, Y.; Sakamoto, W. Amyloplast-Localized SUBSTANDARD STARCH GRAIN4 Protein Influences the Size of Starch Grains in Rice Endosperm. Plant Physiol. 2014, 164, 623–636. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, N.; Mendu, L.; Ghose, K.; Witte, C.S.; Frugoli, J.; Mendu, V. FKF1 Interacts with CHUP1 and Regulates Chloroplast Movement in Arabidopsis. Plants 2023, 12, 542. https://doi.org/10.3390/plants12030542
Yuan N, Mendu L, Ghose K, Witte CS, Frugoli J, Mendu V. FKF1 Interacts with CHUP1 and Regulates Chloroplast Movement in Arabidopsis. Plants. 2023; 12(3):542. https://doi.org/10.3390/plants12030542
Chicago/Turabian StyleYuan, Ning, Lavanya Mendu, Kaushik Ghose, Carlie Shea Witte, Julia Frugoli, and Venugopal Mendu. 2023. "FKF1 Interacts with CHUP1 and Regulates Chloroplast Movement in Arabidopsis" Plants 12, no. 3: 542. https://doi.org/10.3390/plants12030542
APA StyleYuan, N., Mendu, L., Ghose, K., Witte, C. S., Frugoli, J., & Mendu, V. (2023). FKF1 Interacts with CHUP1 and Regulates Chloroplast Movement in Arabidopsis. Plants, 12(3), 542. https://doi.org/10.3390/plants12030542