
Testing Explant Sources, Culture Media, and Light Conditions for the Improvement of Organogenesis in Pinus ponderosa (P. Lawson and C. Lawson)

 ,
,  , , , , ,
, , , , ,  ,
,  and
and
Abstract
:1. Introduction
2. Results
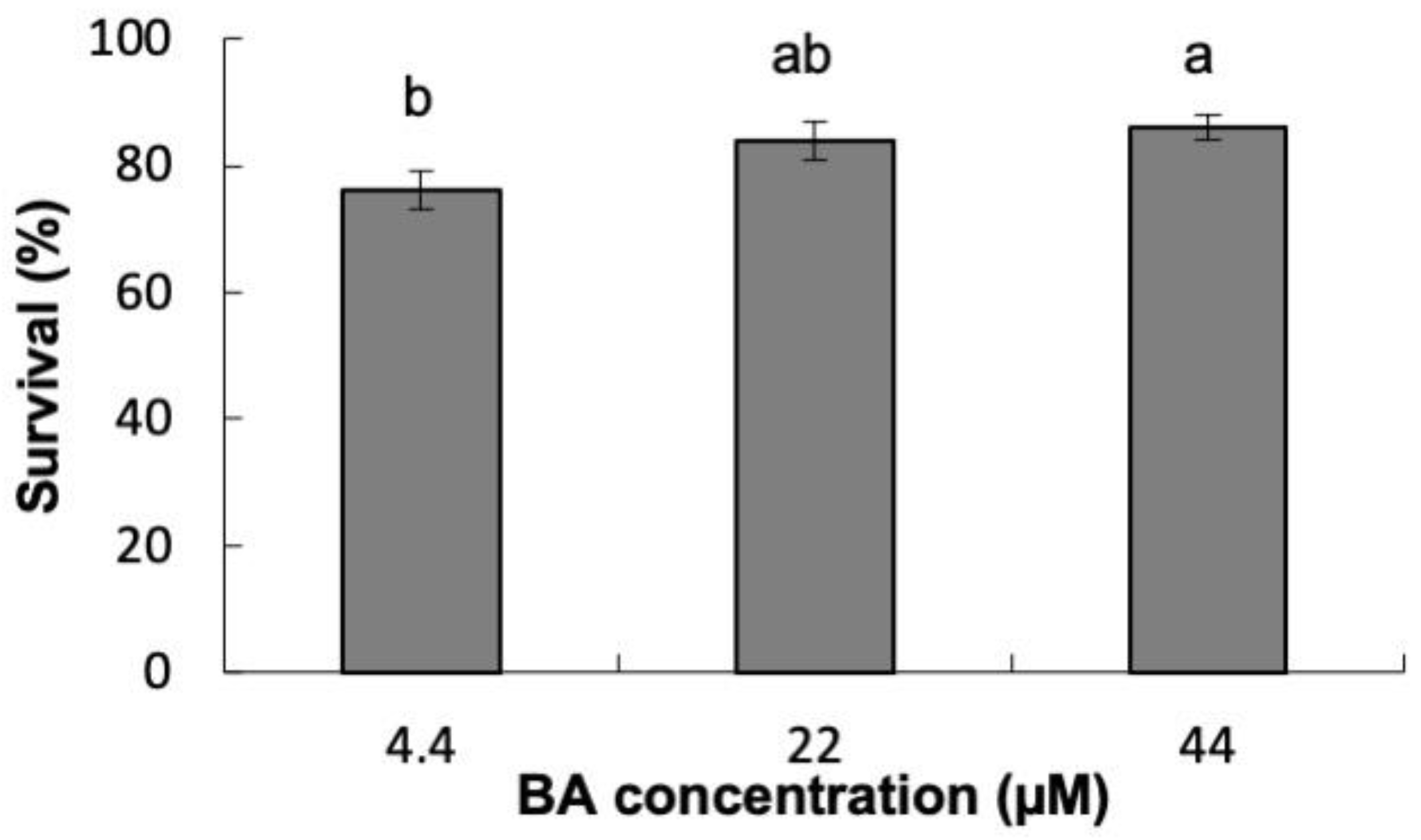
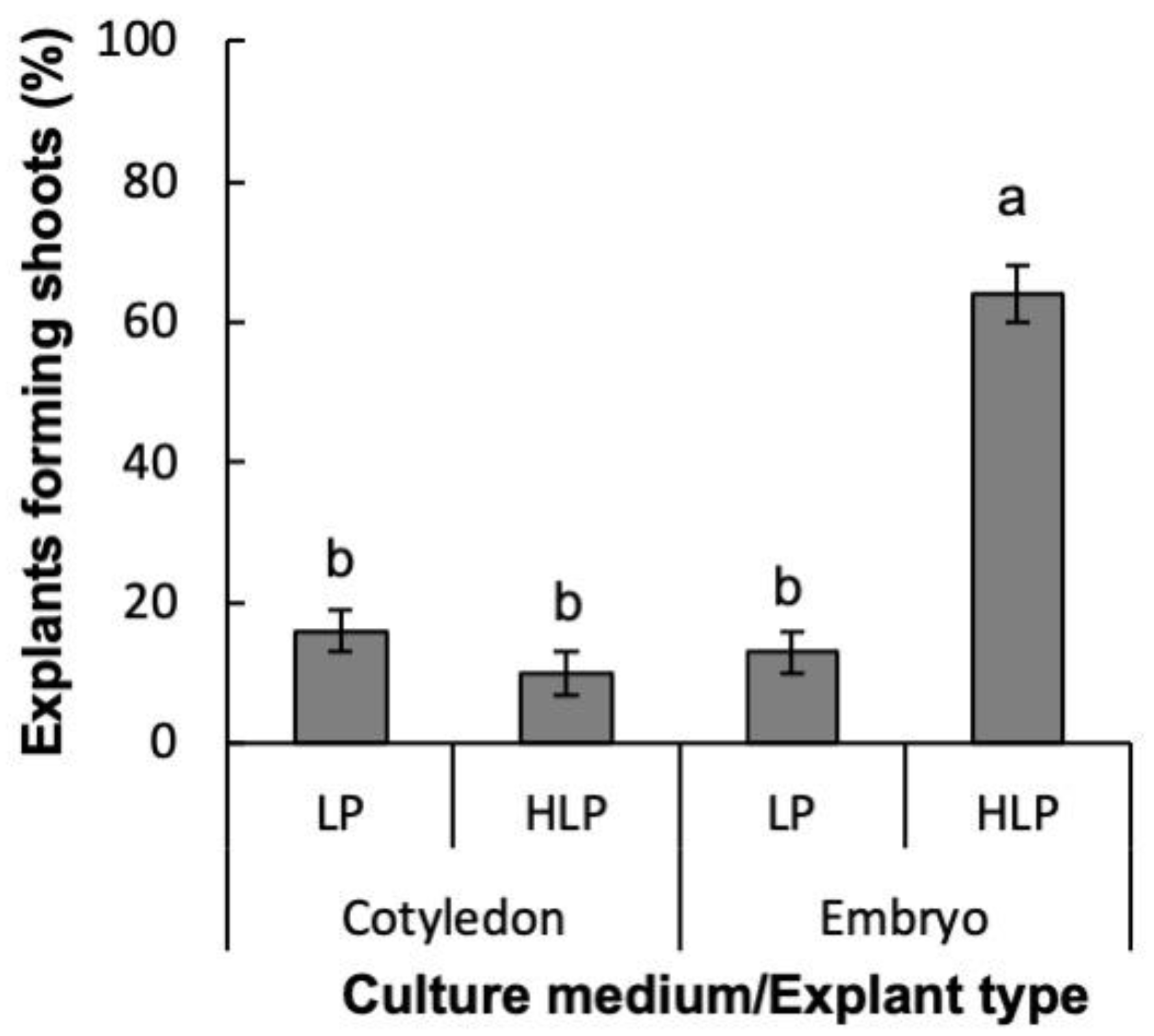
2.1. Induction of Organogenesis
2.1.1. Experiment 1
2.1.2. Experiment 2
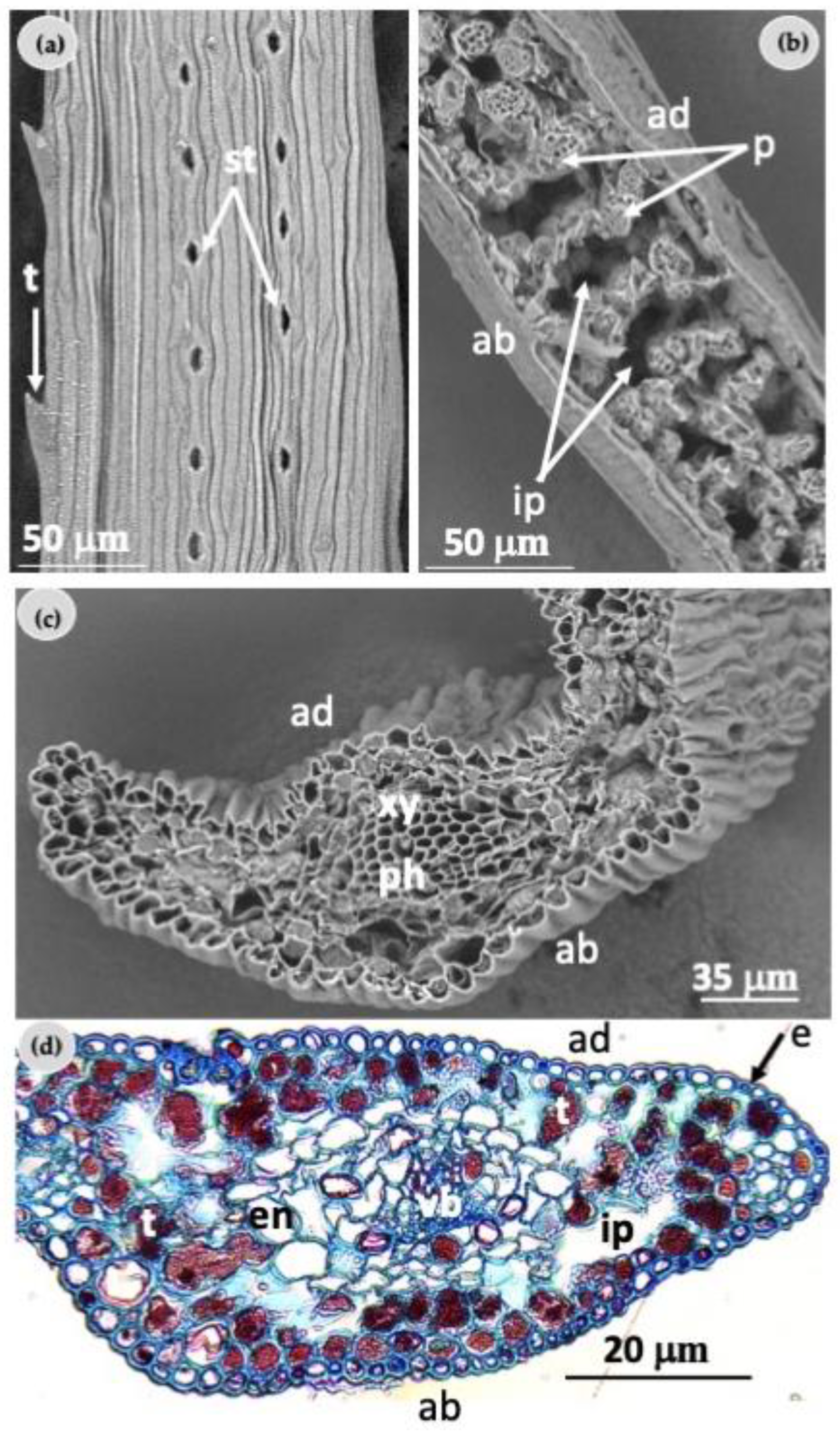
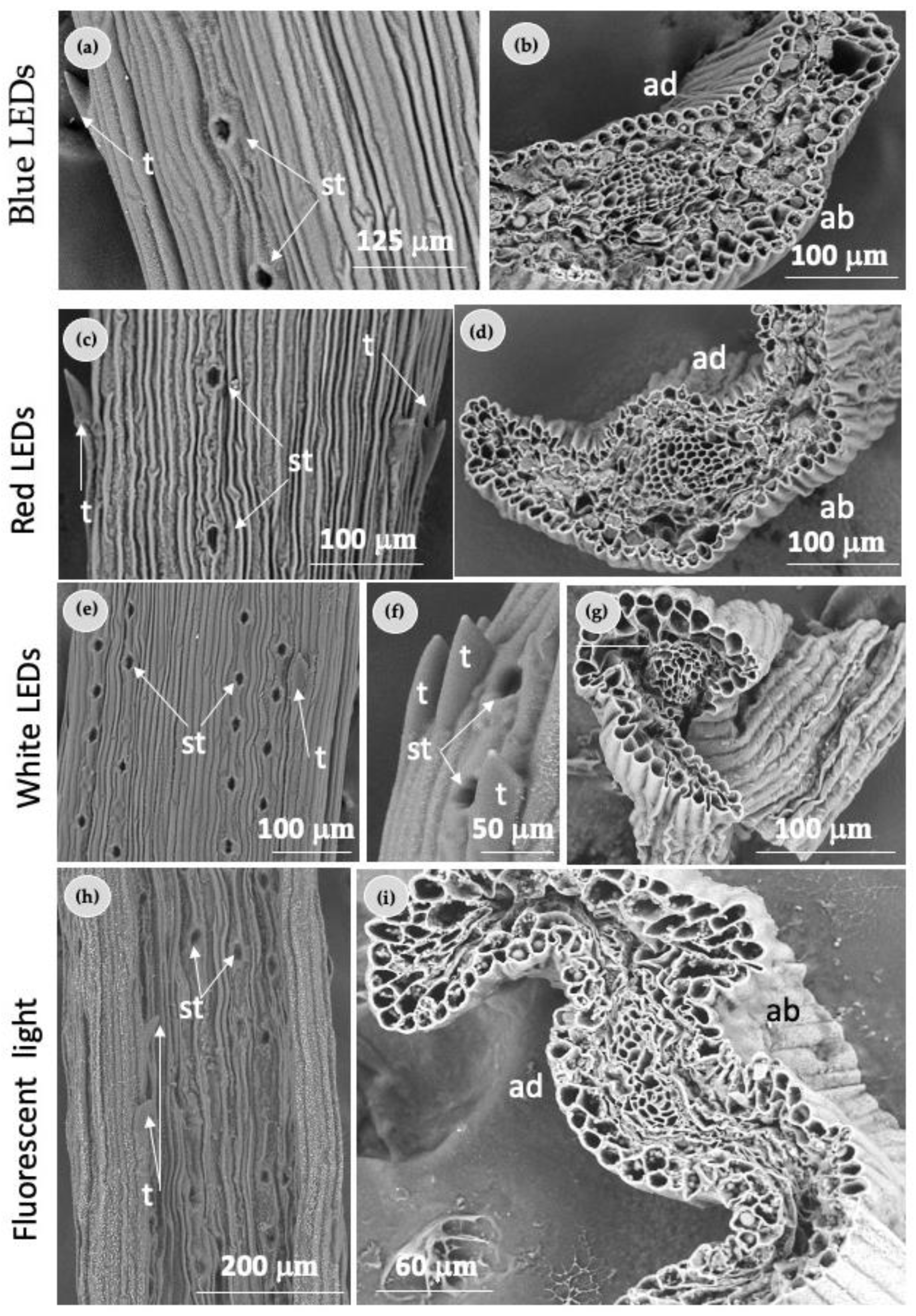
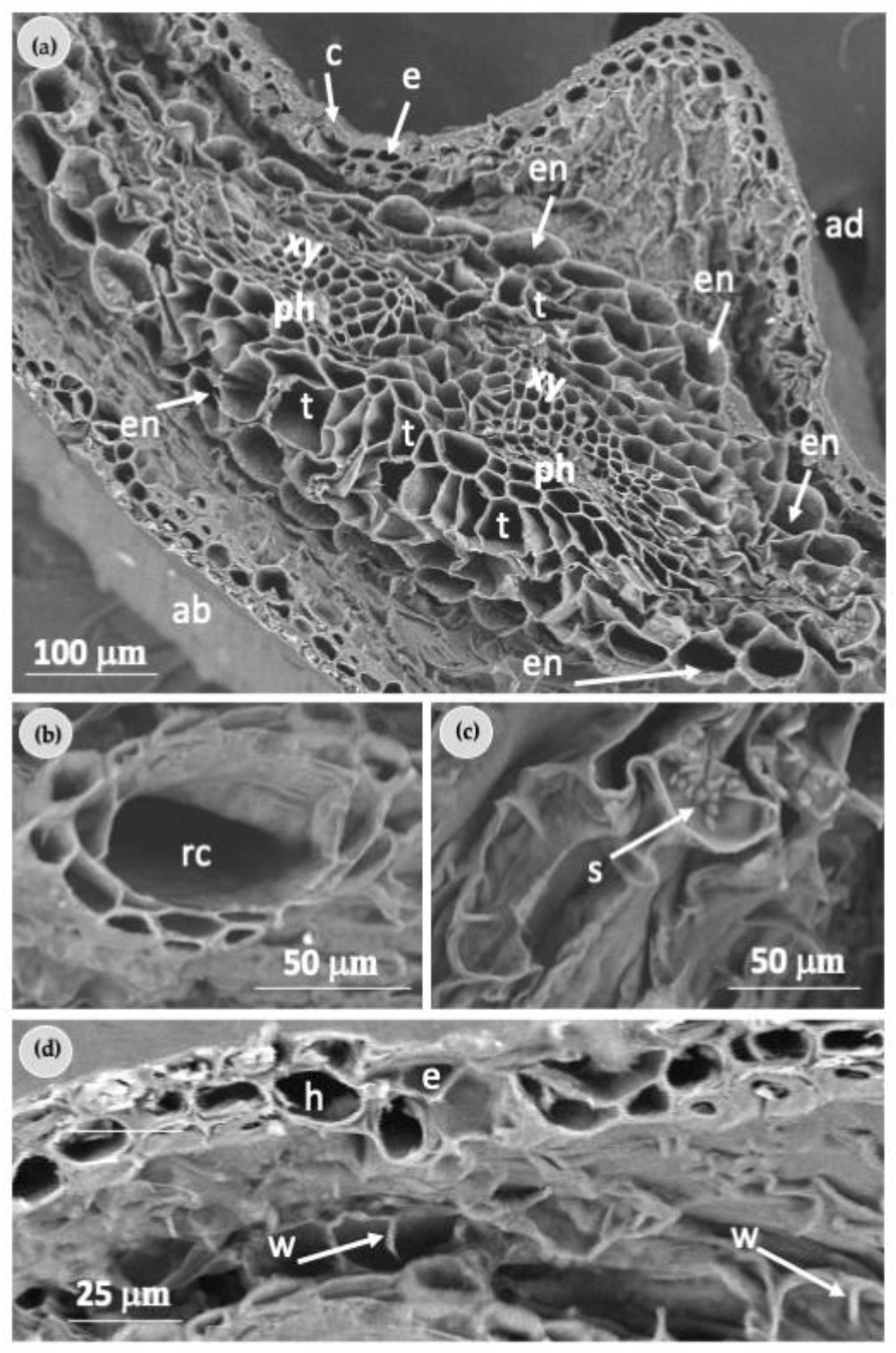
2.2. Morphological Characterization of Needles
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Sterilization
4.3. Organogenic Process
4.3.1. Experiment 1
4.3.2. Experiment 2
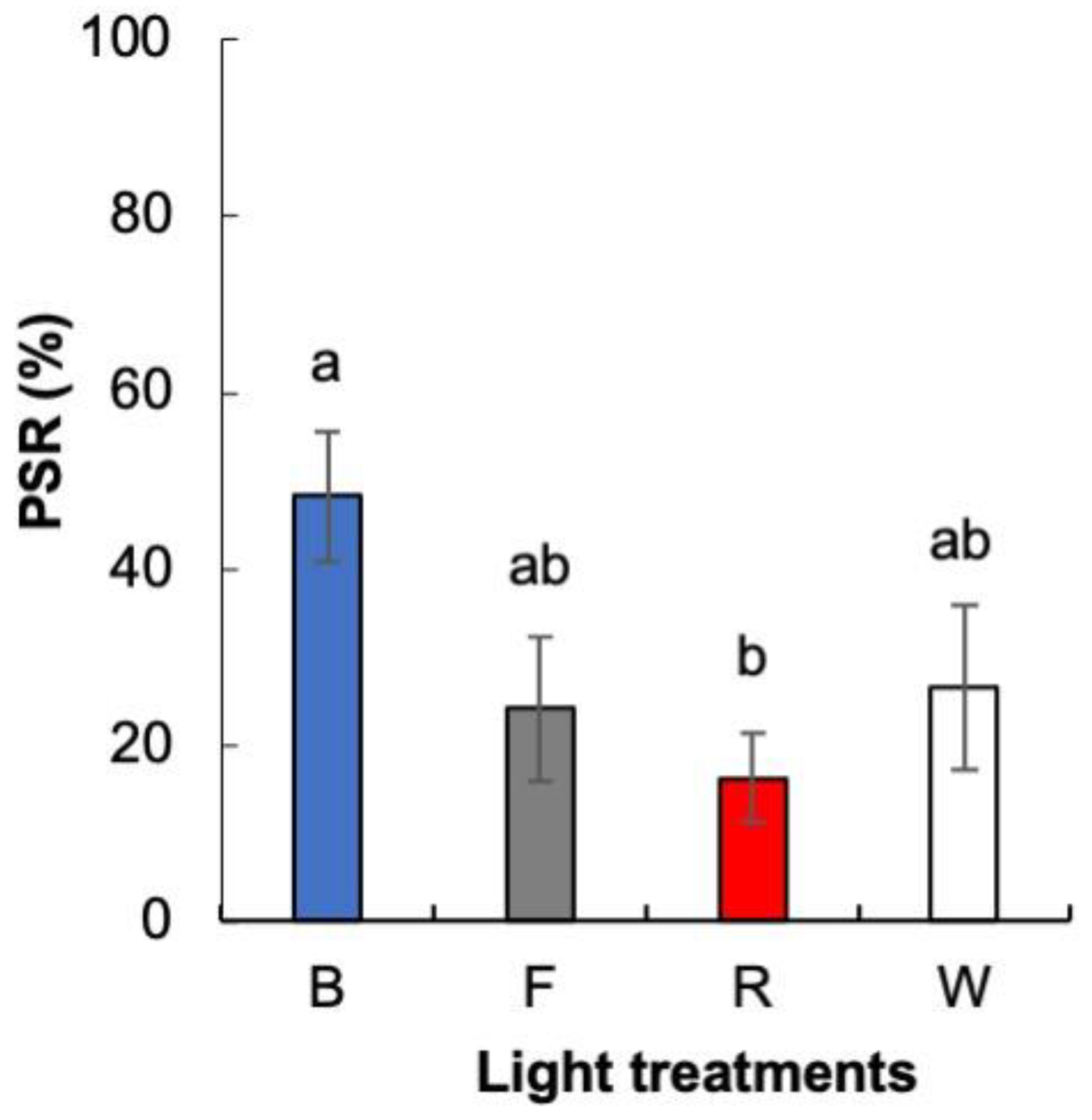
4.3.3. Rooting and Adaptation
4.4. Morphological Characterization of Needles
4.4.1. Light Microscopy Analysis
4.4.2. Scanning Electron Microscopy
4.5. Data Collection and Statistical Analysis
4.5.1. Experiment 1
4.5.2. Experiment 2
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Armenise, L.; Simeone, M.C.; Piredda, R.; Schirone, B. Validation of DNA barcoding as an efficient tool for taxon identification and detection of species diversity in Italian conifers. Eur. J. For. Res. 2012, 131, 1337–1353. [Google Scholar] [CrossRef]
- Farjon, A. The kew review conifers of the world. Kew Bull. 2018, 73, 8. [Google Scholar] [CrossRef]
- Ellis, D.; Bilderback, D.E. Ponderosa pine (Pinus ponderosa P. Lawson & C. Lawson). In Biotechnology in Agriculture and Forestry 16, Trees III; Bajaj, Y., Ed.; Springer: Berlin, Germany, 1991; pp. 339–357. [Google Scholar] [CrossRef]
- Raish, C.; Yong, W.; Marzluff, J. Contemporary human use of south western ponderosa pine forests. In Songbird Ecology in South Western Ponderosa Pine Forests: A Literature Review; Block, W.M., Finch, D.M., Eds.; Rocky Mountain Research Station, Forest Service, U.S. Department of Agriculture: Fort Collins, CO, USA, 1997; pp. 28–42. [Google Scholar] [CrossRef]
- Miller, M. Fire Autecology. In Wildland Fire in Ecosystems: Effects of Fire on Flora; Tech. Rep. RMRS-GTR-42; Brown, J., Smith, J., Eds.; Rocky Mountain Research Station, Forest Service, U.S. Department of Agriculture: Ogden, UT, USA, 2000; Volume 2, p. 257. [Google Scholar] [CrossRef]
- Hall, F.C. Fire history-blue mountains, Oregon. In Proceedings of the Fire History Workshop, Tucson, AZ, USA, 20–24 October 1980; Stokes, M.A., Dieterich, J.H., Eds.; Rocky Mountain Forest and Range Experiment Station, Forest Service, U.S. Department of Agriculture: Fort Collins, CO, USA, 1980; pp. 75–81. [Google Scholar] [CrossRef]
- Fitzgerald, S. Fire ecology of Ponderosa pine and the rebuilding of fire-resilient ponderosa pine ecosystems. In Proceedings of the Symposium on Ponderosa Pine: Issues, Trends, and Management, Klamath Falls, OR, USA, 18–21 October 2005; Pacific Southwest Research Station, Forest Service, U.S. Department of Agriculture: Albany, CA, USA, 2005; pp. 197–225. [Google Scholar] [CrossRef]
- Ellis, D.D.; Bilderback, D.E. Multiple bud formation by cultured embryos of Pinus ponderosa. J. Plant Physiol. 1984, 115, 201–204. [Google Scholar] [CrossRef]
- Phillips, G.C. In vitro morphogenesis in plants—Recent advances. Vitr. Cell. Dev. Biol. Plant. 2004, 40, 342–345. [Google Scholar] [CrossRef]
- Kulus, D.; Woźny, A. Influence of light conditions on the morphogenetic and biochemical response of selected ornamental plant species under in vitro conditions: A mini-review. Bio Technol. 2020, 101, 75–83. [Google Scholar] [CrossRef]
- Alallaq, S.; Ranjan, A.; Brunoni, F.; Novák, O.; Lakehal, A.; Bellini, C. Red light controls adventitious root regeneration by modulating hormone homeostasis in Picea abies seedlings. Front. Plant Sci. 2020, 11, 586140. [Google Scholar] [CrossRef]
- Larraburu, E.E.; Correa, G.S.; Llorente, B.E. In vitro development of yellow lapacho (Bignoniaceae) using high-power light emitting diode. Rev. Arvore. 2018, 42, e420508. [Google Scholar] [CrossRef]
- Fang, W.; Jao, R.C. A review on artificial lighting of tissue cultures and transplants. In Transplant Production in the 21st Century; Kubota, C., Chun, C., Eds.; Springer: Dordrecht, The Netherlands, 2000; pp. 108–113. [Google Scholar] [CrossRef]
- Ellis, D.D.; Judd, R.C. SDS-PAGE analysis of bud-forming cotyledons of Pinus ponderosa. Plant Cell Tissue Organ Cult. 1987, 11, 57–65. [Google Scholar] [CrossRef]
- Tuskan, G.A.; Sargent, W.A.; Rensema, T.; Walla, J.A. Influence of plant growth regulators, basal media and carbohydrate levels on the in vitro development of Pinus ponderosa (Dougl. Ex Law.) cotyledon explants. Plant Cell Tissue Organ Cult. 1990, 20, 47–52. [Google Scholar] [CrossRef]
- Lin, Y.; Wagner, M.R.; Heidmann, L.J. In vitro formation of axillary buds by immature shoots of Ponderosa Pine. Plant Cell Tissue Organ Cult. 1991, 26, 161–166. [Google Scholar] [CrossRef]
- Mazari, A.; Camm, E.L. Effect of cytokinins on plastid development and photosynthetic polypeptides during organogenesis of Pinus ponderosa Dougl. cotyledons cultured in vitro. Plant Cell Tissue Organ Cult. 1993, 33, 81–89. [Google Scholar] [CrossRef]
- Castander-Olarieta, A.; Moncaleán, P.; Montalbán, I.A. Somatic Embryogenesis in Pines. In Somatic Embryogenesis. Methods in Molecular Biology; Ramírez-Mosqueda, M.A., Ed.; Humana: New York, NY, USA, 2022; Volume 2527, pp. 41–46. [Google Scholar] [CrossRef]
- Moncaleán, P.; Alonso, P.; Centeno, M.L.; Cortizo, M.; Rodríguez, A.; Fernández, B.; Ordás, R.J. Organogenic responses of Pinus pinea cotyledons to hormonal treatments: BA metabolism and cytokinin content. Tree Physiol. 2005, 25, 1–9. [Google Scholar] [CrossRef]
- Montalbán, I.A.; De Diego, N.; Moncaleán, P. Testing novel cytokinins for improved in vitro adventitious shoots formation and subsequent ex vitro performance in Pinus radiata. Forestry 2011, 84, 363–373. [Google Scholar] [CrossRef] [Green Version]
- Do Nascimento, A.M.M.; Montalbán, I.A.; Llamazares De Miguel, D.; Goicoa, T.; Ugarte, M.D.; Moncaleán, P. High temperature and water deficit cause epigenetic changes in somatic plants of Pinus radiata D. Don. Plant Cell Tissue Organ Cult. 2022, 151, 107–121. [Google Scholar] [CrossRef]
- Castander-Olarieta, A.; Pereira, C.; Mendes, V.M.; Correia, S.; Manadas, B.; Canhoto, J.; Montalbán, I.A.; Moncaleán, P. Thermopriming-associated proteome and sugar content responses in Pinus radiata embryogenic tissue. Plant Sci. 2022, 321, 111327. [Google Scholar] [CrossRef]
- Pereira, C.; Castander-Olarieta, A.; Sales, E.; Montalbán, I.A.; Canhoto, J.; Moncaleán, P. Heat stress in Pinus halepensis somatic embryogenesis induction: Effect in DNA methylation and differential expression of stress-related genes. Plants 2021, 10, 2333. [Google Scholar] [CrossRef]
- Quoirin, M.; Lepoivre, P. Études des milieux adaptés aux cultures in vitro de Prunus. Acta Hortic. 1977, 78, 437–442. [Google Scholar] [CrossRef]
- Aitken-Christie, J.; Singh, A.P.; Davis, H. Multiplication of meristematic tissue: A new tissue culture system for radiata pine. In Genetic Manipulation of Woody Plants; Hanover, J.W., Keathley, D.E., Eds.; Plenum Publishing Corp: New York, NY, USA, 1988; pp. 413–432. [Google Scholar] [CrossRef]
- Payghamzadeh, K.; Kazemitabar, S.K. In vitro propagation of walnut—A review. Afr. J. Biotechnol. 2011, 10, 290–311. [Google Scholar] [CrossRef]
- Pereira, C.; Montalbán, I.A.; Pedrosa, A.; Tavares, J.; Pestryakov, A.; Bogdanchikova, N.; Canhoto, J.; Moncaleán, P. Regeneration of Pinus halepensis (Mill.) through organogenesis from apical shoot buds. Forests 2021, 12, 363. [Google Scholar] [CrossRef]
- Montalbán, I.A.; de Diego, N.; Moncaleán, P. Enhancing initiation and proliferation in Radiata Pine (Pinus radiata D. Don) somatic embryogenesis through seed family screening, zygotic embryo staging and media adjustments. Acta Physiol. Plant. 2012, 34, 451–460. [Google Scholar] [CrossRef]
- De Souza, R.A.; Padilha Dantas, P.V.; Cavalcante, P.D.F.; Tenório, R.R.; Houllou, L.M. Basic procedure for the in vitro propagation of brazilian trees for reforestation purposes. J. Environ. Anal. Prog. 2017, 2, 107–114. [Google Scholar] [CrossRef]
- Parzymies, M. Nano-silver particles reduce contaminations in tissue culture but decrease regeneration rate and slows down growth and development of Aldrovanda vesiculosa explants. Appl. Sci. 2021, 11, 3653. [Google Scholar] [CrossRef]
- Tung, H.T.; Thuong, T.T.; Cuong, D.M.; Luan, V.Q.; Hien, V.T.; Hieu, T.; Nam, N.B.; Phuong, H.T.N.; Van The Vinh, B.; Khai, H.D.; et al. Silver nanoparticles improved explant disinfection, in vitro growth, runner formation and limited ethylene accumulation during micropropagation of strawberry (Fragaria × Ananassa). Plant Cell Tissue Organ Cult. 2021, 145, 393–403. [Google Scholar] [CrossRef]
- Rojas-Vargas, A.; Castander-Olarieta, A.; Moncaleán, P.; Montalbán, I.A. Optimization of the micropropagation of elite adult trees of Sequoia sempervirens: Forest species of interest in the Basque Country, Spain. Rev. Bionatura. 2021, 6, 1511–1519. [Google Scholar] [CrossRef]
- Aitken, J.; Horgan, K.J.; Thorpe, T.A. Influence of explant selection on shoot forming capacity of juvenile tissue of Pinus radiata. Can. J. For. Res. 1981, 11, 112–117. [Google Scholar] [CrossRef]
- Saborio, F.; Dvorak, W.S.; Donahue, J.K.; Thorpe, T.A. In vitro regeneration of plantlets from mature embryos of Pinus ayacahuite. Tree Physiol. 1997, 17, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Lambardi, M.; Sharma, K.K.; Thorpe, T.A. Optimization of in vitro bud induction and plantlet formation from mature embryos of Aleppo Pine (Pinus halepensis Mill). Vitr. Cell. Dev. Biol. Plant. 1993, 29, 189–199. [Google Scholar] [CrossRef]
- De Diego, N.; Montalbán, I.A.; Paloma, M. Improved micropropagation protocol for maritime pine using zygotic embryos. Scand. J. For. Res. 2011, 26, 202–211. [Google Scholar] [CrossRef]
- Valverde-Cerdas, L.; Rojas-Vargas, A.; Hine-Gómez, A. In vitro propagation of Albizia guachapele, Cedrela odorata, Platymiscium pinnatum and Guaiacum sanctum. Plant Tissue Cult. Biotechnol. 2008, 18, 151–156. [Google Scholar] [CrossRef]
- Bairu, M.W.; Stirk, W.A.; Dolezal, K.; Van Staden, J. Optimizing the micropropagation protocol for the endangered Aloe polyphylla: Can meta-topolin and its derivatives serve as replacement for benzyladenine and zeatin? Plant Cell Tissue Organ Cult. 2007, 90, 15–23. [Google Scholar] [CrossRef]
- Sul, I.W.; Korban, S.S. Effects of salt formulations, carbon sources, cytokinins, and auxin on shoot organogenesis from cotyledons of Pinus pinea L. Plant Growth Regul. 2004, 43, 197–205. [Google Scholar] [CrossRef]
- Alonso, P.; Moncaleán, P.; Fernández, B.; Rodríguez, A.; Centeno, M.L.; Ordás, R.J. An Improved micropropagation protocol for Stone Pine (Pinus pinea L.). Ann. For. Sci. 2006, 63, 879–885. [Google Scholar] [CrossRef] [Green Version]
- Montalbán, I.A.; Novák, O.; Rolčik, J.; Strnad, M.; Moncaleán, P. Endogenous cytokinin and auxin profiles during in vitro organogenesis from vegetative buds of Pinus radiata adult trees. Physiol. Plant. 2013, 148, 214–231. [Google Scholar] [CrossRef]
- Rojas-Vargas, A.; Castander-Olarieta, A.; Montalbán, I.A.; Moncaleán, P. Influence of physico-chemical factors on the efficiency and metabolite profile of adult Pinus radiata D. Don bud organogenesis. Forests 2022, 13, 1455. [Google Scholar] [CrossRef]
- Chen, Y.C.; Chang, C.; Lin, H.L. Topolins and red light improve the micropropagation efficiency of passion fruit (Passiflora edulis Sims) “Tainung No. 1”. HortScience 2020, 55, 1337–1344. [Google Scholar] [CrossRef]
- Aremu, A.O.; Bairu, M.W.; Doležal, K.; Finnie, J.F.; Van Staden, J. Topolins: A panacea to plant tissue culture challenges? Plant Cell Tissue Organ Cult. 2012, 108, 1–16. [Google Scholar] [CrossRef]
- Werbrouck, S.P.O.; Strnad, M.; Van Onckelen, H.A.; Debergh, P.C. Meta-topolin, an alternative to benzyladenine in tissue culture? Physiol. Plant. 1996, 98, 291–297. [Google Scholar] [CrossRef]
- De Diego, N.; Montalbán, I.A.; Moncaleán, P. In vitro regeneration of adult Pinus sylvestris L. trees. S. Afr. J. Bot. 2010, 76, 158–162. [Google Scholar] [CrossRef]
- dos Santos Junior, C.F.; Rech, T.D.; Costa, M.D.; Navroski, M.C.; Boff, P.; Boff, M.I.C. Efeito da qualidade de luz na multiplicação e enraizamento in vitro de Cedrela odorata L. Ciênc. Florest. 2022, 32, 2419–2432. [Google Scholar] [CrossRef]
- Gupta, S.; Jatothu, B. Fundamentals and applications of light-emitting diodes (LEDs) in in vitro plant growth and morphogenesis. Plant Biotechnol. Rep. 2013, 7, 211–220. [Google Scholar] [CrossRef]
- Gupta, S.; Agarwal, A. Influence of LED lighting on in vitro plant regeneration and associated cellular redox balance. In Light Emitting Diodes for Agriculture: Smart Lighting; Gupta, S., Ed.; Springer: Singapore, 2017; pp. 273–298. [Google Scholar]
- Pawłowska, B.; Żupnik, M.; Szewczyk-Taranek, B.; Cioć, M. Impact of LED light sources on morphogenesis and levels of photosynthetic pigments in Gerbera jamesonii grown in vitro. Hortic. Environ. Biotechnol. 2018, 59, 115–123. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, K.; Gong, X.; Wang, H.; Gao, Y.; Wang, X.; Zeng, Z.; Hu, Y. Effects of different LEDs light spectrum on the growth, leaf anatomy, and chloroplast ultrastructure of potato plantlets in vitro and minituber production after transplanting in the greenhouse. J. Integr. Agric. 2020, 19, 108–119. [Google Scholar] [CrossRef]
- Gupta, S.; Sahoo, T.K. Light emitting diode (LED) induced alteration of oxidative events during in vitro shoot organogenesis of Curculigo orchioides Gaertn. Acta Physiol. Plant. 2015, 37, 233. [Google Scholar] [CrossRef]
- Gnasekaran, P.; Rahman, Z.A.; Chew, B.L.; Appalasamy, S.; Mariappan, V.; Subramaniam, S. Development of micropropagation system of Zingiber officinale var. rubrum Theilade using different spectrum light-emitting diode (LED) irradiation. Ind. Crops Prod. 2021, 170, 113748. [Google Scholar] [CrossRef]
- Chashmi, K.A.; Omran, V.O.G.; Ebrahimi, R.; Moradi, H.; Abdosi, V. Light quality affects protocorm-like body (PLB) formation, growth and development of in vitro plantlets of Phalaenopsis pulcherrima. Biol. Bull. Acad. Sci. 2022, 49, 476–484. [Google Scholar] [CrossRef]
- Guerra, F.; Badilla, L.; Cautin, R.; Castro, M. Micropropagation of Citronella mucronata D. Don, a vulnerable chilean endemic tree species. Plants 2022, 11, 2425. [Google Scholar] [CrossRef]
- Lai, C.S.; Kho, Y.H.; Chew, B.L.; Raja, P.B.; Subramaniam, S. Organogenesis of Cucumis metuliferus plantlets under the effects of LEDs and silver nanoparticles. S. Afr. J. Bot. 2022, 148, 78–87. [Google Scholar] [CrossRef]
- Poudel, P.R.; Kataoka, I.; Mochioka, R. Effect of red- and blue-light-emitting diodes on growth and morphogenesis of grapes. Plant Cell Tissue Organ Cult. 2008, 92, 147–153. [Google Scholar] [CrossRef]
- Boddi, S.; Bonzi, L.M.; Calamassi, R. Structure and ultrastructure of Pinus halepensis primary needles. Flora 2002, 197, 10–23. [Google Scholar] [CrossRef]
- Mitić, Z.S.; Zlatković, B.K.; Miljković, M.S.; Jovanović, S.Č.; Marin, P.D.; Stojanović, G.S. First insights into micromorphology of needle epicuticular waxes of south-eastern european Pinus nigra J. F. Arnold populations. Iheringia Sér. Bot. 2017, 72, 373–379. [Google Scholar] [CrossRef]
- Stabentheiner, E.; Pfeifhofer, H.W.; Juliane Peters, M.; Jiménez, S.; Morales, D.; Grill, D. Different surface characteristics of primary and secondary needles of Pinus canariensis. Flora 2004, 199, 90–99. [Google Scholar] [CrossRef]
- Vendrame, W.A.; Feuille, C.; Beleski, D.; Bueno, P.M.C. In vitro growth responses of ornamental Bananas (Musa sp.) as affected by light sources. Horticulturae 2022, 8, 92. [Google Scholar] [CrossRef]
- Zheng, L.; Van Labeke, M.C. Effects of different irradiation levels of light quality on Chrysanthemum. Sci. Hortic. 2018, 233, 124–131. [Google Scholar] [CrossRef]
- Silva, T.D.; Batista, D.S.; Fortini, E.A.; Castro, K.M.; Felipe, S.H.S.; Fernandes, A.M.; Sousa, R.M.J.; Chagas, K.; Silva, J.V.S.D.; Correia, L.N.F.; et al. Blue and red light affects morphogenesis and 20-hydroxyecdisone content of in vitro Pfaffia glomerata accessions. J. Photochem. Photobiol. B 2020, 203, 111761. [Google Scholar] [CrossRef]
- Fritsche, Y.; Pinheiro, M.V.M.; Guerra, M.P. Light quality and natural ventilation have different effects on protocorm development and plantlet growth stages of the in vitro propagation of Epidendrum fulgens (Orchidaceae). S. Afr. J. Bot. 2022, 146, 864–874. [Google Scholar] [CrossRef]
- Gnasekaran, P.; Rahman, Z.A.; Chew, B.L.; Uddain, J.; Mariappan, V.; Appalasamy, S.; Subramaniam, S. Regulation of chloroplast ultrastructure, adventitious bud anatomy and clonal fidelity of in vitro Zingiber officinale var. rubrum Theilade plantlets in response to LED spectral quality. Ind. Crops Prod. 2022, 181, 114825. [Google Scholar] [CrossRef]
- Castander-Olarieta, A.; Moncaleán, P.; Montalbán, I.A. Pinus canariensis plant regeneration through somatic embryogenesis. For. Syst. 2020, 29, 1–6. [Google Scholar] [CrossRef]
- Giacomolli Polesi, L.; Pacheco de Freitas Fraga, H.; Goeten, D.; Panato Back, F.; de Medeiros Oliveira, E.; Steiner, N.; Guerra, M.P. Morphohistological and biochemical features of the Guadua chacoensis (Bambusoideae; Poaceae) somatic embryogenesis. Plant Cell Tissue Organ Cult. 2022, 148, 479–499. [Google Scholar] [CrossRef]
- Rossi, S.; Anfodillo, T.; Menardi, R. Trephor: A new tool for sampling microcores from tree stems. IAWA J. 2006, 27, 89–97. [Google Scholar] [CrossRef]
- Marques, M.P.; Martins, J.; de Carvalho, L.A.E.B.; Zuzarte, M.R.; Costa, R.D.; Canhoto, C. Study of physiological and biochemical events leading to vitrification of Arbutus unedo L. cultured in vitro. Trees 2021, 35, 241–253. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Light Treatment | Root Induction (%) | Number of Roots per Explant | Length of the Longest Root (cm) |
|---|---|---|---|
| Blue LEDs | 7 ± 30 | 1.71 ± 0.29 | 1.60 ± 0.77 |
| Red LEDs | 8 ± 40 | 1.25 ± 0.25 | 0.98 ± 0.34 |
| White LEDs | 19 ± 80 | 2.40 ± 0.87 | 2.78 ± 1.64 |
| Fluorescent | 8 ± 40 | 2.00 ± 0.71 | 2.15 ± 1.35 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rojas-Vargas, A.; Castander-Olarieta, A.; do Nascimento, A.M.M.; Vélez, M.L.; Pereira, C.; Martins, J.; Zuzarte, M.; Canhoto, J.; Montalbán, I.A.; Moncaleán, P. Testing Explant Sources, Culture Media, and Light Conditions for the Improvement of Organogenesis in Pinus ponderosa (P. Lawson and C. Lawson). Plants 2023, 12, 850. https://doi.org/10.3390/plants12040850
Rojas-Vargas A, Castander-Olarieta A, do Nascimento AMM, Vélez ML, Pereira C, Martins J, Zuzarte M, Canhoto J, Montalbán IA, Moncaleán P. Testing Explant Sources, Culture Media, and Light Conditions for the Improvement of Organogenesis in Pinus ponderosa (P. Lawson and C. Lawson). Plants. 2023; 12(4):850. https://doi.org/10.3390/plants12040850
Chicago/Turabian StyleRojas-Vargas, Alejandra, Ander Castander-Olarieta, Antonia Maiara Marques do Nascimento, María Laura Vélez, Cátia Pereira, João Martins, Mónica Zuzarte, Jorge Canhoto, Itziar A. Montalbán, and Paloma Moncaleán. 2023. "Testing Explant Sources, Culture Media, and Light Conditions for the Improvement of Organogenesis in Pinus ponderosa (P. Lawson and C. Lawson)" Plants 12, no. 4: 850. https://doi.org/10.3390/plants12040850
APA StyleRojas-Vargas, A., Castander-Olarieta, A., do Nascimento, A. M. M., Vélez, M. L., Pereira, C., Martins, J., Zuzarte, M., Canhoto, J., Montalbán, I. A., & Moncaleán, P. (2023). Testing Explant Sources, Culture Media, and Light Conditions for the Improvement of Organogenesis in Pinus ponderosa (P. Lawson and C. Lawson). Plants, 12(4), 850. https://doi.org/10.3390/plants12040850